PI3K/Akt/GSK-3β/CREB通路参与调控高原适应人群运动氧化应激的机制研究
2022-10-08孔海军李新龙王凤华聂武生
孔海军,李新龙,王凤华,聂武生
氧化应激是一种被称为“氧悖论”的生理现象,与各种病理生理反应有关,其生成过程与活性氧(Reactive oxygen species,ROS)水平升高或抗氧化能力下降诱导的细胞抗氧化防御系统衰减效应密切相关。研究表明,高原缺氧状态下氧利用效率下降导致细胞线粒体呼吸减弱,ROS生成系统随着酶和非酶抗氧化系统的抑制而被激活,诱导线粒体电子传递链发生ROS堆积。正常范围内的氧化应激对于机体代谢调控、细胞信号转导、组织修复以及运动适应性反应发挥重要作用,但超限度的氧化应激可能诱导组织细胞损伤。

cAMP反应元件结合蛋白(cAMP-response element binding protein,CREB)是细胞增殖和凋亡所需的一种基本核转录因子,因为它参与调节影响细胞存活相关的一系列基因表达,包括B细胞淋巴瘤2(B-cell lymphoma-2,Bcl-2)和B细胞超大淋巴瘤-XL(Bcl-XL)。CREB在多数器官中普遍表达,是细胞增殖、生长、存活和分化等多种生物学过程的必需活性物,参与调节记忆、学习、突触传递及脑神经元存活、分化和轴突生长。磷酸肌醇3-激酶(PI3K)/Akt信号通路已被证实在介导细胞凋亡及能量代谢适应进程中发挥重要作用,磷酸化AKT触发下游靶点的磷酸化,导致糖原合成酶激酶-3(GSK-3)和促凋亡因子Bcl-2家族成员Bad失活,进一步激活cAMP反应元件结合蛋白(CREB)、核因子kB(NF-kB)、p70核糖体S6激酶(p-p70S6K)和内皮型一氧化氮合酶(NOS3)。以往的研究证实,PI3K/Akt/GSK-3/CREB信号通路可能在运动氧化应激进程中发挥重要的调控作用。但国内外鲜见高原低氧环境中PI3K/Akt/GSK-3/CREB通路调控高原适应人群运动氧化应激的机制研究。基于此,通过研究不同高原适应时期人群运动氧化应激特征,并讨论不同高原适应时期PI3K/Akt/GSK-3/CREB信号通路及其磷酸化水平对运动氧化应激的调节作用,以期为高原适应及高原训练提供实验依据。
1 测试对象与方法
1.1 测试对象
研究随机抽取世居3 000 m以上高原的20~40岁健康男性及移居高原(移居3 000 m以上高原超1年)20~40岁健康男性为研究对象。受试者纳入标准:①身体形态、体格发育正常;②无肢体残疾;③近一年无手术史;④无酒精、药物依赖及吸烟史;⑤可正常完成预定Conconi运动试验。排除或剔除标准:①患有严重的心、肺、肝、肾等系统疾病;②近3个月有急进或急下高原史;③患慢性病及长期服药史。最终,本次研究纳入202名受试者,受试者的年龄(Y)及BMI(kg/m)等人口学特征指标均不存在显著差异(>0.05),见表1。所有研究对象在测试前均知情同意,研究经喀什大学生物及医学伦理委员会审核批准。
研究选择新疆喀什地区塔什库尔干塔吉克自治县班迪尔乡为测试点,该地区属高原荒漠气候,平均海拔3 200 m,年均气温3.3℃,年均光照时间2 831 h,大气压及氧分压分别为70.5 kpa(530 mmHg)及14.8 kpa(111 mmHg)。按照受试者高原习服时相特征区分为初步高原适应即高原适应1组(High Altitude Adaptation 1 Group,HA)、基本高原适应即高原适应2组(High Altitude Adaptation 2 Group,HA)、完全高原适应即高原适应3组(High Altitude Adaptation 3 Group,HA)及世居高原组(Control Group,Con),受试者上高原年限见表1。

表1 受试者基本信息表
1.2 测试方法
1.2.1 Conconi运动试验
Conconi运用改良后逐级递增负荷运动试验。操作流程包括准备活动(10 min,慢跑+拉伸训练)、递增负荷跑(6级,3 min/级,6 m·s为起始,组间间歇1 min),具体测试流程见图1。

