mTOR和JNK通路调节神经病理性大鼠杏仁核MCP-1表达的作用分析*
2022-11-03李耿章符文红杨冯睿
李耿章,符文红,杨冯睿
(1.邵阳学院附属第一医院麻醉科,湖南邵阳 422001;2.南华大学附属第一医院麻醉科,湖南衡阳 421001)
神经病理性疼痛(NP)是临床难治性疾病,杏仁核活动与疼痛行为密切相关[1-2],抑制杏仁核活动可缓解NP[3]。哺乳动物雷帕霉素靶蛋白(mTOR)是一种丝氨酸/苏氨酸蛋白激酶,其在调控长时程突触可塑性上发挥重要作用[4]。mTOR及其下游效应物被发现表达于疼痛处理主要区域的脊髓背根神经节(DRG)和脊髓背角,有助于伤害性信息的传递和调制。研究发现,c-Jun氨基末端激酶(JNK)是丝裂原活化蛋白激酶(MAPK)家族成员之一,可在神经损伤后的脊髓星形胶质细胞中持续活化,而JNK抑制剂SP600125可改善脊髓损伤后的NP症状[5-6]。本课题组前期研究发现,单核细胞趋化因子-1(MCP-1)参与NP状态下杏仁核功能异常,然而,NP状态下杏仁核中mTOR和JNK表达的变化及其与MCP-1关系尚不清楚,故本研究将进行初步探讨,现报道如下。
1 材料与方法
1.1 材料
1.1.1主要试剂
兔抗MCP-1单克隆抗体(批号:sc-4906,美国Santa Cruz公司),磷酸化mTOR(p-mTOR)多克隆抗体(批号:ab45989,美国Abcam公司),磷酸化JNK (p-JNK) 单克隆抗体(批号:ab124954, 美国Abcam公司),mTOR抑制剂雷帕霉素(批号:HY-10219,美国MCE公司),JNK抑制剂SP600125(批号:420119-5MG,德国Merck公司),MCP-1 ELISA试剂盒(美国RD公司)。
1.1.2实验动物与分组
选取2月龄200~260 g雄性SD大鼠96只,采用左侧第5/6腰椎(L5/6)脊神经结扎(SNL)制备NP模型。实验分为3个部分:(1)将24只大鼠于SNL术前及术后第7、14和21天(d0、d7、d14和d21,各6只)取杏仁核,采用Western blot检测p-mTOR、p-JNK表达情况并分析二者与MCP-1水平的相关性;(2)48只NP模型大鼠双侧杏仁核注射不同浓度mTOR抑制剂雷帕霉素(0、0.1、1.0和10.0 μmol/L)或JNK抑制剂SP600125(0、0.1、1.0和10.0 μmol/L),于术后21 d(每个浓度各6只)取杏仁核,采用ELISA检测MCP-1水平,免疫组织化学法检测MCP-1阳性细胞数;(3)24只大鼠分为3组,即假手术组(仅暴露脊神经而不结扎)、模型组及抑制剂组(双侧杏仁核注射10 μmol/L SP600125或雷帕霉素3 μL),于d0、d4、d7、d14、d21共5个时间点测定各组机械痛阈值(mechanical withdrawal threshold,MWT)。本实验所用大鼠购自上海斯莱克公司,饲养于无特殊病原体(SPF)级实验动物中心,明暗时间设定为12 h∶12 h,保持室温20~22 ℃,湿度60%~70%。
1.2 方法
1.2.1NP模型制备
采用左侧L5/6 SNL制备NP模型,简述如下:经50 mg/kg戊巴比妥钠腹腔注射麻醉后,俯卧位固定在显微外科装置下,取后正中切口暴露第4腰椎(L4)至第3骶椎(S3)的棘突和椎板,小心地移除左侧L6横突,分离左侧L5和L6脊神经并结扎,随后采用6-0丝线连续缝合肌肉和皮肤切口。假手术组大鼠仅暴露L5和L6脊神经,但不结扎。所有大鼠均给予连续5 d青霉素腹腔注射(20万U/d)。剔除术后出现神经功能缺损的大鼠。
1.2.2杏仁核p-mTOR、p-JNK水平检测
分别于d0、d7、d14、d21将大鼠过量麻醉致死,断头取脑,液氮速冻,迅速取出杏仁核,研磨至匀浆,蛋白裂解液提取细胞中的总蛋白,然后采用Western blot检测提取的蛋白水平。具体操作如下:每组提取等量蛋白(添加适量的上样缓冲液),水浴锅加热使蛋白变性,将处理好的蛋白样品(每个胶孔上样量为20 μL左右)进行常规十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),采用湿转法将凝胶上的蛋白转移至聚偏氟乙烯((PVDF)膜。将膜放置于5%脱脂奶粉中封闭2 h,之后用配制好的p-mTOR、p-JNK一抗(稀释比为1︰1 000)在4 ℃冰箱中孵育过夜。第2天加入二抗室温孵育1 h,用TBST洗涤3次后加入电化学发光(ECL)显色液显色,并使用凝胶成像设备进行结果观察和拍照存档。
