基于SSR分子标记的中国黄连木遗传多样性分析
2022-11-09苏淑钗
任 重,白 倩,苏淑钗*
(1 北京林业大学 林学院,省部共建森林培育与保护教育部重点实验室,北京100083;2 北京林业大学,国家能源非粮生物质原料研发中心,北京 100083)
中国黄连木(PistaciachinensisBunge)是漆树科黄连木属落叶乔木,又名楷木、孔木、药树、茶树、黄连茶、木黄连、鸡冠木等。黄连木属植物有阿月浑子(P.veraL.)、中国黄连木(P.chinensisBunge)和大西洋黄连木(P.atlanticaDesf)等20余种或变种[1],中国有阿月浑子、中国黄连木、清香木3种[2]。
黄连木原产中国,分布很广,在中国华北、华中、华南、西南、西北、中南地区27个省、市、自治区都有其资源分布。由于其适应性强、萌芽力好、耐干旱耐瘠薄等特点,在荒山绿化中被广泛用作先锋树种[3]。21世纪以来,随着生物质能源在全球内的迅速兴起和发展,黄连木作为中国乡土木本油料树种,由于其可以全果榨油且果实含油率高、油脂品质好等独特的优势而被人们广泛的重视和研究,成为中国首批确定重点发展的生物质能源树种之一[4-5]。近年来,随着各地区黄连木雌雄同株特异资源的相继发现[6-7],关于黄连木性别分化决定机制的相关研究也日渐深入[8-13]。此外,黄连木还在用材、药用、观赏和绿化等方面具有广泛的利用价值[14]。多功能、多用途的利用价值,使得黄连木拥有着极为广阔的发展空间。
分子标记技术是研究植物遗传多样性的重要手段,能够有效揭示种内和种间的遗传变异。SSR又被称为微卫星标记,作为一种重复性好、多态性高、标记数量多、共显性的分子标记,已经被广泛的应用于基因定位及克隆、亲缘分析或品种鉴定、农作物育种、进化研究及分子身份证构建等研究[15-17]。DNA分子身份证的构建是基于DNA指纹图谱,运用多种编码数字化处理电泳图谱获得字符串,并辅以条形码或二维码等科学表述[18]。与聚丙烯酰胺凝胶电泳相比,荧光毛细管电泳提高了数据结果的准确性、减少了操作者的主观实验误差、提高了检测效率[19]。张雅楠等[20]利用14对EST-SSR分子标记,分析了额济纳湿地12个天然多枝柽柳种群的遗传多样性和遗传结构,结果表明所调查的多枝柽柳种群遗传变异信息丰富、遗传多样性较高、基因交流频繁,且种群内的遗传变异明显高于种群间,种群间的遗传距离与地理距离无相关性,为额济纳湿地多枝柽柳种质资源的管理、保护和利用提供了理论依据。陈小红等[21]利用35个高基元SSR构建了来源于4个生态栽培区的130份黍稷种质资源的分子身份证。赵路宽等[22]利用SSR分子标记技术对99份甘薯品种材料间的亲缘关系及遗传结构进行探究,揭示了不同品种个体间的遗传背景和基因渗透情况。
