长江口地区五种腹毛类纤毛虫的形态学研究
2022-10-25张子慧祝晓璇姜佳枚
张子慧 祝晓璇 姜佳枚
(上海海洋大学水产与生命学院, 上海 201306)
纤毛虫是原生动物中最复杂和最高等的类群,在微食物网及生态系统的物质循环和能量流动中作用重大[1]。腹毛类纤毛虫广泛见于各种生境, 多底栖或是周丛生, 以细菌和藻类为食, 部分种类阶段性肉食, 可捕食其他原生生物, 是一大类最为复杂和高等的纤毛虫[2]。近20年来对于纤毛虫的研究主要集中于我国北方黄渤海水域[3], 长江口地区腹毛类纤毛虫的形态学报道几近空白状态。本文对采自长江口地区的5种纤毛虫开展了活体观察和现代银染制片, 提供了详尽的形态学描述和显微图像资料, 丰富和提高了我国腹毛类纤毛虫物种多样性的认识。
1 材料与方法
冠突伪尾柱虫(Pseudourostyla cristata)于2020年9月23日采自上海市崇明县横沙乡泥滩(温度20.1℃、pH 7.35、盐度0.25‰、培养盐度0.2‰),仁川巴库虫(Bakuella incheonensis)和红色伪角毛虫(Pseudokeronopsis rubra)于2020年10月23日采自江苏省启东市寅阳镇泥滩(温度22.6℃、pH 7.93、盐度10‰、培养盐度6‰), 三种纤毛虫均取泥滩表层约10 cm处孔隙水和底质。黄色伪角毛虫(Pseudokeronopsis flava)于2020年9月23日采自上海市崇明县河道(温度19.2℃、pH 7.4、盐度0.4‰、培养盐度0.2‰), 以采水器取10 L水通过13#和25#的浮游生物网后装入采样瓶。希斯多利织毛虫(Histriculus histrio)于2021年3月8日以载玻片法[1]采自上海海洋大学校内河道水深约1 m处, 期间采样点水温4.3—9.2℃、pH 8.3—8.78、盐度0.5‰。底质样品取部分用双层纱布包裹, 河道样品直接倾倒或取载玻片放入培养皿后, 加原位水和米粒置于室温培养。使用Olympus BX53显微镜放大100—1000倍进行活体观察、记录和拍照, 蛋白银法染色。分离、培养、活体观察和染色制片等研究方法及名词术语主要参照Corliss[4]、Song等[5]。分类系统参照Lynn[6]。
2 结果与讨论
2.1 仁川巴库虫Bakuella (Bakuella) incheonensis Jo, et al., 2015(图版I-1—7)
尾柱目Urostylida Jankowsk, 1979
巴库科Bakuelidae Jankowski, 1979
形态学描述: 活体大小(88—144) μm×(30—65) μm(图版I-1), 长宽比为3: 1; 虫体长椭圆形, 两端钝圆,后部略窄。虫体柔软可弯曲。皮膜坚实, 具有两种皮层颗粒: 一种近似椭球状(约0.4 μm×0.7 μm), 黄棕色至黄绿色(图版I-6, 长箭头), 腹面沿棘毛基部分布,背面分布呈不规则纵列(图版I-4, 5, 短箭头), 使细胞呈淡淡的半透明黄绿色或灰色; 另一种圆球状(直径0.8—1.2 μm), 无色(图版I-6, 短箭头), 数目众多, 在虫体背面和腹面密集排布。虫体腹面沿着中腹棘毛复合体具有纵沟。胞质无色, 通常含有许多油球(直径3—5 μm)和绿色食物泡(直径9—13 μm)。温度降低(约5℃)时虫体形成深灰色圆球形包囊(直径29—41 μm),表面分布有黄绿色椭球状皮层颗粒(图版I—2, 3)。伸缩泡一个, 位于虫体左缘1/3处。58—81枚椭圆形大核。通常于底质上缓慢直向爬行, 遇刺激时则迅速后退、转向, 有时于水体中绕体纵轴旋转游动。
