不同生境对苦竹鞭根形态结构及其异速生长的影响
2022-11-09应益山杨丽婷程建新兰春宝陈双林郭子武
应益山,杨丽婷,程建新,兰春宝,陈双林,郭子武*
(1 浙江省龙游县林业技术推广站,浙江龙游 324400;2 中国林业科学研究院亚热带林业研究所,杭州 311400)
鞭根系统是竹林生长发育的基础,对竹林生态系统功能良好发挥及生产力长期维持尤为重要[1]。竹子是典型的克隆植物,其鞭根系统具有较强的可塑性。当生境发生变化时,竹子通过调节鞭根系统的分布格局、形态塑性和结构,改变鞭根系统对养分斑块的敏感性以及养分获取能力,从而影响竹子生长发育、种间竞争关系等过程[2-3]。研究表明,随着郁闭度的增加,华西箭竹(Fargesianitida)通过增大分枝角度、叶生物量分配比例、比叶面积和叶面积率以提高光能利用效率,有效适应弱光环境;且林缘和林窗中华西箭竹间隔子(竹鞭)长度、间隔子直径和分枝强度均显著高于林内[4]。在异质光照生境下,处于光照较好环境下的紫耳箭竹(Fargesiadecurvata)会增加对地下茎(竹鞭)的投资,而处于光照较差环境下的紫耳箭竹则将较多的生物量投资于秆和叶片的生长[5],以促进高生长和叶片生物量积累,从而获得更多的光资源。在水分有效性较低的生境中,斑苦竹(Pleioblastusmaculata)会将更多的生物量分配到地下鞭根系统,从而促进竹鞭的伸长和生物量积累,这在一定程度上增加了斑苦竹对限制性资源水分的吸收[6]。可见,鞭根形态塑性反映了竹子对差异性生境的生态适应策略,因而,研究不同生境对竹子鞭根系统形态塑性及其异速生长的影响具有重要科学意义。
苦竹(Pleioblastusamarus)隶属禾本科竹亚科大明竹属,为地下茎复轴混生型竹种,生态适应性强,分布广,是优良笋材兼用竹种,具有较好的经济、生态和社会效益[7]。目前苦竹的研究多集中于生长发育规律[8]、叶片化学计量特征[9]、元素分布规律[7]、竹笋品质及苦竹林丰产培育技术[10-11]等方面。苦竹鞭根系统的研究仅限于竹鞭生长规律[12]、鞭根分布特征[13-14]等,而苦竹鞭根形态塑性及其相对增长关系对不同生境的响应机制尚不清楚,这极大地限制了对不同生境下竹子养分利用、觅养机制和生态适应策略的阐释。为此,本研究以苦竹鞭根系统为研究对象,选取相邻连续分布的苦竹纯林和苦竹-杉木(Cunninghamialanceolata)混交林两种林分类型,测定了苦竹纯林和混交林中心区及界面区的鞭根形态结构性状指标,并分析了其异速增长关系,试图探讨生境差异是否会对苦竹鞭根主要功能性状及生物量产生明显影响,进而改变其相对生长关系,从而产生形态塑性及其异速生长的适应性调节,最终初步阐明苦竹资源利用特征及权衡策略对不同生境的响应规律,为苦竹林可持续经营提供理论参考。
1 材料和方法
1.1 试验地概况
试验地位于浙江省龙游县溪口林场(119°11′38″E、28°51′45″N)。该地区属中亚热带季风气候,四季分明,年平均降水量1 621 mm,年平均气温17.3 ℃,极端高温和极端低温分别为41 ℃和-11.4 ℃,年平均无霜期261.5 d,年平均相对湿度80%,年平均日照时数1 769 h。土壤为山地黄壤,土层厚度100 cm以上,pH值为6.5。
1.2 试验方法
