乳酸菌产γ-氨基丁酸及生物合成技术研究进展
2022-11-07李朔李潇张晓黎贾瑶吴兴壮
李朔,李潇,张晓黎,贾瑶,吴兴壮*
(1.沈阳农业大学 食品学院,沈阳 110866;2.辽宁省农业科学院 食品与加工研究所,沈阳 110161)
γ-氨基丁酸(γ-aminobutyric acid, GABA)是一种重要的非蛋白氨基酸,具备多种用途。研究表明,GABA不仅在农业中被用于调节植物生长、发育和应激反应,还在化学工业中作为前体物质用于合成可生物降解尼龙[1]。同时,作为哺乳动物中枢神经系统的主要抑制性神经递质,GABA对健康的益处已被充分证实,其在治疗高血压、缓解压力和预防糖尿病等方面起到了良好的治疗效果。鉴于GABA在食品和医药领域具有重要用途,我国卫生部于2009年正式批准GABA为新资源食品的生产原料。近年来,人们将其作为生物活性因子,已开发出富含GABA的酱油、豆豉、食醋、豆酱、海鲜调味料等调味品以及发酵腌制品和奶制品[2]。然而,GABA在自然生物中浓度很低且不易提取,因此,为了实现GABA的有益功能,GABA的高效合成受到了极大关注。
谷氨酸脱羧酶(GAD)是生物体催化谷氨酸或钠盐经过脱羧生成GABA的唯一关键酶。乳酸菌(LAB)作为一种公认安全的细菌被广泛应用于食醋、腐乳、泡菜等食品生产中[3],已有许多科学研究证实,乳酸菌存在谷氨酸脱羧酶活性,能够催化谷氨酸脱羧生成GABA。目前,GABA主要通过生物合成法来生产,如在生物转化方法中使用分离的GAD或在微生物直接发酵法中利用具有GAD活性的各种微生物菌株,这是因为该法具有反应程序简单、催化效率高、反应条件温和以及环境兼容性等优点[4]。因此,通过利用食品安全级乳酸菌生物合成达到更高GABA生产目标、开发更多富含GABA种类的功能性食品正在吸引着世界各地的科学工作者。
1 γ-氨基丁酸的生理作用
GABA作为哺乳动物的一种抑制性神经递质,起到分子信号的作用,并具有许多生理功能。科学研究已经证实,GABA不但有降低神经元活力、避免神经细胞过热、降血压的功效,还可作为肝、肾和肠道的保护剂以及预防毒素引起的损伤,同时GABA对癫痫发作、惊厥、帕金森病等多种精神疾病都有一定的治疗效果[5]。虽然GABA有多种生理功能,但由于人们压力的增加和年纪的增长,身体中的GABA含量也日益减少。因此,在饮食中补充GABA对人体保健具有很重要的意义。
GABA对生物体的影响主要通过其与相应的受体融合而产生。
GABA受体一般有3类:(1)GABAA受体主要分布在神经元细胞膜上,且具有高度敏感性。其被激活后使得突触后电位发挥作用,抑制神经元的兴奋,保持大脑自我调节。因此,调节不平衡也可引起精神疾病、创伤后应激功能障碍 (PTSD) 综合征、癫痫等病症,此外,与女性生殖周期及相关抑郁症发作也有着密切的关系[6]。(2)GABAB受体主要产生在神经元的突触前、后部位,且在非突触部位也有其存在。其受体被激活后,通过突触后膜使得K+形成输出电导,从而形成长抑制性突触后电位,相反,突触前膜则抑制钙离子电导,使兴奋性或抑制性递质的释放量降低,另外,该受体被激活后还与消化系统功能调节等许多作用有关[7]。(3)GABAC受体和GABAA受体相似,主要存在于视网膜双极细胞中,另外也有研究表明,其在视网膜中视杆通路的信息传导与调控中起重要作用[8]。
2 生产GABA乳酸菌的种类