在黄连木的SSR分子标记相关研究中,吴志庄等[23]采用SSR分子标记技术对8个居群进行遗传多样性分析,揭示了黄连木天然居群在遗传多样性水平上的差异及遗传分化状况。郝丽娟[24]应用SSR分子标记技术对黄连木的遗传距离与地理位置进行研究,结果表明遗传距离在省份、经纬度方面基本趋于一致,二者存在密切的相关性。阮福娜等[25]应用SSR分子标记技术对140份黄连木样本进行亲缘关系和居群遗传结构分析,结果表明在同一棵雌雄同株树上,不同性别表现的花序,其SSR基因型是相同的,即花序性别类型的改变并未造成本身基因型的改变;而不同的株雌雄同株个体其SSR基因型是不同的。杨子玥[26]通过SSR分子标记技术鉴定雌雄同株黄连木子代的父本,选择出雌雄同株黄连木适宜的授粉组合,提出了利用 DNA 亲子鉴定确定适宜授粉组合的新方法。在黄连木的SSR分子标记相关研究中,存在使用引物质量较低、电泳结果分辨率不高以及取样地范围较小等问题,不能较为全面地反映全国分布的黄连木遗传分化及遗传多样性等问题。因此,为了解黄连木在中国的分布状态及不同地区种质资源的亲缘关系,评估各地区种源间的遗传多样性和群体遗传结构,实现分子层面的种质资源鉴定和保护,本研究在黄连木种质资源全国调查的基础上,选取具有代表性的采样区进行SSR分子标记试验,利用毛细管电泳方法提高试验准确性,并尝试建立黄连木DNA分子身份证体系。
1 材料和方法
1.1 试验材料
在黄连木全国分布区展开调查的基础上,对其中20个省、市、自治区的29个黄连木分布区进行了调查收集(表1),基于调查并初筛优质资源后进行采样,供试材料分布广泛且代表性强。每份材料采取若干幼嫩叶片,干燥处理后保存至-80 ℃超低温冰箱备用。将29个黄连木居群按照其地理位置大致区划为华北地区、华中地区、华东地区、秦巴山区、西南地区和东南沿海地区等6个区域。
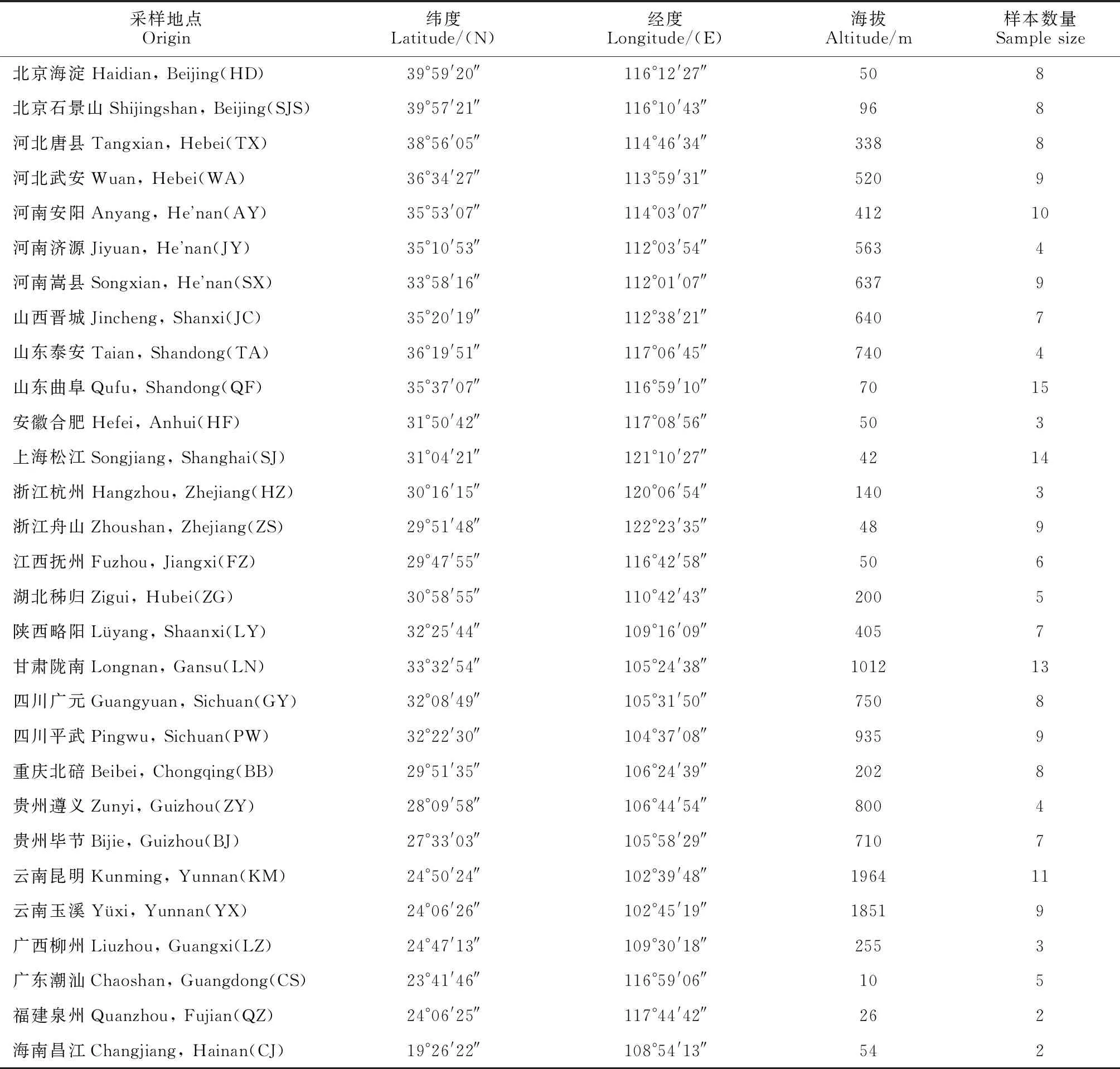
表1 黄连木29个居群信息
1.2 试验方法
1.2.1 基因组DNA提取黄连木基因组DNA的提取使用擎科植物基因组提取试剂盒,用紫外分光光度计(NANODROP 8000)和1%的琼脂糖凝胶电泳检测所提DNA的浓度和质量。检测合格的DNA样品稀释至30 ng/μL置至-20 ℃冰箱备用。
1.2.2 PCR扩增体系聚丙烯酰胺凝胶电泳PCR扩增采用20 μL体系:1 μL模板DNA(30 ng/μL)、10 μL 2×Taq PCR MIX、正、反引物各0.5 μL(10 μmol/L)、8 μL ddH2O补齐至20 μL体系。