1.2.3杏仁核MCP-1水平检测
于术后21 d取制备好的杏仁核匀浆液,采用ELISA试剂盒检测MCP-1水平。操作步骤严格遵循试剂盒说明书,在450 nm波长下采用ELx800型酶标仪检测吸光度(A450)值。
1.2.4杏仁核MCP-1阳性细胞数检测
于d21将大鼠过量麻醉致死,经升主动脉灌注4%多聚甲醛500 mL,迅速取出杏仁核,经包埋剂处理后,以20 μm厚度行横向冰冻切片,直接转移至预冷的显微镜载玻片,经煮沸的柠檬酸缓冲液抗原修复和3%过氧化氢消除内源性过氧化物酶活性后,采用稀释比例为1∶200的MCP-1过夜孵育。经相应二抗处理后用二氨基联苯胺(DAB)显色,再经过苏木素复染,镜下观察阳性细胞数目。
1.2.5MWT测定
分别于d0、d4、d7、d14、d21测定各组大鼠的MWT。所有的行为测试均在9:00—14:00进行。MWT参照文献[7]采用序贯法进行测定。将大鼠置于透明的塑料盒中适应至少30 min,采用0.4、0.6、0.8、1.0、2.0、4.0、6.0、8.0、10.0和15.0 g的von Frey纤维丝,初始刺激为2.0 g,垂直刺激左后足跖面,以细丝稍弯曲作为完全受力的标准,每次持续6 s。当出现左后爪急剧缩回或细丝移开立即退缩为阳性反应,依次增加和减少刺激强度,直至出现第1次阳性和阴性(或阴性和阳性)反应的骑跨,再向下连续测定4次。不同刺激之间相隔30 s,以消除前一刺激的影响。最后计算诱发50%大鼠缩足反应的刺激强度即MWT。
1.3 统计学处理
2 结 果
2.1 SNL后不同时间点p-mTOR水平
Western blot检测结果显示,与d0相比,d7、d14、d21NP模型大鼠杏仁核中p-mTOR水平升高(P<0.05),且d21水平最高;进一步分析发现,d21的大鼠杏仁核p-mTOR水平与MCP-1水平呈正相关(r=0.629,P<0.05),见图1。
2.2 SNL后不同时间点p-JNK水平
Western blot检测结果显示,与d0相比,d7、d14、d21NP模型大鼠杏仁核中p-JNK水平升高(P<0.05),且d21水平最高;进一步分析发现,d21的大鼠杏仁核p-JNK水平与MCP-1水平呈正相关(r=0.564,P<0.05),见图2。
2.3 杏仁核注射SP600125对MCP-1表达的影响
与0 μmol/L组相比,0.1、1.0、10.0 μmol/L组大鼠杏仁核MCP-1水平及阳性细胞数均降低,差异有统计学意义(P<0.05),且10.0 μmol/L组大鼠杏仁核MCP-1水平及阳性细胞数最低,见图3、4。
2.4 杏仁核注射雷帕霉素对MCP-1表达的影响
与0 μmol/L组相比,0.1、1.0、10.0 μmol/L组大鼠杏仁核MCP-1水平及阳性细胞数均降低,差异有统计学意义(P<0.05),且10 μmol/L组大鼠杏仁核MCP-1水平及阳性细胞数最低,见图5、6。
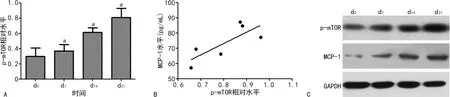
A:p-mTOR相对表达水平柱状图;B:p-mTOR水平与MCP-1水平的相关性分析;C:p-mTOR与MCP-1的蛋白电泳条带;a:P<0.05,与d0比较。

A:p-JNK相对表达水平柱状图;B:p-JNK水平与MCP-1水平的相关性分析;C:p-JNK与MCP-1的蛋白电泳条带;a:P<0.05,与d0比较。

A:0 μmol/L组;B:0.1 μmol/L组;C:1.0 μmol/L组;D:10.0 μmol/L组。

A:MCP-1水平柱状图;B:MCP-1阳性细胞数柱状图;a:P<0.05,与0 μmol/L比较。
2.5 杏仁核注射SP600125对MWT的影响
假手术组、模型组和SP600125组大鼠d0MWT比较,差异无统计学意义(P>0.05);假手术组大鼠d0、d4、d7、d14、d21MWT比较,差异无统计学意义(P>0.05);与d0相比,模型组和SP600125组d4、d7、d14、d21的MWT均降低(P<0.05)。模型组d4、d7、d14、d21的MWT均明显低于假手术组(P<0.05);经SP600125治疗后大鼠降低的MWT获改善,SP600125组d7、d14、d21的MWT均高于模型组(P<0.05),但仍低于假手术组(P<0.05),见图7。