乳酸菌是各种食品原料发酵过程中最重要的微生物之一,乳酸则是在发酵过程中形成的主要代谢产物之一,影响乳酸菌的生理活动。许多原料或食品中都含有大量的谷氨酸,乳酸菌则可以利用谷氨酸来提高其对酸性条件的耐受性,这是因为在酸性条件下,一些乳酸菌启动了不同的耐酸系统来维持细胞活力,如生产GABA则是利用了其体内的GAD抗酸系统[9]。GAD是一种依赖5′-磷酸吡哆醛(PLP)的细胞内酶,其催化谷氨酸向GABA的转化,需要通过与H+结合将底物脱羧成中性化合物GABA,进而保护乳酸菌不受酸胁迫的伤害[10]。目前,许多生产GABA的乳酸菌已从广泛的发酵食品中分离出来,包括奶酪、酸奶、酸菜、泡菜、发酵香肠以及各种发酵鱼制品[11]。生产GABA乳酸菌的起始菌株是影响GABA生产的最主要原因,具备合成GABA能力的乳酸菌较多,乳酸杆菌属也被列为生产GABA的最主要种类,包括短乳杆菌、副干酪乳杆菌、保加利亚乳杆菌、布氏乳杆菌、植物乳杆菌、瑞士乳杆菌等,而在这些具备产GABA能力的乳酸菌株中,异型发酵的短乳杆菌是目前研究最佳的菌株,在适当条件下可生产大量的GABA[12]。
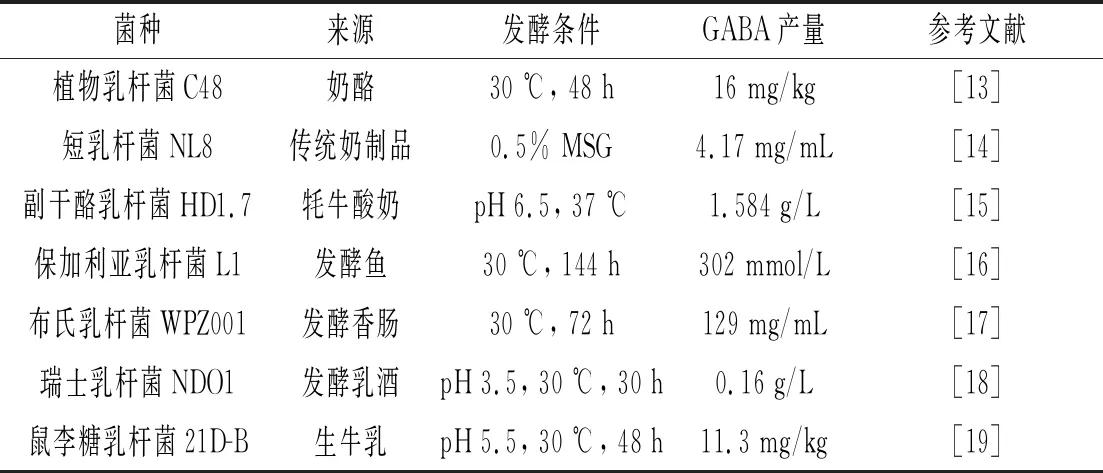
表1 不同乳酸菌的发酵特性
3 提高GAD活性和GABA产量方法
3.1 通过优化培养基及发酵条件提高GABA生物合成产量
菌株间生成GABA的能力差异较大,同时菌株也受到碳氮源、底物浓度、起始pH值、培养温度、培养时间和辅酶因子等因素的影响。因此,通过优化实验条件可以提高菌株生产GABA的效率。周青[20]的研究表明,在优化碳氮源及添加量的条件下,植物乳杆菌WZ011能产生1.261 g/L的GABA。谷氨酸钠(MSG)或L-谷氨酸是使用纯化酶或产GABA能力的菌株来生产GABA的主要合成底物,其浓度能够控制GAD的活性,进而影响GABA的产量[21],添加0.5%的MSG增加了屎肠球菌JK29的GABA产量,培养48 h后达到14.9 mmol/L。当L-谷氨酸以10~20 g/L的浓度添加到嗜热链球菌的生长培养基中时,不能有效增加GABA的产量,表明该菌株不能耐受高浓度的L-谷氨酸,这是因为高浓度的底物会增加细胞渗透压,干扰细菌代谢,影响GAD的活性[22]。因此,增加底物浓度的策略不能用于全部菌株。因为PLP是GAD必需的辅酶,所以产生GAD活性的乳酸菌还必须在培养基中添加PLP以增加GABA的产量,Komatsuzaki等[23]研究证实,将PLP添加到初始培养基中可以极大地促进副干酪乳杆菌的GABA生成。但是Yang等[24]研究表明,在初始发酵时加入PLP无法有效地提高唾液链球菌嗜热亚种GABA的产生,然而在发酵进行48 h后,添加PLP在提高GABA生产方面却表现出了有效的功能,推测这或许是游离PLP在培养过程中很容易变性,失去了作为GAD辅酶的作用,因此补充PLP可以部分恢复GAD活性,增加GABA产量。发酵时间和温度也是影响菌株生产GABA的关键因素,温度过低或过高都会抑制菌株的生长速度和产GABA能力,一般来说,在25~45 ℃范围内,菌株产生GABA的含量最高[25]。Villegas等[26]通过短乳杆菌CRL1942研究了GABA的形成,结果发现,在30 ℃条件下使用270 mmol/L的MSG发酵48 h,MRS培养基中GABA的最大产量为255 mmol/L,表明GABA的产生有时间依赖性。这是因为由MSG向GABA转化需要菌株生长,产生较高的细胞密度和合适的培养温度[27],另外,乳酸菌在发酵过程中会形成乳酸,导致发酵液pH下降,当发酵液pH降至GAD活性范围时,培养基中GABA才开始出现。微生物中GAD的活性主要由pH值调节,pH值的升高或降低会造成GAD活性部分失活,从而影响GABA产量,由于各菌株GAD生化特性有所不同,所以不同菌株之间的GAD活性最佳pH值也不相同,总体来说,GAD的最适pH值为4.0~5.0[28]。例如,pH为5.0时,布氏乳杆菌产生最多的GABA,产量达251 mmol/L;pH为6.5时,植物乳杆菌IFK10产GABA最多,达2.68 g/L。
