结核分枝杆菌Mce4蛋白家族研究进展
2022-10-14刘原园李璐吴托雅鲁洁
刘原园 李璐 吴托雅 鲁洁
2020年全球约有结核分枝杆菌(Mycobacteriumtuberculosis,MTB)感染者990万例,我国结核病患者数占全球总数的8.5%,位居世界第二[1]。结核病治愈率较低,死亡率较高,对公众健康构成巨大威胁[2-3]。MTB是一种胞内病原菌,在代谢和复制活跃的阶段侵入宿主巨噬细胞后可进入长期休眠,以此来逃避宿主的免疫防御。一旦宿主的免疫压力减弱,MTB即乘机复苏,恢复到代谢和复制活跃的阶段[4]。MTB中的哺乳动物细胞入侵(mammalian cell entry,Mce)蛋白家族由mce操纵子所编码,在侵袭宿主巨噬细胞和促进MTB存活过程中的作用逐渐被重视。其中Mce4系统具有调控宿主免疫和转运胆固醇的作用[5-6],笔者就mce4操纵子及其编码蛋白功能的研究进展进行综述。
一、mce4操纵子组成及功能
1.mce操纵子组成:对结核分枝杆菌H37Rv基因组的完整序列进行分析,发现其有4个mce操纵子,分别为mce1~mce4(图1)[7]。每个操纵子主要由2个yrbE基因、6个mce基因组成[8]。此外,还包括mce1操纵子下游的4个基因(mas1A~1D)及mce3、mce4操纵子下游的2个基因(分别为mas3A和mas3B、mas4A和mas4B)。
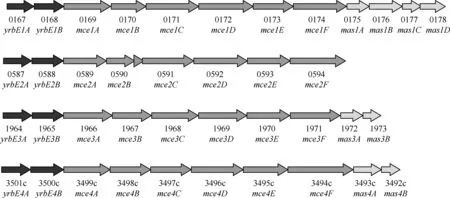
图1 结核分枝杆菌H37Rv mce(1~4)操纵子示意图
2.mce4操纵子及编码蛋白:mce4操纵子具有8个基因[8],其中yrbE4A(Rv3501c)和yrbE4B(Rv3500c)既是mce4操纵子的一部分,也是YrbE家族的成员,编码YrbE4A和YrbE4B蛋白,注释为“假想保守整合膜蛋白”,预测可能是ATP结合盒(ATP-binding cassette, ABC)转运体,与脂代谢相关[9-10]。而mce4A~4F(Rv3499c~Rv3494c)分别编码Mce4A~4F 蛋白,都被预测与入侵宿主细胞或脂代谢相关[9-10]。操纵子多态性程度分析中发现,mce4操纵子的多态性明显高于mce1操纵子,yrbE4A和Rv3495c在mce4操纵子中多态性最高[11]。
3.mce4操纵子的表达调节:细胞内环境使结核分枝杆菌暴露于多种应激因素,包括缺氧、营养限制、氧化和酸性条件[12-13]。为了适应不利条件并持久存活,结核分枝杆菌对其基因表达进行调整,以便在恶劣的环境中生存[14]。研究发现,在存在表面应激和细胞内氧气缺乏的情况下,耻垢分枝杆菌mce4操纵子启动子活性被上调了1.5倍[15]。Rathor等[14-15]研究也证实,在表面应力、缺氧以及在胆固醇的存在时,结核分枝杆菌mce4操纵子的启动子被诱导,且其所有基因都比在有氧培养条件下表达升高。此外,mce4操纵子启动子区域显示出对肽基脯氨酰顺反异构酶A(peptidyl-prolyl cis-trans isomerase A,PPIase A)的亲和力[15],而PPIase A是结核分枝杆菌在胆固醇作为唯一碳源的情况下生存所必需的[16]。以上研究证明了缺氧和胆固醇是影响结核分枝杆菌mce4操纵子表达的重要因素。