2021年10月依据试验区苦竹分布状况,选取海拔、坡度、坡向、立地条件基本一致的连续分布的苦竹纯林和苦竹-杉木混交林2种林分类型的试验林各3个。在各试验林中,以苦竹纯林和苦竹-杉木混交林交界区为界面区,分别在界面区苦竹林方向和苦竹-杉木混交林方向10 m以上和3 m处设置试验样地,即苦竹林中心区(CA)、苦竹林界面区(BA)、混交林界面区(BM)和混交林中心区(CM)4种条带长方形试验样地,样地大小长20 m、宽60 cm(图1)。调查各样地基本林分结构和立地状况(表1),分别在BA、BM 2个样地中心向两边设置2 m×2 m的样方,在CA、CM 2个样地中心向两边设置5 m×5 m的样方,对样方内的苦竹或杉木进行每木检尺,记录杉木与苦竹的数量、胸径、树高等,然后计算树种组成及密度。依据已有试验苦竹林地下鞭根系统主要分布于0~20 cm土层,分别在试验林的4个样地中随机取样,挖取6个10 cm ×10 cm ×20 cm(长×宽×深)的原状土块,取样时注意避开竹鞭,将取好的土块做好标记分别装入密封袋中,置于冰盒带回实验室。

表1 试验林样地基本情况
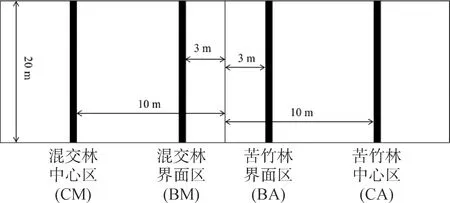
CM.混交林中心区;BM.混交林界面区;BA.苦竹林界面区;CA.苦竹林中心区;下同
1.3 指标测定
将各处理土样过筛冲洗,重复多次后除去草根及杉木等根系和其他杂物,用吸水纸吸干鞭根表面水分,挑选相对完整的鞭根进行形态结构性状的测定。将鞭根放置于万深LA-S系列植物图像扫描仪上,对鞭根进行扫描,扫描量不少于总量三分之一,运用万深LA-S系列植物图像分析系统测定苦竹鞭根根长、根直径(AvDiam)、表面积、节点数、根尖数和分形维数。然后将各处理扫描后的鞭根和未扫描的鞭根分别放入烘箱内,65 ℃烘干至恒重,使用电子天平(0.01 g)称重(g),计算苦竹鞭根比根长、比根面积以及根组织密度。将扫描后鞭根生物量和未扫描鞭根生物量相加得到各处理鞭根总生物量。
比根长(SRL, cm/g)=鞭根根长/干重
比根面积(SRA, cm2/g)=表面积/干重
鞭根根组织密度(RTD, g/cm3)=干重/体积
1.4 数据分析
试验数据在Excel 2010中进行数据处理和图表制作,采用SPSS 23.0的K-S检验进行数据的正态分布检验,采用SPSS 23.0进行单因素(One-way ANOVA)方差分析和邓肯(Duncan)多重比较(α=0.05),分析各处理苦竹鞭根比根长、比根面积、根组织密度、根直径、节点数、根尖数、分形维数和生物量的差异显著性,试验数据均表示为平均值±标准差(mean ± SD)。
同时,采用R4.0.3统计软件的Smart软件包对不同处理苦竹鞭根比根长、比根面积、根组织密度、根直径进行标准化主轴回归分析(Standardized Major Axis,SMA)。异速增长方程为Y=a×xb,经对数转化后为lgY=lg a + b lgx,其中方程斜率b为异速生长指数,x分别为比根面积、根组织密度、平均根直径,Y分别为比根长、比根面积、根组织密度;若|b|与1.0差异显著,表明x和Y间呈异速生长关系,同时比较不同处理斜率之间的差异性,若其无显著差异则计算共同斜率,并采用Wald 检验不同处理苦竹沿共同主轴位移差异的显著性[15-16]。
2 结果与分析
2.1 不同生境苦竹鞭根形态结构性状