3.2 采用补料分批发酵技术提高GABA生物合成产量
各菌株产生GABA的能力差异很大,一些产生GABA的菌株在大规模发酵生产GABA方面显示出巨大的潜力。由于发酵的主要目的是成本效益以及提高生产生物产品的效率,所以选择适当的发酵工艺来提高目标产品的产量非常重要。在分批发酵时,底物通常在发酵罐中只放置1次,产生的主要问题是较高的发酵底物初始浓度会抑制细胞生长,而较低的底物浓度则无法适应高生产的要求[29],然而补料分批培养能够克服这一问题,已广泛应用于各种生物制品的生产中,在补料分批培养过程中,可选用不抑制细胞生长的适宜的初始底物浓度,随着发酵时间的延长,一个或多个营养成分被供应到发酵罐中,产物则留在发酵罐中直到发酵结束,这种方法可以获得高产量的目标产物。Li等[30]设计了一种简单的分批补料发酵方法,当菌株生长至对数生长期时,在温度32 ℃、搅拌速度100 r/min、pH 5.0、发酵时间48 h的条件下进行补料发酵,且在12 h和24 h分别向发酵罐中加入280.70 g和224.56 g的谷氨酸,结果显示,48 h 后GABA浓度达到(1005.81±47.88)mmol/L。由于GAD主要在酸性条件下具有活性且发酵液pH值会随着时间的变化而变化,所以应及时调整pH值,使菌株在最适宜pH值下生产GABA。李胜男[31]通过恒定pH分批补料发酵建立了高效合成GABA的发酵工艺体系,在恒定pH 5.0条件下,GABA的产量由不控制pH的30.34 g/L增加到44.73 g/L,增长了47.43%,且发酵后期的培养液中仍保持着较高的细胞浓度,另外发现与未控制pH相比,发酵液中的葡萄糖以及底物几乎被耗尽,表明恒定pH分批补料发酵有利于提高GAD的活性,进一步提高GABA的产量。
3.3 复合菌培养提高GABA生物合成产量
复合菌培养旨在提高GABA的产量。不同菌株可以相互利用各自的发酵特性,克服单一菌株发酵的缺点,已在食品、农业、医药等行业中得到广泛应用[32]。Kim等[33]将产GABA的短乳杆菌GABA 100与不产GABA的双歧杆菌BGN4复合培养发酵天麻,培养过程中观察到的GABA产量高于只接种短乳杆菌GABA 100,在报道中,并未阐明复合培养对GABA产生影响的确切机制,推测双歧杆菌BGN4在复合培养过程中降低pH值的幅度大于短乳杆菌GABA 100单独培养,从而导致GABA产量增加。张恕铭等[34]通过植物乳杆菌BC112与屎肠球菌AB157复合培养并优化试验条件提高了GABA产量。Kwon等[35]以水芹菜为主要原料,采用肠膜明串珠菌与植物乳杆菌复合发酵,并通过优化增加了水芹中GABA的含量,成功生产了这种同时富含葡聚糖和GABA的新型功能性水芹。
另外,微生物复合培养技术不仅有利于食品生产,而且有利于人类健康,这是因为动物肠道内存在大量的微生物,不同的微生物通过合作执行重要的生物学功能。然而,人们对复合培养机制的了解很少。因此,不同微生物复合培养的机理有待探索,研究结果可推广到工业应用和改善人类健康方面。
3.4 实施现代生物技术提高GABA生物合成产量