荧光毛细管电泳PCR扩增采用20 μL体系:1 μL模板DNA(30 ng/μL)、10 μL 2×Taq PCR MIX、0.1 μL M13正引物(10 μmol/L)、0.3 μL反引物(10 μmol/L)、0.2 μL荧光标记(FAM,HEX,ROX 或 TAMRA)的M13引物、8.4 μL ddH2O补齐至20 μL体系。
反应程序均为:94 ℃预变性3 min;94 ℃变性30 s,51~58 ℃退火30 s,72 ℃延伸30 s,35个循环;最后72 ℃延伸10 min,4 ℃保存[27]。
1.2.3 引物筛选在阮福娜[28]所筛选的14对黄连木SSR引物及卢剑波等[29]公开发表的12对黄连木微卫星位点及引物专利基础上,进一步通过非变性聚丙烯凝胶电泳试验在全部26对引物中筛选出条带清晰、重复性好、多态性高的引物用于全样品的毛细管电泳分析。引物由上海生工生物工程技术有限公司合成。
1.2.4 毛细管电泳每次将4组不同荧光标记的PCR扩增产物混样送测至上海生工生物工程技术有限公司进行毛细管电泳检测。电泳结果使用Gene Marker v2.2.0软件(Soft Genetics LLC,PA,USA)读取等位变异片段大小信息。
1.2.5 数据统计分析使用 Power Marker V3.25[30]软件对不同引物扩增的各样品基因型数据统计主要等位基因频率(MAF)、等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)、基因多样性(GD)、Shannon’s多样性指数(I)和多态信息含量(PIC)等信息。通过GenAlEx 6.502[31]计算种群间的遗传分化固定指数(Fst)、主坐标分析(PCoA)和分子方差分析(AMOVA),利用这些数据评估各种群间的遗传分化和遗传变异特征。
1.2.6 DNA分子身份证构建利用筛选出高质量SSR引物,对全部供试样品进行荧光毛细管电泳,根据毛细管电泳结果来构建黄连木种质资源单株DNA分子身份证。将选用的SSR引物按照固定顺序,每个样品在对应的引物扩增出来的目的片段的大小按照引物顺序直接编码成字符串即为黄连木单株DNA分子身份证,再将单株种质资源的采样地、编号及身份证号等信息导入二维码生成器中即可得到二维码形式的DNA分子身份证。
2 结果与分析
2.1 SSR分子标记引物对的筛选
利用部分样品采用非变性聚丙烯酰胺凝胶电泳方法,对全部的26对黄连木SSR分子标记引物对进行筛选(图1),共筛选出7对条带清晰、多态性高、重复性好的高质量SSR引物(表2),并将它们用于全部黄连木样品的荧光毛细管电泳试验。