由表2可知,苦竹林中心区(CA)至混交林中心区(CM)方向,苦竹鞭根比根长、比根面积呈升高趋势,根组织密度呈“V”型变化趋势,根尖数、节点数则呈“Λ”型变化趋势,根直径和分形维数呈降低趋势,且鞭根分形维数各处理间差异不显著(P>0.05)。与CA区相比,苦竹林界面区(BA)苦竹比根长、比根面积、根尖数、节点数显著增加,根直径显著降低,而根组织密度则无明显变化。与CM区相比,混交林界面区(BM)苦竹比根长显著降低,根直径显著增加,而比根面积、根组织密度、根尖数、节点数则无明显变化。BA区与BM区相比,二者间苦竹鞭根比根长、比根面积、根组织密度、根直径均无显著差异,但二者苦竹的比根长均显著高于CA区,却显著低于CM区,其鞭根直径表现则与之相反。可见,不同生境下苦竹鞭根性状存在明显差异,界面区苦竹鞭根形态塑性较强,拥有更高的鞭根活性以及更活跃的生理功能。

表2 试验林苦竹鞭根形态结构性状
2.2 不同生境苦竹鞭根生物量积累特征
从苦竹纯林中心区至混交林中心区方向,苦竹鞭根生物量呈逐渐降低趋势(图 2)。与CA 区相比,BA区苦竹鞭根生物量显著下降;与CM区相比,BM区苦竹鞭根生物量显著升高;但BA区和BM区苦竹鞭根生物量之间并无显著差异。由此可知,不同生境下苦竹鞭根生物量存在明显差异,界面区苦竹鞭根较混交林中心区有更高的生物量积累;界面区苦竹通过提高根系的水分、养分吸收能力来增强对空间和资源的竞争。

图2 试验林不同生境苦竹鞭根生物量
2.3 不同生境苦竹主要鞭根性状间异速生长关系
由图3分析可知,苦竹鞭根比根长与比根面积间存在共同斜率,且与1.00无显著差异,二者呈等速生长模式;苦竹鞭根比根长与根组织密度间、比根面积与根组织密度间均存在共同斜率,且均与-1.00无显著差异,两组性状间均呈负等速生长模式;苦竹鞭根比根长与根直径间、根组织密度与根直径间、比根面积与根直径间均存在共同斜率,且均与-1.00差异显著(P<0.001),三组性状间均呈负异速生长模式。经Wald检验,BA区与BM区各主要鞭根形态结构性状间的截距无明显差异,均显著高于CA区,且均显著低于CM区,说明各生境区苦竹沿共同主轴发生明显位移,且以BA差异性位移量最高。由此可知,生境对苦竹主要鞭根形态结构性状增长速率无显著影响,但却显著提高了苦竹林界面区主要鞭根形态结构性状的差异性位移量。

图3 试验林苦竹鞭根主要形态结构性状异速生长关系
3 讨 论
形态可塑性是植物适应环境的一种基本形式,当外界环境发生改变时,植物根系通过生物量积累和形态方面的变化来适应环境变化[17]。竹类植物因其克隆生长特性而具有更高的形态可塑性,可以最大程度地获取资源,并进行资源的再分配[18-19],从而提高其对不同生境的适应能力。本研究中,从苦竹林中心区至混交林中心区方向,苦竹鞭根生物量呈逐渐降低趋势,纯林界面区与混交林界面区苦竹鞭根生物量间差异不显著,但两者均显著低于纯林中心区,却显著高于混交林中心区。这说明不同生境下苦竹鞭根生物量存在明显差异,而界面区苦竹可通过克隆整合作用提高根系水分、养分吸收能力,增强对空间和资源的竞争,从而降低了苦竹林界面区与混交林界面区间苦竹鞭根生物量的差异。