口围带为体长的1/4—1/3, 由30—38片小膜组成(图版I-7)。3根额棘毛, 活体下长16—20 μm; 5或6根额前棘毛。体纤毛较为细弱, 活体下8—10 μm。口侧膜长度约是口内膜长度的2/3, 二者在接近口侧膜末端处相交。1根口棘毛靠近口侧膜, 1根拟口棘毛。中腹棘毛延伸至虫体3/5—4/5处, 中腹棘毛复合体由12—17对中腹棘毛对和1列中腹棘毛列共同组成; 左、右缘棘毛各1列, 分别含32—42根、27—46根棘毛, 左缘棘毛延伸至虫体末端; 4—6根横棘毛, 位于虫体近末端; 1根横前棘毛(4/15个体缺失)。3列背触毛纵贯虫体, 右缘棘毛延伸至虫体背部, 其前端具有两对毛基体。
比较与讨论: 巴库虫属由Agmaliev等[7]于1976年建立, 海洋巴库虫(Bakuella marina)为该属模式种。1992年Alekperov[8]为其中具两根额棘毛的种建立伪巴库虫属(Pseudobakuella)。Berger[9]根据额棘毛的数量将巴库虫属分为两个亚属: 巴库虫亚属Bakuella(Bakuella)和伪巴库虫亚属Bakuella(Pseudobakuella)。前者具有两根以上额棘毛, 后者具有两根额棘毛。巴库虫属其余特征为口围带连续, 2根或多根额前棘毛, 1或多根口棘毛; 中腹棘毛复合体由中腹棘毛对和一列中腹棘毛共同组成, 左右各一列缘棘毛, 具有横棘毛, 无尾棘毛[10]。
此种为国内新记录种, 原始种群由Jo等[11]在2015年报道自韩国仁川阿姆多海岸公园的微咸水(盐度4.1‰)内。与原始描述相比本种群个体较大(88—144 μmvs.70—105 μm), 口围带小膜数较多(30—38vs.21—25),额前棘毛较多(5或6vs.3或4)。该种与Chen等[12]于2013年报道的亚热带巴库虫(Bakuella subtropica)最为相似, 主要区别为仁川巴库虫的伸缩泡位置(近虫体左缘距体前1/3处vs.近虫体左缘距体前1/2处), 额前棘毛较少(5或6vs.4—12), 中腹棘毛对较少(12—17vs.9—23)。
2.2 希斯多利织毛虫Histriculus histrio Corliss,1960(图版I-8—11)
散毛目 Sporadotrichida Faure-Fremiet, 1961
尖毛科 Oxytrichidae Ehrenberg, 1838
形态学描述: 活体大小(122—149) μm ×(47—61) μm, 长宽比2∶1—3∶1, 宽厚比约4∶1(图I-8)。虫体近似长椭圆形, 前端钝圆, 后端略窄, 右缘近平直,左缘略外凸, 左右边缘的中部近乎平行。腹面扁平,背中部微微隆起。皮膜坚硬。虫体坚实, 不可伸缩或弯折。无皮层颗粒。单一伸缩泡位于虫体中间近左缘(图版I-9, 长箭头), 直径约13 μm, 收缩周期约为6s。细胞质无色, 内含有许多球状颗粒、油球、不规则晶体、杆状藻类, 致使虫体在低倍镜下呈半透明灰色。虫体表面有发达的表膜泡状结构(图版I-10, 箭头), 左缘密集, 右缘稀疏分布。两枚卵球形大核, 1—3枚小核。通常在底质上缓慢爬行或停滞, 遇到刺激时急促爬行。