表2 筛选出的7对SSR引物序列信息

M.DL500;1-25. KL007、KL008、KL009、KL010、KL011、KL012、KL013、LGC001、LGC002、LGC003、LGC004、MCC001、MCC002、MCC003、MCC004、WJC001、WJC002、WJC003、WJC004、WJC005、MAS001、MAS002、MAS003、MAS004、MAS005
2.2 SSR分子标记多态性分析
对选取的7对高质量SSR引物,利用荧光毛细管电泳方法对210份黄连木种质资源进行分析,部分样品毛细管电泳结果见图2。表3显示,7对引物在210份种质中共扩增出158个等位基因位点,平均每对引物的等位基因数为22.571个。基因多样性(GD)变化幅度为0.654~0.913,平均为0.804;期望杂合度(He)变化范围0.257~0.771,平均为0.532;多态信息含量(PIC)变化范围0.639~0.907,平均为0.784。7对引物的多态信息含量(PIC)均高于0.5,具有较高的多态性,可用于构建DNA分子身份证[32]。
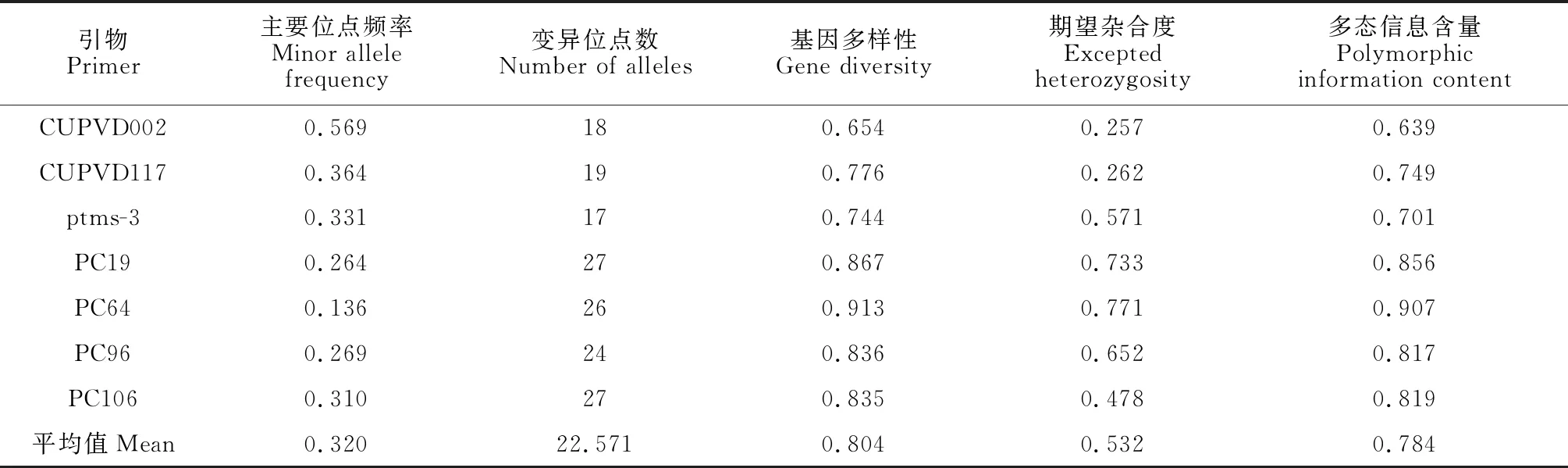
表3 7对SSR引物的检测数据
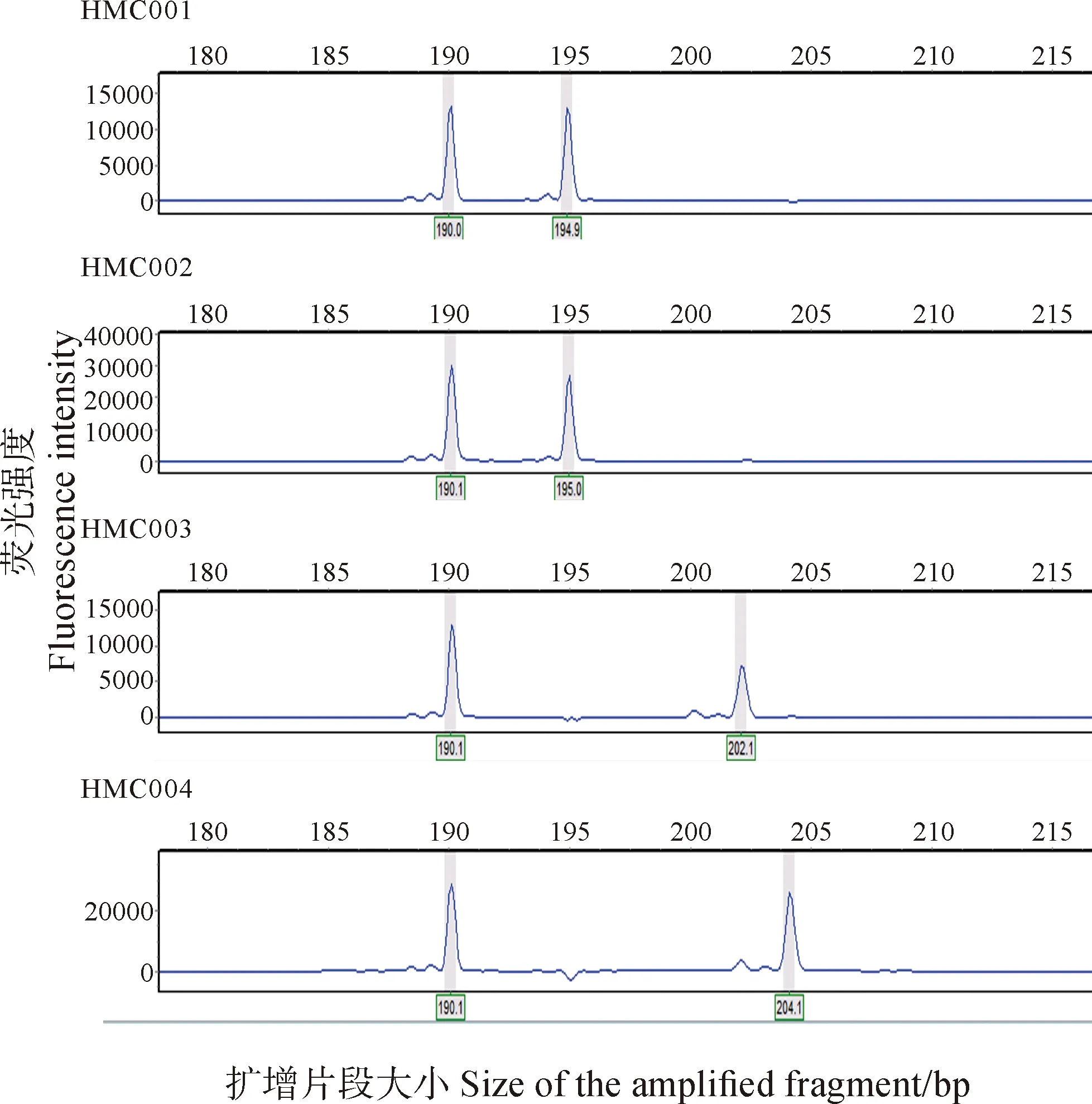
图2 采用引物PC96对部分样品的扩增结果
2.3 聚类分析
基于UPGAM方法对29个采样地的210份黄连木资源进行聚类分析,结果(图3)表明,全部采样地被分为2个大类,西南地区的6个采样点单独聚为Ⅰ 类,其他地区采样点聚为 Ⅱ 类,而其中东南沿海的4个采样点在 Ⅱ 类里相对较为集中,聚集为一小类。
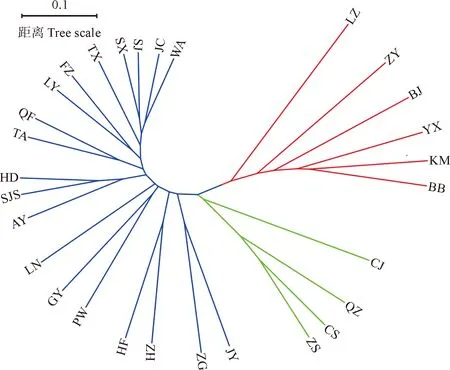
红色为组群Ⅰ;蓝色和绿色为组群Ⅱ;图中采样点“同表1”
2.4 群体间遗传多样性
黄连木作为中国的乡土树种,虽然在中国有着较为广阔的分布空间,但并未形成较大的遗传分化,也鲜有品种的驯化和审定。故本研究在全国较大范围采样的基础上,参考聚类分析的结果,将采样地根据中国的地理分区,大致区划为6个群体分布区进行相关研究,分别为华北地区、华中地区、华东地区、秦巴山区、西南地区和东南沿海地区(图4)。

图4 黄连木中国的分布图及采样点信息