研究表明,GABA是利用生物体内的谷氨酸脱羧酶作为催化剂对谷氨酸进行脱羧反应而产生的,许多细菌的GAD在pH为4.0~5.0时表现出最优活性,而在近中性pH时,活性急剧下降。在大多数情况下,直接发酵法的GABA产量相对较低,前期的研究者主要通过优化培养基组成和培养条件来提高GABA含量,因此,GABA的生物合成通过基因工程、代谢工程以及纯化或固定化GAD等生物技术的研究越来越受到人们的关注[36]。
不同来源的乳酸菌的GAD基因已在不同宿主中过度表达,包括大肠杆菌、谷氨酸棒杆菌以及枯草芽孢杆菌[37],许多乳酸菌GABA合成的效率基本上取决于GadC和GadB的活性,而GadC和GadB的活性受到培养条件的极大影响。Tajabadi等[38]为了提高GABA的合成,将获得的GAD基因克隆到pMG36e-LbGAD载体中,并且通过SDS-PAGE和GAD活性证实了在植物乳杆菌Taj-Apis362 细胞中进行过度表达,与原始GAD活性相比,其酶活性提高了7倍,达到了53 kDa蛋白。Yang等[39]将基于大肠杆菌表达乳酸乳杆菌GadB基因的全细胞生物催化剂用于代谢途径工程方法的起点:(1)通过向GadB中引入突变,使其脱羧活性向中性pH方向转移,构建了工程菌株;(2)通过修饰谷氨酸/GABA逆向转运蛋白GadC,促进中性pH下的转运;(3)通过过度表达GroESL分子伴侣,增强可溶性GadB的表达;(4)通过使大肠杆菌基因组中的gadA和gadB失活来抑制GABA的降解,最终该工程菌株每小时产GABA量为44.04 g/L。Liu等[40]开发了一种乳酸链球菌发酵系统,可以同时生产乳酸链球菌肽(一种抗菌肽)和 GABA(一种抗氧化剂),在此研究中通过表达谷氨酸脱羧酶和谷氨酸/GABA逆转运蛋白代谢工程改造了乳酸链球菌素生产菌株L.lactisF44用于 GABA 生产,GABA 生物合成可以通过增强菌株的耐酸性来促进乳酸链球菌素的产生,通过采用两阶段 pH 控制发酵策略,工程菌株GABA产量高达9.12 g/L,是恒定pH值发酵的2.2倍。
一些突变方法,如定向进化和定点突变被认为是优化或改善GAD催化活性的有力工具,并应用于全细胞生物催化。谷氨酸棒杆菌是谷氨酸的工业生产菌,且在中性pH值附近生长最好,Shi等[41]为了在扩大的pH范围内提高短乳杆菌Lb85的GAD活性,通过表达短乳杆菌Lb85 GadB变体获得重组谷氨酸棒杆菌细胞,由于变体是通过结合GadB的定向进化和位点特异性突变来构建的,通过将这些GadB变体引入该生物体,在简单的葡萄糖基培养基上无需外源谷氨酸即可生产GABA,产量高达7.13 g/L,且该重组细胞在较高pH值范围内仍表现出较高的催化能力。GadC对底物运输具备严格的pH依赖性,在pH 6.5及以上时没有可检测到的运输活性,Shin等[42]利用诱变研究了植物乳杆菌GadC的C末端依赖pH的催化作用,通过截短C端区域Ile454-Thr468中的Δ3和Δ11变体,该酶在pH 5~7范围内增加了活性,其中Δ11变体明显显示出更好的结果,在pH值为5.0和7.0时,变体的催化效力分别提高了1.26倍和28.5倍,推测由于pH诱导的构象变化,C端区域通过关闭催化位点,使得植物乳杆菌GAD的活性在较低或较高pH值下增加。Lyu等[43]将短乳杆菌GadC的C末端Δ14变体通过定向突变被截短,使得该变体在较高的中性pH值下表现出更好的活性。这些研究表明,在较高pH值下,GadC的C末端对于改善酶活性位点以及增加活性具有重要影响,因此GadC在波动的pH条件下通过代谢途径工程是产生GABA的重要途径。