口围带延伸至虫体约1/2处, 由36—49片小膜组成, 口纤毛最长可达16 μm。3根十分粗壮的额棘毛;4根额腹棘毛呈“v”字形排列, 其中3根呈直线列, 几乎与波动膜平行(图版I-11)。口侧膜与口内膜长度几乎相等, 略弯曲, 二者相交于中部, 呈尖毛虫属(Oxytricha)模式。一根粗壮的口棘毛, 位于波动膜上方并靠近波动膜前端; 3或4根口后腹棘毛位于胞口后方; 2根横前腹棘毛位于横棘毛前方; 左、右缘棘毛各1列, 末端分离不明显, 分别具21—29、28—39根棘毛; 5根横棘毛, 明显前移, 呈“J”字形紧密排列并伸出体外, 活体下长约20 μm。6列纵贯虫体的背触毛。
比较与讨论: Corliss[13]于1960年建立织毛虫属,该属虫体僵硬, 波动膜为尖毛虫属模式, 左右缘棘毛各一列, 无尾棘毛, 6列背触毛。
此种为国内新记录种, 在德国、匈牙利均曾多次发现[14—18], 国内外种群纤毛图式高度一致。 虽然中国种群大小(122—149 μm)和部分国外种群存在差异, 但仍在诸多种群的整合数据范围内(86—215 μm)。该种与扁平织毛虫(Histriculuscomplanatus)在活体特征及纤毛图式上近乎一致, 最大的差别在于后者体型较小(149 μmvs.80 μm)、活体状态下呈明显椭圆形以及横棘毛没有明显前移[19]。
2.3 红色伪角毛虫Pseudokeronopsis rubra(Ehrenberg,1836)Borror & Wicklow, 1983(图版I-12—15)
尾柱目Urostylida Jankowsk, 1979
伪角毛科Pseudokeronopsidae Borror & Wick-low, 1983
形态学描述: 活体大小(92—337) μm ×(35—79) μm,长宽比3∶1—4∶1, 宽厚比约3∶2, 个体间大小、体型差异较大(图版I-14)。虫体通常为细长的椭圆形,不可伸缩, 在低倍镜下观察, 虫体呈暗红色。皮膜厚实, 柔软易弯曲。两种皮层颗粒(图版I-13): 一种圆球状(直径0.5—1.5 μm), 红棕色, 腹面沿棘毛基部排列, 背面每6—8个一组, 绕背触毛呈花团状, 另有若干散布于皮膜下; 另一种红细胞状(直径约1 μm),无色, 于背腹面均匀密集排布。胞质无色, 具若干球形油滴, 较大虫体的胞质中普遍含有1—4个橙红色食物泡。单一伸缩泡(图版I-12, 长箭头)位于虫体左缘下方3/4处。大核约150枚, 散乱分布在胞质中。运动方式为附底质缓慢爬行。
口围带延伸至虫体4/1—1/3处, 由45—67片小膜组成, 口纤毛最长可达17 μm(图版I-15)。额棘毛呈双冠状排布, 前列具有7—9根棘毛, 后列具有6—9根棘毛; 2根额前棘毛。口侧膜较短, 约为口内膜长度的1/2; 二者平直。通常1根口棘毛, 位于口侧膜中部附近; 中腹棘毛复合体由23—46对锯齿状排布的棘毛组成, 起始于口侧膜中部; 左、右缘棘毛各1列, 分别具43—81、40—79根棘毛; 6—11根横棘毛。横棘毛长约12 μm, 其他棘毛长约8 μm。背面4列背触毛, 纵贯虫体。