分析不同地区黄连木群体的遗传多样性数据可知(表4),观测杂合度(Ho)介于0.373~0.600之间,平均值为0.520;期望杂合度(He)介于0.632~0.811之间平均值为0.737;均高于0.500,表明黄连木在物种水平上具有较高的遗传多样性。变异位点数(Na)、Shannon’s多样性指数(I)均为秦巴山区>华东地区>西南地区>华北地区>华中地区>东南沿海地区。
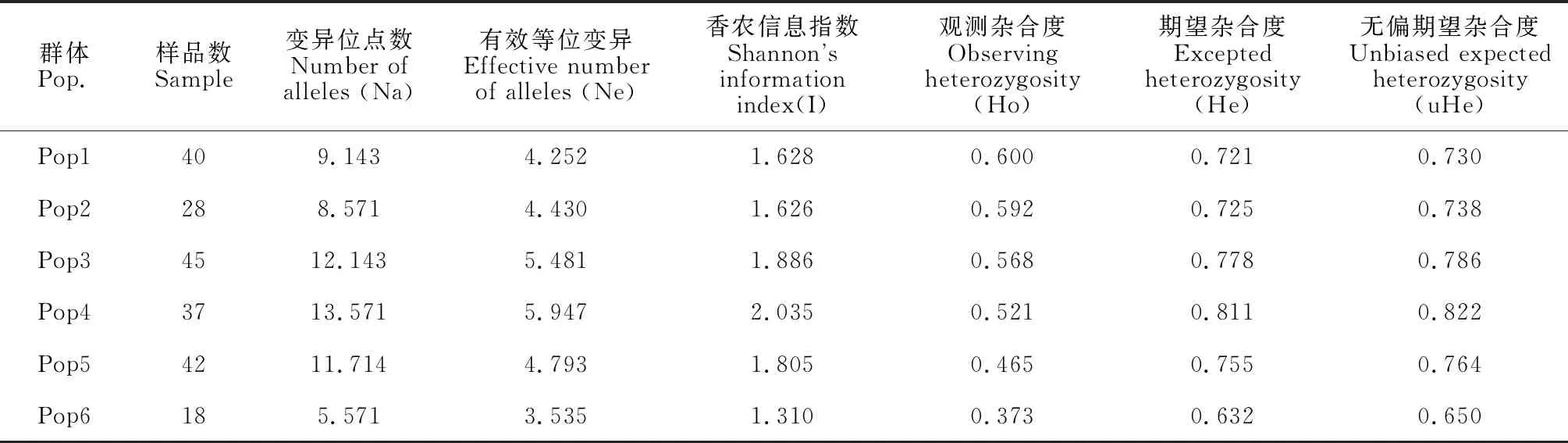
表4 不同地区黄连木群体遗传信息
2.5 群体遗传分化与变异分布
Fst固定系数是反映群体等位基因杂合性水平、用于衡量种群分化程度的指标,其取值范围在0~1之间。通过计算不同群体间的Fst固定系数可知(表5),黄连木各地区群体间的遗传分化值在0.015~0.099之间,并不存在较高的遗传分化。其中西南地区与华北、华东地区,东南沿海地区与华北、华中、华东、西北、西南5个地区的黄连木群体遗传分化值在0.05~0.15之间,表现出中等程度的遗传分化。而其他各地区群体之间的遗传分化值均小于0.05,表明这些区域间只存在较低的遗传分化,具有较近的亲缘关系。
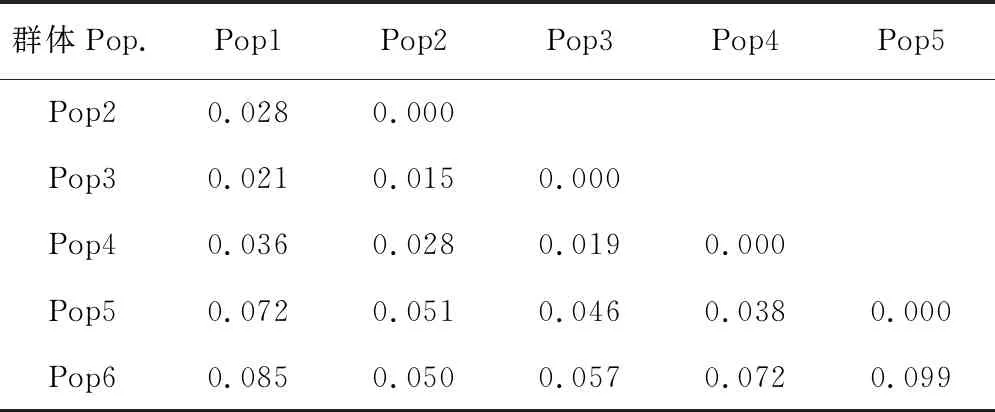
表5 各地区群体间的Fst值
对各地区黄连木群体进行分子方差分析以确定群体间位点的变异总量及在群体间和群体内的变异分布(表6)。结果表明,群体间的变异占总变异量的6%,群体内的变异占总变异量的94%,变异主要发生在群体内,群体内的基因交流明显大于群体间基因交流。

表6 各地区黄连木群体间的AMOVA分析
2.6 群体结构分析
通过对210份黄连木资源进行群体遗传结构分析后发现,当K=2时,ΔK出现峰值(图5),将全部调查资源分为2个组群(图6),其中绿色组群Ⅰ主要为黄连木西南地区群体,而红色组群Ⅱ则为其他5个调查区群体。可见黄连木群体分析结果与聚类分析结果也是相一致的,西南地区黄连木较全国其他地区黄连木有着相对较远的亲缘关系和相对较大的遗传分化与变异,成为中国黄连木自然分布区内一个较为独立的群体。

图5 各群体黄连木群体结构分析的ΔK值分布
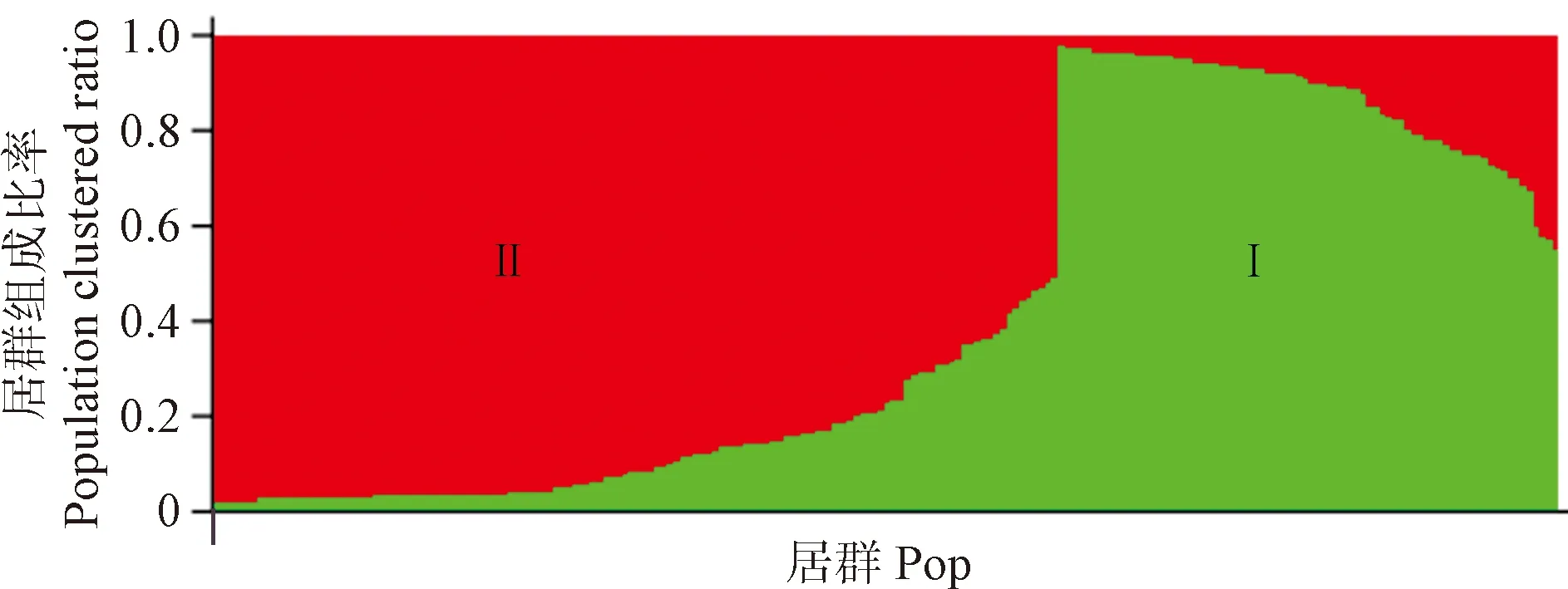
K=2时Structure分组结果,图中同一颜色表示属于同一分支;绿色代表pop5,红色代表pop1、pop2、pop3、pop4和pop6
2.7 主坐标分析
对全部黄连木资源进行主坐标分析(PCoA),结果可知(图7),全部资源除西南地区和东南沿海地区大致聚为相对集中的组群外,其他地区的黄连木资源相互掺杂、重叠较多,分组不明显,共同聚集为一个大组。这一结果也与调查地区黄连木的聚类分析和群体分化与变异结果一致,同时也证明了中国黄连木暂未形成较大的遗传分化,全国范围内的黄连木种群均具有较近的亲缘关系。