4.Mce4系统在胆固醇转运中的作用:Mce4系统被认为是重要的胆固醇转运系统。Casali和Riley[7]认为放线菌是mce位点编码的一种新型的ABC转运蛋白。随后Mohn等[17]在约氏红球菌(rhodococcusjostii,RHA1)转录分析中发现,在胆固醇和丙酮酸的作用下,mce4操纵子的表达上调了4.0倍;且生物信息学分析表明,测序基因组中的所有mce4位点均与类固醇代谢基因连锁,而胆固醇是类固醇的一种。Van der Geize等[18]发现,mce4和yrbE突变体在有胆固醇培养基中的生长能力都严重受损,证明了Mce4和yrbEA及yrbEB参与了约氏红球菌中胆固醇的摄取,也表明了约氏红球菌的mce4基因座可编码类固醇摄取系统。Pandey和Sassetti[5]也证明,mce4可编码一个胆固醇输入系统,使结核分枝杆菌能够摄取和利用宿主膜的胆固醇以获得碳和能量。2012年,Klepp等[19]发现了mce4操纵子的缺失也会损害耻垢分枝杆菌的胆固醇摄取,这一发现更加明确了Mce4蛋白也参与了耻垢分枝杆菌对胆固醇的转运。2017年,García-Fernández等[20]研究发现了携带mce4操纵子上任一基因突变体的耻垢分枝杆菌在含胆固醇的培养基中无生长,且缺失的mce4基因均不能被耻垢分枝杆菌其他mce操纵子中的同源基因所取代,推测Mce4系统可能是耻垢分枝杆菌中唯一的胆固醇主动转运系统。2020年,有研究通过依折麦布(ezetimibe)药物来降低斑马鱼体内游离胆固醇和中性脂,继而有效抑制了海洋分枝杆菌的生长,而这一过程依赖于mce4操纵子[21],可见该操纵子是脂类利用的关键决定因素。同年Alonso等[22]发现鸟分枝杆菌副结核亚种中mce4操纵子的缺失诱导了甘油三酯的积累,也导致了菌落形态的改变,提示mce4操纵子可能与脂质利用以及细胞膜的结构相关。
二、Mce4蛋白功能研究进展
Mce4蛋白家族有8个蛋白,分别是YrbE4A、YrbE4B、Mce4A~4F,都具有Mce家族特征性的Mce结构域。此外,Mce4A和Mce4D还具有胆固醇摄取转运蛋白结构域[23],提示与胆固醇转运功能相关。已有的相关研究报道多集中在Mce4A、Mce4C~4F这5个蛋白,现对其功能的最新研究进展进行综述。
(一) Mce4A蛋白
MTB H37Rv菌株Mce4A蛋白由400个氨基酸组成[24]。蛋白稳定性研究证明十二烷基硫酸钠与Mce4A有较强的结合力,具有稳定Mce4A的作用[25]。生物信息学预测表明,Mce4A蛋白不仅是膜蛋白,而且含有18个B细胞抗原表位和23个T细胞抗原表位[26],属于免疫优势抗原,推测容易被免疫系统识别而产生相应的免疫应答。相互作用蛋白预测显示Mce4A蛋白主要与 Mce4B、Mce4E、YrbE4B、Mce4A、Mce4D 蛋白协同发挥作用[26],作用机制尚不清楚。配体结合研究表明25-羟基胆固醇作为配体与Mce4A有较强的结合亲和力,有助于设计Mce4A的竞争性抑制剂[27]。此外, IP-MS确定耻垢分枝杆菌Mce4A免疫沉淀富集了Mce4B~4F, 且通过免疫共沉淀证实Mce4A和Mce4E存在相互作用,提示Mce4A~4F可能形成高度稳定的复合体或作为多蛋白Mce4转运复合体发挥作用[28]。
1.Mce4A蛋白的侵袭力和毒力:有研究表明,结核分枝杆菌Mce4A蛋白定位于菌体细胞壁部分,在非致病性大肠杆菌中表达的Mce4A蛋白使其能够进入并存活于HeLa细胞和巨噬细胞内[29]。这验证了Mce4A蛋白在细胞侵袭以及在巨噬细胞中存活的作用与Mce1A相似。Sinha等[30]发现Mce4A蛋白过表达的结核分枝杆菌感染后,小鼠体内的细菌载量升高近两倍,并具有早期疾病状况和肺部广泛的组织破坏和淋巴细胞浸润等病理改变。也有研究发现Mce4A在H37Rv中过表达可导致与结核分枝杆菌致病性相关的炎性脂邻苯二酚二霉菌酸酯(phthiocerol dimycocerates,PDIM)和硫脂(sulfolipid,SL1)增加[31]。这些都提示Mce4A蛋白对结核分枝杆菌的致病能力有一定影响。