比较与讨论: 该种最早由Ehrenberg[20]于1836年发现, 命名为Oxytricha rubra。1983年Borror等[21]以该种作为模式种建立伪角毛虫属, 并将其更名为P.rubra。该属种类普遍具有颜色鲜艳的皮层颗粒,种间伸缩泡位置差别很大, 是重要的定种依据。口围带连续, 额棘毛双冠状排布, 不少于2根额前棘毛,口棘毛存在; 中腹棘毛复合体由中腹棘毛对组成,左右缘棘毛各一列, 具有横棘毛; 不少于3列背触毛, 无尾棘毛; 许多大核。本种群与Hu等[22]所报道的青岛种群相比, 个体大小波动范围较大(92—337 μmvs.160—200 μm), 可能因实验室培养时间过长, 个体大小的分化严重; 与Li等[23]于2017年报道的宋氏伪角毛虫(P.songi)相比, 二者皮层颗粒具有相同的颜色、大小和排布方式, 但二者的伸缩泡位置不同(虫体左缘vs.虫体右缘), 横棘毛数目(6—11vs.2—4)和背触毛列数(4vs.6)较多。
2.4 黄色伪角毛虫Pseudokeronopsis flava (Cohn,1866) Wirnsberger, Larsen & Uhlig, 1987(图版Ⅱ-1—10)
尾柱目Urostylida Jankowsk, 1979
伪角毛科Pseudokeronopsidae Borror & Wicklow, 1983
形态学描述: 活体大小(175—215) μm×(25—40) μm, 长宽比5∶1—6∶1, 宽厚比约3∶2(图版Ⅱ-1,3—5)。虫体通常为细长的椭圆形至长带状, 后端略窄; 背腹不平, 略微隆起; 柔软、易弯曲, 不可伸缩。低倍镜下虫体呈黄棕色。皮膜单薄, 两种皮层颗粒: 一种圆球状(直径0.4—10.9 μm), 橘黄色, 腹面沿棘毛基部呈短列状排布成一列(图版Ⅱ-6、7长箭头), 共3列, 背面绕背触毛呈花团状排布; 另一种红细胞状(直径0.9—1.8 μm), 无色, 于背腹面均匀密集排布(图版Ⅱ-6、7短箭头)。胞质无色, 具若干大小不一的食物泡和油球(图版Ⅱ-2)。单一伸缩泡位于虫体左缘体后3/4—4/5处, 收缩周期约30s。椭球形大核, 约100枚, 散布在胞质中。运动方式为附底质缓慢爬行, 后退时身体微微收缩并扭曲以改变运动方向。
口围带延伸至虫体1/5—1/4处, 由45—62片小膜组成, 口纤毛最长可达18 μm(图版Ⅱ-10)。9—17根额棘毛双冠状排布(图版Ⅱ-8), 前、后列各含4—8、5—9根棘毛, 长约9 μm; 2或3根额前棘毛(图版Ⅱ-9, 圈)。口侧膜与口内膜几乎等长, 相交于1/4处。口棘毛1根, 靠近波动膜前1/4处。中腹棘毛复合体由23—36中腹棘毛对组成, 延伸至体后约4/5处; 左、右缘棘毛各1列, 分别具43—61、40—59根棘毛; 3根横棘毛, 长约15 μm, 活体下明显伸出虫体后缘。3列背触毛, 纵贯虫体。
比较与讨论: 此种群与Song等[24]报道的湛江种群相比, 额棘毛数目较多(9—17vs.8—12), 其余特征基本相符合。该种与Song等[25]2007年报道的浅黄伪角毛虫(P.flavicans)最为相似, 二者体色、体型近乎相同且皮层颗粒颜色及排布方式基本一致,主要区别为伸缩泡位置(虫体体后3/4—4/5处vs.虫体体前1/3—2/5处)以及背触毛列数(3vs.4或5)。
2.5 冠突伪尾柱虫 Pseudourostyla cristata (Jerka-Dziadosz, 1964) Borror, 1972(图版Ⅱ-11—17)
尾柱目Urostylida Jankowsk, 1979
伪尾柱科 Pseudourostylidae Jankowski, 1979