图7 各群体黄连木主坐标分析图(PCoA)
2.8 DNA分子身份证构建
对于选择的7对高质量SSR引物,可将全部供试样品区分开,故而可以用来构建DNA分子身份证。将选用的7对SSR引物按照CUPVD002、CUPVD117、ptms-3、PC19、PC64、PC96、PC106的固定顺序,每个样品在对应的7对引物扩增出来的目的片段的大小按照引物顺序编码成的字符串即为DNA分子身份证,在对应位置上,相同的代码表示该引物在样品中扩增出单个条带。每对引物在样品中扩增出的目的条带大小均在110~400 bp之间,因此每对引物均为两个3位数的字符串编码,未扩增出条带的则以两个0编码,最后以“g”终止,即为一个完整的分子身份证。最后将单株的采样地、编号及身份证号等信息导入二维码生成器中以得到二维码DNA分子身份证进行科学保存。
以其中来自北京海淀地区编号为ZWS001的样品为例,其在该体系下构建的分子身份证号为:267267174174158160262274180190190194228228g,表示该样品在引物CUPVD002扩增条带为267/267,在引物CUPVD117扩增条带为174/174,在引物ptms-3扩增条带为158/160,在引物PC19扩增条带为262/274,在引物PC64扩增条带为180/190,在引物PC96扩增条带为190/194,在引物PC106扩增条带为228/228,最后以“g”结束编号。下图为部分样品的DNA分子身份证二维码(图8)。

图8 部分样品DNA分子身份证二维码
3 讨 论
3.1 中国黄连木种质资源遗传多样性分析
遗传多样性水平是由进化历史事件、分布区大小、生物学特性和生态条件以及人为干扰等因素综合作用的结果[33],黄连木在中国广泛的分布区和古老的栽培史也使得其拥有着丰富的遗传基础。遗传多样性是生命进化和物种分化的基础,任何一个物种或生物个体都蕴涵着大量的基因,它所包含的基因变异越丰富,对环境的适应能力就越强,进化的潜力也越大。作为一个极具开发潜力、兼具观赏、绿化、药用、油用的多功能树种,关于黄连木遗传多样性的研究并不多。王超等[34]利用随机扩增微卫星多态性(RAMP)对中国6个地区的180份黄连木样品进行遗传多样性研究,结果表明群体内的基因分化较高,是变异的主要来源,群体间的基因流为1.7633,也表明各个群体间基因交流较为顺畅。吴志庄[35]利用简单重复序列(SSR)对8个黄连木天然种群遗传多样性进行研究,由遗传距离将8个居群聚为3大类,各居群的遗传多样性处于中等水平。整体来看,近10年来中国关于黄连木遗传多样性的研究较少且涉及地域范围不大,对黄连木种质资源的开发利用和保护工作仍有待进一步加强。