比根长是根系形态结构与生理功能的一个重要指标,其高低与土壤资源有效性密切相关[20],可综合反映植物根系吸收资源的能力与生态适应性[21];比根面积的大小表示与土壤接触机会高低,可反映植物根系就地利用资源的能力;根组织密度与抗逆相关,反映了植物物质贮存和环境的耐受能力,是根系生长和吸收能力的重要指标[22]。本研究中,苦竹纯林中心区至混交林中心区方向,苦竹鞭根比根长、比根面积呈升高趋势。这可能是因为苦竹林中心区郁闭度低,强光会加快林地土壤水分散失,造成水资源相对匮乏[5],导致苦竹纯林鞭根比根长、比根面积显著低于其他处理。而竹-杉混交林经营一方面由于其林地郁闭度提高,土壤水分散失较慢,土壤孔隙度增加[23],处于竹-杉混交林中的苦竹可通过增加鞭根比根长、比根面积来增强对水分和养分的吸收。一般而言,同一物种的根直径越细,其生理功能越活跃[24-25]。本研究中,苦竹纯林界面区与混交林界面区间苦竹鞭根直径无显著差异,但均显著低于纯林中心区,却显著高于混交林中心区。这可能是因为界面区资源竞争较大,苦竹分株间通过克隆整合作用提高鞭根对水分、养分斑块的敏感性,导致界面区苦竹鞭根生理功能活跃。本研究发现,与纯林中心区相比,纯林界面区苦竹比根长、比根面积、根尖数、节点数显著增加,根直径显著减小。说明生境对苦竹林界面区苦竹鞭根性状产生显著影响,苦竹纯林界面区因郁闭度较小,没有杉木与之竞争,加之苦竹纯林界面区的立竹可以通过克隆整合作用得到混交林界面区立竹水分和养分资源的支持,从而显著促进了苦竹林界面区苦竹鞭根的生长。

表3 试验林苦竹鞭根主要形态结构性状间的相关生长指数及等速生长检验
异速增长采用幂函数方程来描述植物构件特征间的相对变化关系,是目前植物表型可塑性研究较为理想的方法[26]。植物性状间的异速关系是生态系统和植物进化关系共同影响的结果,在不同生态系统和植物群落间具有显著差异,说明植物在不同环境中的生长策略不同[27-28]。有研究指出较薄较细的根具有高的比根长,往往拥有更好的开发能力和更高的生长速度和养分吸收效率[29];相反,更厚更粗的根拥有更厚的皮层和皮质细胞层,因此需要在细胞壁中投入更多的物质和能量,具有更高的组织密度[30]。本研究中,苦竹鞭根比根长、比根面积、根组织密度与鞭根直径均呈异速增长关系,且存在共同斜率,表明较粗的鞭根拥有较低的比根长、根组织密度以及较高的比根面积,这说明在异质性环境下苦竹获取资源时更加趋向于采取增加面积和降低根直径的保守策略,同时也说明苦竹鞭根直径是影响苦竹获取异质资源的重要影响因素。本研究发现,生境对苦竹主要鞭根形态结构性状异速增长速率无明显影响,但显著提高了苦竹林界面区主要鞭根形态结构性状的差异性位移量,这说明界面区立竹形态可塑性更强,可通过克隆整合作用获取更多可利用资源用于生长发育,使得苦竹林界面区以及混交林界面区苦竹表现出更好的鞭根性状,确保竹林向有利于种群繁衍的方向进行。
综上所述,不同生境下苦竹林鞭根形态结构存在明显差异,界面区苦竹林鞭根形态结构可塑性较高,拥有更高的鞭根生长活性以及更活跃的生理功能。苦竹林界面区和混交林界面区苦竹较混交林中心区苦竹拥有更高的鞭根生物量,苦竹分株间的克隆整合增强了界面区苦竹鞭根的水分、养分获取能力及对空间和资源的竞争能力,从而显著提高了鞭根生物量和生长活性。不同生境下苦竹鞭根主要形态结构指标相对增长速率并无明显差异,苦竹为了最大化获取资源,趋向于采取增加比根表面积和降低鞭根直径的生态策略,其中根直径是苦竹获取异质资源的重要影响因素。