形态学描述: 活体大小(166—340) μm ×(32—83) μm, 长宽比为4∶1—5∶1, 宽厚比约为2∶1, 个体间体型差异较大(图版Ⅱ-11, 12)。虫体长椭圆形, 两端钝圆。虫体灵活, 可收缩、易弯曲; 皮膜柔软, 存在皮层颗粒(图版Ⅱ-13, 长箭头), 圆球状, 无色, 直径(0.4—1.6 μm), 在背腹面散乱排布, 前后端较密集。未激发的射出体(图版Ⅱ-13, 短箭头)密集排布于皮膜下方。胞质无色, 内含众多食物泡、油球等内质颗粒, 导致虫体呈灰黑色。单一伸缩泡(图版Ⅱ-12, 短箭头)位于虫体左缘前方约2/5处, 直径约24 μm。椭球状大核, 39—63个, 散乱分布在胞质中。虫体通常在底质上爬行, 前进时较为缓慢, 遇到刺激时迅速后退、调转方向。
口围带延伸至虫体约2/5处, 由83—94片小膜组成(图版Ⅱ-17)。额棘毛双冠状排布(图版Ⅱ-15), 前列具有11—13根棘毛, 后列具有9—11根棘毛; 2或3根额前棘毛。口侧膜较口内膜短, 二者相交于口侧膜后2/3处。1根口棘毛, 靠近口侧膜1/2处;中腹棘毛复合体由16—25中腹棘毛对组成; 4—5列右缘棘毛; 4—6列左缘棘毛; 7—9根横棘毛呈“J”字形排列(图版Ⅱ-16), 位于距体后缘1/6处。8列触毛,纵贯虫体背部(图版Ⅱ-14)。

图版 Ⅱ 黄色伪角毛虫(1—10)和冠突伪尾柱虫(11—17)的活体(1—7, 11—14)及蛋白银染色后(8—10, 15—17)的显微照片Plate Ⅱ Morphology and infraciliature of Pseudokeronopsis flava (1—10) and Pseudourostyla cristata (11—17) in vivo (1—7, 11—14)and after protargol staining (8—10, 15—17)
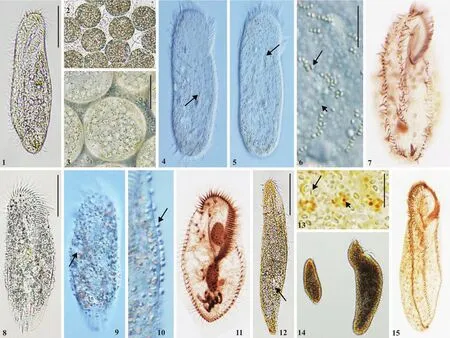
图版 Ⅰ 仁川巴库虫(1—7)、希斯多利织毛虫(8—11)和红色伪角毛虫(12—15)的活体(1—6, 8—10, 12—14)及蛋白银染色后(7, 11,15)的显微照片Plate Ⅰ Morphology and infraciliature of Bakuella incheonensis (1—7), Histriculus histrio (8—11) and Pseudokeronopsis rubra (12—15)in vivo (1—6, 8—10, 12—14) and after protargol staining (7, 11, 15)
比较与讨论: Borror[26]于1972年建立伪尾柱虫属, 指定冠突伪尾柱虫为模式种。该属的基本特征包括: 口围带连续, 额棘毛呈双冠状排布, 具额前棘毛、口棘毛, 中腹棘毛复合体仅有中腹棘毛对, 具有多列左右缘棘毛, 存在横棘毛、横前棘毛, 无尾棘毛。
与1996年Oberschmidleitner等[27]报道的奥地利种群相比, 该种群体型较小(166—340 μmvs.220—450 μm)、大核数较多(39—63vs.27—55)、小核数波动范围较大(2—8vs.3—7), 其余特征吻合良好。与Chen等[28]报道的日本种群相比, 本长江口种群大核较多(39—63vs.15—36), 活体形态、伸缩泡位置、纤毛的排布模式及数量等特征吻合良好。该种与贵州伪尾柱虫(P.guizhouensis)[29]具有相似的体型、体色和伸缩泡位置等, 但后者右缘棘毛列较少(4或5vs.2)。