本研究利用7对高质量的黄连木SSR引物,采用荧光毛细管电泳方法,对中国29个采样点的210株黄连木资源进行遗传多样性分析。7对引物共检测到158个等位基因,平均每对引物有22.571个等位变异,明显高于吴志庄和郝丽娟等使用聚丙烯凝胶电泳方法的检测结果,说明利用SSR能检测到黄连木丰富的遗传变异。多态信息含量(PIC)变化范围0.639~0.907,平均为0.784,表明供试材料具有较高的多态性,其原因可能是本研究在全国进行了大范围的调查采样。
在遗传分化方面,本研究通过UPGMA聚类、PCoA分析和群体遗传结构分析对黄连木种质资源进行划分,3种分析方法得到的结果较为一致,210份种质被划分为两大类,西南地区群体单独为一类,其他地区单独为一大类。种群间遗传分化值在0.015~0.099之间,表明黄连木各地区群体间存在中等以下程度的分化,西南地区和东南沿海地区黄连木群体与其他群体有相对较大的遗传分化。这表明在全国范围内中国黄连木亲缘关系较近,不同群体间相似程度高,并未形成明显的遗传分化,这可能与黄连木在中国开发利用仍处于起步阶段,无性繁育和人为选择少、功能应用较为单一有关。此外分子方差分析(AMOVA)可知,黄连木各地区群体内的遗传变异占94%,群体间的变异占6%,这说明黄连木群体间遗传分化较小,遗传分化主要存在于群体内,这也与管菊[36]和李琬婷等[37]在河南地区黄连木居群的研究结论较为一致。
在黄连木群体间遗传多样性方面,对不同地区黄连木群体间的关键遗传数据分析可知,变异位点数(Na)、Shannon’s多样性指数(I)均为秦巴山区>华东地区>西南地区>华北地区>华中地区>东南沿海地区,其中秦巴山区、华东地区、西南地区具有较高的遗传多样性,有利于黄连木优良品种的选育和遗传改良。在较高遗传多样性的3个地区中,华东地区可能是由于采样地包含合肥大蜀山植物园、上海辰山植物园、曲阜鼓楼北街行道树等人工移栽苗木较多有关,存在不同群体间种源调拨的过程。秦巴山区黄连木群体的变异位点数(Na)、有效等位变异数(Ne)、Shannon’s多样性指数(Ⅰ)均为各地区最高,是调查区遗传多样性最丰富的地区,其原因可能是四川广元剑阁翠云廊自然保护区作为秦巴山区群体的主要采样点,拥有着较为集中分布的黄连木古树群落,进而使得该地区具有较高的遗传多样性。而与秦巴山区地理位置相邻的西南地区也同样表现出较高的遗传多样性水平,该地区的采样点也多以天然古树群落、散生古树单株、黄连木天然林等为主。其聚类结果也单独聚为一类,遗传距离相对较远,表明西南地区黄连木群体相对独立且有极大可能为中国黄连木的起源所在,是黄连木种质资源遗传多样性和遗传分化的核心区。