2.Mce4A蛋白对宿主免疫的调控:有实验证实,重组牛分枝杆菌Mce4A蛋白可上调肺泡巨噬细胞肿瘤坏死因子α(tumour necrosis factor-α,TNF-α)、诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)和白细胞介素-6(interleukin-6,IL-6)的表达[32]。He等[33]研究也发现类似结果,表达感染的重组牛分枝杆菌Mce4A蛋白显著上调了巨噬细胞IL-1β、iNOS和TNF-a的mRNA水平。这提示Mce4A蛋白可能通过诱导巨噬细胞产生促炎细胞因子从而影响机体对病原体的免疫反应。其中iNOS能生成高反应性自由基一氧化氮(NO),NO的产生会抑制呼吸并诱导结核分枝杆菌的休眠[34]。推测Mce4A蛋白可能通过NO使结核分枝杆菌在宿主体内持久存活。最近的研究表明,结核分枝杆菌Mce4A蛋白可诱导巨噬细胞分泌TNF-α或γ-干扰素(interferon-γ,IFN-γ)等促炎细胞因子[6]。其中TNF-α能正向调节半胱氨酸蛋白酶的激活和NO的产生,从而诱导DNA损伤导致巨噬细胞发生凋亡[6,35]。因此,一方面,NO的呼吸抑制诱导结核分枝杆菌休眠,有利于结核分枝杆菌在宿主体内的持久存活;另一方面,Mce4A蛋白以TNF-α依赖的方式诱导巨噬细胞凋亡。这表明Mce4A蛋白诱导巨噬细胞产生促炎细胞因子,与结核分枝杆菌保护性免疫反应的发展有关[35-36]。最近有研究设计了一种能针对Mce4A的反义封闭核酸间隔体(Mce4A-ASO),能显著降低结核分枝杆菌感染的成骨细胞的细菌载量以及IL-6的分泌,提示可作为脊柱结核治疗的研发方向[37]。
3.Mce4A蛋白介导结核分枝杆菌摄取利用胆固醇:等温滴定热法首次证实Mce4A蛋白与胆固醇在体外结合,为Mce4A在结核分枝杆菌发病机理中摄取利用胆固醇的可能作用提供了直接证据[38]。在胆固醇存在下,过表达Mce4A的结核分枝杆菌菌株的生长速度是其他菌株的2倍,证明Mce4A能更有效地利用胆固醇[30]。电镜观察到过表达Mce4A的结核分枝杆菌细胞壁增厚,其原因可能是延长的游离脂质如PDIM转移积累到细胞壁导致的,而PDIM的合成途径是由Mce4A蛋白中类似甲基接受趋化蛋白(methyla-ccepting chemotaxis protein,MCP)的特定结构域调控[30-31],提示Mce4A是结核分枝杆菌细胞壁PDIM积累所需代谢途径的调节因子。而PDIM已经被证实是致病性分枝杆菌具有的细胞壁毒力相关脂质[39-40],这是Mce4A蛋白参与结核分枝杆菌致病过程的可能证据。以上证据表明,Mce4A在介导结核分枝杆菌摄取胆固醇的同时,在增强胞内存活期间的细胞壁强度和毒力中也具有令人信服的作用。
(二)Mce4C蛋白
结核分枝杆菌的Mce4C蛋白由357个氨基酸组成[24],但相关研究很少。有研究预测了Mce4C蛋白MHC Ⅰ类表位—SLAGNSAKV,认为是抗结核疫苗研发的候选表位[41]。
(三)Mce4D蛋白
结核分枝杆菌的Mce4D蛋白由451个氨基酸组成[24]。在牛分枝杆菌弱毒株和超强毒株的差异转录组图谱分析中,证实mce4D在强毒株中表达上调[42]。Mce4D蛋白功能预测也是与脂质代谢相关,但还有待研究。此外,也预测了Mce4D蛋白的MHCⅡ类表位—PNLVAARFIQLTPVY被认为是另一个抗结核疫苗研发的候选表位[41]。
(四)Mce4E蛋白