热稳定性不足是酶的常见问题,大多数GAD即使在中等温度下也表现出低稳定性。改进热稳定性的一个合理策略是通过序列比对来识别关键区域或氨基酸残基,林玲等[44]从短乳杆菌中获得GAD1407基因,并通过克隆在大肠杆菌中表达,然后将GAD1407的氨基酸序列与拟南芥的GAD1进行比较,发现了一个影响GAD1407催化pH特性的关键位点S307,然后对这个关键位点进行饱和突变和筛选,最后确定S307N扩大了该酶催化的最佳pH范围。另外,鉴定共有序列也可以提高蛋白质的热稳定性,Zhang等[45]对短乳杆菌CGMCC 1306 GAD的基因开发了一种并行策略,通过比较这种嗜热性GAD与同源嗜热性酶的序列和结构,确定影响稳定性的氨基酸残基并获得了两种突变酶,结果表明,两种突变酶的半失活温度分别比野生型高2.3 ℃和1.4 ℃,表现出较高的热稳定性,此外,在60 ℃孵育20 min期间,变体的活性增加了1.67倍。汪钟[46]通过与嗜热GAD进行多序列比对,首先鉴定出了最有可能提高酶热稳定性的氨基酸残基替换位点及类型,然后又计算了这些突变对蛋白质自由能的影响,最后制定了最佳的酶分子改造方法,他推测这种方法是提高GAD热稳定性的有效工具。
研究表明,通过全细胞将谷氨酸生物催化为GABA有一些不足之处,这是因为GABA在中性环境下经GABA转氨酶(GABA-T)和琥珀酸半醛脱氢酶(SSADH)连续催化两步反应后转变成琥珀酸,从而导致分解[47]。另外,全细胞生物催化需要加入一些缓冲液,使得发酵体系保持一定的酸性条件,这种化学缓冲液不利于后期GABA的纯化。由于简化了从不太复杂的反应混合物中对该化合物的下游纯化,因此在生产纯GABA时,使用纯化或固定化技术似乎比全细胞生物催化更经济可行。研究表明,许多固定化技术已应用于生物催化剂的重复使用,例如将GadB固定在海藻酸钙中,然后将其用于生物反应器,将GAD/纤维素融合蛋白固定在纤维素上以及将GAD固定在卡拉胶上[48]。Lin等[49]在补料分批反应器中对固定化GAD的性能进行了评估,结果表明,当培养基中的底物浓度通过添加固体谷氨酸保持恒定时,GABA的产率较高,此外,通过向反应器中添加少量的α-酮戊二酸,避免了催化过程中PLP对琥珀酸半醛的失活反应,从而再生辅助因子PLP,且在反应过程中未观察到酶活性的显著降低。Lee等[50]使用共价偶联多孔硅固定植物乳杆菌GAD,并在酸性和碱性条件下表现出高稳定性,热稳定性得到改善。这些结果表明,当使用(部分)纯化的GAD从谷氨酸生产GABA时,固定化技术为工业应用提供了有利的依据。
4 展望
综上所述,GABA作为一种抑制性神经递质,被视为药物和食品中的生物活性成分,具有促进身体健康的特性。同时生产GABA的乳酸菌也为研发含有GABA的功能性食品创造了有利条件,已受到国内外研究者的重点关注。GAD在GABA的生物合成中起着重要作用,乳酸菌GAD结构信息将有助于酶工程提高GAD的热稳定性以及在广泛的pH范围内提高其活性和催化效率。虽然现在已有研究对部分乳酸菌GAD的结构信息进行了阐明,但是主要集中在对短乳杆菌GAD的研究。然而现代生物技术的发展能够使我们更深入地了解其他乳酸菌的GAD结构性质和催化特性。未来,GABA资源的开发方向之一是通过对GABA产生的分子机制、调控机制以及对产GABA的细胞生理学的深入研究,使GAD在遗传和代谢水平上具有较高的转化效率,从而促进GABA产量的增加。另一方向是开发出不同类型富含GABA的功能性食品,如开发功能性谷薯类食品(面包、馒头、米胚芽、米糠等)、功能性动物性食品(发酵香肠、发酵酸奶、发酵奶酪等)、功能性蔬菜食品(发酵酸菜、绿豆芽等)、功能性调味品(酱油、腐乳、豆瓣酱、食醋、大豆酱等),以期在人们的日常饮食中通过摄入富含GABA的食品来达到对人体的保健效果。