从大尺度群体进化角度看,秦巴地区和西南地区作为秦岭南侧,横断山脉北麓的中间区域,对植物的生存有着得天独厚保障,也极有可能是第四纪冰川期重要的“植物避难所”。这也与中国近些年开展的第四纪冰期生物避难所的研究相契合。
第四纪冰期的低温导致适应了温带和热带环境的物种迁移到避难所生长,成为它们新的生存中心。由于高大山脉的保护作用,秦巴山区独特的地理条件缓冲了第四纪冰期冰川低温对植物造成的破坏。再加上四川盆地、云贵高原和众多河谷的良好水热组合,形成了冰川时期的保护区,成为了亚热带植物区系扩散和分化的核心地区[38]。植物避难所具有独特的地理条件、适宜的生存环境和丰富的生物资源,因此,植物避难所的确定也可以为生物多样性保护区域的划分提供依据,有助于生物多样性的研究和有效保护[39]。研究黄连木种质资源的遗传多样性,对于探索其遗传多样性的丰富程度,掌握其特点和分布规律,促进黄连木种质资源的开发利用与保护、育种亲本的合理选择和优良基因的定位等方面都具有重要的意义[40]。
3.2 中国黄连木种质资源DNA分子身份证的构建
黄连木作为中国极具发展前景的优良乡土树种,其种质资源的调查与保护工作将为其未来的开发利用提供良好的基础与支撑。DNA分子身份证作为一种直观且方便快捷的种质资源鉴定方法,能有效地区分单株,是种质资源保护与开发利用的有力手段[41]。本研究利用筛选出的7对黄连木SSR引物,可将全部210份样品鉴别出,形成唯一指纹图谱,进而将DNA指纹数字化,得到每份材料的DNA分子身份证。由于其遗传物质直接以DNA的形式表现,其差异体现的是不同的基因型差异,因而更加准确、客观。在过往的研究中,虽然DNA分子身份证技术在诸多树种中有广泛的应用,但在黄连木的相关研究中却鲜有报道,本研究利用等位基因赋值编码方法,将每对引物扩增的等位基因按从小到大顺序依次编码,获得每份资源的DNA分子身份证。借鉴DNA分子身份证技术,为黄连木种质资源“身份”的精准鉴定提供参考,也为其种质资源的收集、利用、和保护奠定基础。
从公开发表的26对黄连木SSR引物中筛选出7对重复性好、多态性强、条带清晰的引物进行黄连木遗传多样性分析,遗传多样性分析结果表明在全部采样范围内,秦巴山区、华东、西南地区黄连木遗传多样性较高。遗传分化结果表明,全国范围内黄连木遗传分化不大且主要遗传分化主要存在于群体内。用SSR分子标记结果创建了黄连木DNA分子身份证,为标准化管理黄连木种质资源提供参考。