结核分枝杆菌的Mce4E蛋白又称为LprN蛋白,由384个氨基酸组成[24]。生物信息学预测Mce4E蛋白为一种脂蛋白[9-10]。脂蛋白具有高效的免疫原性,影响先天和获得性免疫反应[43]。有报道发现,牛分枝杆菌Mce4E蛋白能够抑制牛肺泡巨噬细胞的活性,同时促使炎性细胞因子TNF-α和IL-6的mRNA表达上调,抑制iNOs mRNA的表达[44],由此可推测Mce4E蛋白通过诱导巨噬细胞产生促炎细胞因子从而影响机体对病原体的免疫反应。实验证明,在大肠杆菌中表达的重组Mce4E蛋白在小鼠体内引发了细胞免疫应答,表现为明显提高T细胞增殖和增加TNF-a和IFN-γ细胞因子的产生,从而诱导小鼠Th-1型保护性免疫应答;同时,组织病理学检查显示组织破坏增加[45],表明Mce4E蛋白具有增强细菌毒力的免疫原性潜力。由于mce4E基因是在结核分枝杆菌感染后期表达的操纵子的一部分[15],因此认为Mce4E蛋白可能通过调节其免疫应答而在动物的疾病维持中发挥作用。还有研究提示,Mce4E蛋白可能比Mce4A更能促进细菌在巨噬细胞中的持久性,从而导致其在被巨噬细胞吞噬后能够更好地存活[33],但更有力的证据还有待进一步研究。此外,Mce4E蛋白也被证明是耻垢分枝杆菌吸收胆固醇所必需的[28]。
(五)Mce4F蛋白
结核分枝杆菌的Mce4F蛋白由564个氨基酸组成,被预测在细菌脂质分解代谢中发挥着重要作用[18]。Mce4F蛋白被认为是可能的外膜蛋白之一,它分泌在菌体外膜上,具有与分枝杆菌致病相关的重要功能[46]。免疫电镜证实了其存在于分枝杆菌表面,并用125I 结合试验鉴定了Mce4F蛋白的高活性结合肽(high-activity binding peptides,HABP),结果发现,所有的HABP均有抑制分枝杆菌进入肺泡上皮细胞A549和单核细胞来源的巨噬细胞U937的作用[47]。以上结果提示,Mce4F蛋白可能在分枝杆菌的毒力和感染适应中发挥了负性作用,其抑制侵袭宿主细胞的机制及意义有待进一步研究。此外,Mce4F蛋白也被证明是耻垢分枝杆菌吸收胆固醇所必需的[28]。
三、展望
在MTB感染宿主过程中,越来越多的细菌蛋白被报道能够调控宿主细胞信号或利用宿主促进自身存活,例如MTB的Rv1579c(probable PhiRv1 phage protein,EST-12)能够通过活化C激酶1受体诱导NLRP3炎症体活化,从而促进巨噬细胞凋亡[48];MTB的Rv0859能够靶向宿主脂肪酸代谢从而抑制炎症,促进细菌存活[49]。目前,研究认为MTB的Mce4蛋白家族也主要在参与调控宿主免疫、入侵宿主细胞、介导胆固醇摄取中起着重要作用。尤其是摄取胆固醇并从中获得碳和能量对于结核分枝杆菌在宿主巨噬细胞内长期存活是必不可少的。
结核分枝杆菌维持慢性感染的能力与其从宿主获取胆固醇的能力密切相关,但仍缺乏mce4操纵子上的mce4B和mce4C等相关的研究。同时,mce4操纵子及编码蛋白在胆固醇摄取以及发病机制方面是否存在更复杂或多重的功能仍未可知,对Mce4蛋白功能更整体全面的理解还有待进一步研究。目前研究认为,mce4操纵子编码一种类似的ABC转运蛋白的胆固醇摄取系统,但还不清楚为什么Mce系统比经典的ABC转运蛋白需要这么多的蛋白质。有人认为这是由于Mce4蛋白形成了一个复合体,在经典ABC摄取转运蛋白中发挥底物结合蛋白的作用[20]。另外,Mce4F对结核分枝杆菌进入细胞的抑制作用也需要进一步研究,这有望成为治疗结核感染的新方向。
以往对于Mce系统的研究局限于单一蛋白的功能研究,而不是以复合物形式表达的整个操纵子。若是对Mce4蛋白系统整体的生物学功能及其系统中各个蛋白的相互作用机制进行研究,则可能为理解Mce4蛋白在结核分枝杆菌发病机制中的作用提供新的视角,也可能为结核病的治疗提供一个新的靶点。
利益冲突所有作者均声明不存在利益冲突
作者贡献刘原园:文章撰写;李璐和吴托雅:文献检索和筛选,文章校对;鲁洁:文章校对及修改,经费支持