A:0 μmol/L组;B:0.1 μmol/L组;C:1.0 μmol/L组;D:10.0 μmol/L组。
2.6 杏仁核注射雷帕霉素对MWT的影响
假手术组、模型组和雷帕霉素组大鼠d0MWT比较,差异无统计学意义(P>0.05);假手术组大鼠d0、d4、d7、d14、d21MWT比较,差异无统计学意义(P>0.05);与d0相比,模型组和雷帕霉素组d4、d7、d14、d21的MWT均降低(P<0.05)。模型组d4、d7、d14、d21的MWT均明显低于假手术组(P<0.05);经雷帕霉素治疗后大鼠降低的MWT获改善,雷帕霉素组d7、d14、d21的MWT均高于模型组(P<0.05),但仍低于假手术组(P<0.05),见图8。
3 讨 论
越来越多的证据表明,趋化因子MCP-1通过促进趋化因子受体2(CCR2)活化在慢性疼痛中发挥关键作用。MCP-1在DRG初级感觉神经元中表达,且神经损伤后表达增加[8]。到目前为止,针对产生痛觉过敏已经提出两种不同的机制:(1)MCP-1从一部分感觉神经元的中释放出来,刺激周围感觉神经元[9],在DRG中导致这些神经元过度活跃。(2)MCP-1在DRG神经元中央末端释放,诱导小胶质细胞活化,是NP发生、发展的重要机制[10]。然而,MCP-1不仅在DRG中表达,激活的星形胶质细胞体外也可产生MCP-1。研究表明,MCP-1在脱髓鞘病变、机械损伤、轴突横断及局灶性脑缺血后的星形胶质细胞中也表达[11]。此外,MCP-1在肿瘤坏死因子-α(TNF-α)刺激后的体外培养星形胶质细胞及脊髓损伤后的脊髓星形胶质细胞中表达升高[12]。本研究发现SNL后,参与调控NP的重要核团杏仁核中的MCP-1水平升高,但导致其MCP-1升高的具体机制尚不清楚。
目前有两种mTOR作为核心组分与其他不同蛋白形成的复合物,包括mTOR复合物1 (mTORC1)和mTOR复合物2(mTORC2)。在一般情况下,mTORC1由raptor、mLST8和mTOR构成,通过磷酸化下游效应分子来影响其表达,如4E-BPs和p70核糖体S6Ks。mTOR、S6K1和4E-BP1在哺乳动物神经系统中表达,尤其是在脊髓背角[13]。由于浅表背角是外周传入神经向中枢神经系统的第一个突触部位,在调节痛觉和吗啡耐受方面起着重要作用[14],故mTOR在SCI后的机械和热痛觉过敏调控中发挥重要作用。
MCP-1被证明能通过不同的机制调节疼痛,尤其对MCP-1(外周敏化)的外周机制研究较为透彻。如DRG神经元释放MCP-1可激活邻近伤害感受神经元的CCR2,通过激活瞬时受体电位香草酸亚型1(TRPV1)通道来提高这些神经元的兴奋性并抑制电压依赖性钾通道[15]。近年来MCP-1的中枢机制也成为研究热点。脊髓注射MCP-1可激活野生型脊髓小胶质细胞而不是CCR2基因敲除小鼠[16]。神经损伤引起的脊髓小胶质细胞激活可通过MCP-1抗体来缓解。此外,脊髓小胶质细胞中神经损伤诱导脊髓小胶质细胞中p38激活在CCR2基因敲除小鼠中较轻[17]。由于小胶质细胞的激活是NP产生的关键,MCP-1可能通过这种机制增强NP。
研究发现,JNK是MAPK家族成员之一,可在神经损伤后的脊髓星形胶质细胞中持续活化,而JNK抑制剂SP600125可改善脊髓损伤后的NP症状[5-6]。虽然JNK通路和MCP-1通路均参与了NP的调控,但杏仁核中JNK/MCP-1信号通路是否参与神经病理性疼痛的调控尚未见报道,本研究对其进行了初步的探讨。
本研究结果表明,SNL诱导后杏仁核的p-mTOR、p-JNK表达持续增强,且该增强效应呈时间依赖性,表明在脊髓损伤引发NP的过程中,杏仁核中的mTOR和JNK途径均被激活。进一步分析显示,d12的p-mTOR、p-JNK表达与MCP-1水平呈正相关,提示两个途径可能参与了杏仁核中MCP-1/CCR2途径的激活。杏仁核注射p-mTOR、p-JNK抑制剂发现,MCP-1水平及阳性细胞数均降低,提示两条通路的激活可能参与了杏仁核MCP-1的释放增多。此外,采用化学手段抑制两条通路活性可减轻脊髓损伤引起的NP症状,提示两条通路在NP治疗中有重要意义,可作为NP防治的靶点。
综上所述,在NP中mTOR和JNK通路激活,抑制mTOR和JNK通路可达到缓解NP大鼠疼痛及杏仁核MCP-1升高的作用,对于NP的治疗有一定意义。