图1 Conconi运动试验流程图
1.2.2 氧化应激、炎症反应指标测试
所有受试者均于Conconi运动后即刻卧位抽取静脉血5 ml置于肝素抗凝管中,离心取血清置于EDTA管中,冻存于低温环境中备用。采用ELISA法测定谷胱甘肽过氧化物酶(Glutathione peroxidase,GSH-Px)、血清一氧化氮(Nitric Oxide,NO)、活性氧(Reactive oxygen species,ROS)、丙二醛(Maleicdialdehyde,MDA)、超氧化物歧化酶(Superoxide dismutase,SOD)、白介素6(Interleukin 6,IL-6)、白介素10(Interleukin 10,IL-10)、肿瘤坏死因子(Tumor necrosis factor,TNF-)及干扰素(Interferon-,INF-)水平,ELISA试剂盒均购自南京建成生物有限公司。
1.2.3 Real TimePCR
取预先制备血清,采用Trizol法提取各组受试者血清总RNA。PCR反应体系为20 uL,反应条件为预变性 95℃,l min;95℃ 15 s变性、58℃ 20 s退火、72℃ 45 s延伸,循环50次。根据熔解曲线剔除异常数据,导入相对定量公式(2-ΔΔCt)逐个计算样品基因的相对表达量,目标基因引物序列见表2。

表2 目标基因引物序列表
1.2 4Western Blot
1.3 统计学方法
采用SPSS 25.0统计学软件进行统计分析。数据以平均数±标准差(±)表示,两样本平均数间比较采用检验分析,多样本平均数间比较采用单因素方差分析(one way ANOVA)和两因素方差分析(one way ANOVA),组间比较采用LSD检验,<0.05表示有显著性差异。
2 研究结果
2.1 受试者Conconi运动心率反应
Conconi运动心率反应显示,世居高原受试者各级负荷心率均低于移居高原受试者(HA、HA、HA),同时随移居高原年限延长受试者各级负荷心率呈下降趋势。组间比较显示,负荷1、负荷3、负荷4、负荷5中HA组、HA组心率显著高于HA组、Con组;负荷2即刻,HA组、Con组心率显著低于HA组;负荷6即刻,Con组心率显著低于HA组,其余负荷条件下组间比较不存在显著差异。此外,由表3可知,移居高原受试者Conconi运动负荷1至负荷4心率反应均存在较为显著的突变特征。

表3 受试者Conconi运动心率反应表(beats/min-1)
2.2 受试者运动氧化应激水平
由表4可知,受试者运动氧化应激水平呈现出显著的高原适应时间依赖性。就血清GSH-px而言,Con组显著低于HA组及HA组(<0.05),HA组显著低于HA组(<0.05);就血清NO而言,HA组及HA组显著低于Con组(<0.05),HA组显著高于HA组(<0.05);就血清ROS及MDA而言,Con组及HA组显著低于HA组及HA组(<0.05),Con组与HA组比较不存在统计学意义(>0.05);就血清SOD而言,Con组及HA组显著高于HA组及HA组(<0.05),Con组与HA组比较不存在统计学意义(>0.05)。此外,受试者运动氧化应激适应特征显示。

表4 受试者运动氧化应激水平表
2.3 受试者血清炎症因子水平
由表5可知,世居高原受试者血清IL-6、TNF-低于移居高原受试者,世居高原受试者血清IL-10、INF-高于移居高原受试者,且移居高原受试者上述指标的变化特征存在时间依赖性。就血清IL-6而言,HA组、Con组显著高于HA组及HA组(<0.05);就血清IL-10而言,Con组显著高于移居高原受试者,且HA组、HA组及Con组显著高于HA组(<0.05);就血清TNF-而言,Con组显著低于移居高原受试者,HA组、HA组及Con组显著高于HA组,HA组及Con组显著低于HA组(<0.05);就血清INF-而言,HA、Con组显著高于HA组及HA组,HA组显著高于HA组(<0.05),其余组间比较无显著差异(>0.05)。

表5 受试者血清炎症因子水平
2.4 PI3K/Akt/GSK-3β/CREB通路相关mRNA表达
RT-PCR结果表明,受试者AKT mRNA表达随高原适应时间延长呈现下降趋势,其中HA组及Con组AKT mRNA表达显著低于HA组及HA组(<0.05),其余各组间比较不存在显著差异(>0.05)。就PI3K而言,HA3组及Con组PI3K mRNA表达显著低于HA组及HA组(<0.05)。就GSK-3而言,Con组GSK-3mRNA表达显著低于其余各测试组(<0.05),此外,HA组显著高于HA组及HA组(<0.05),HA组亦显著高于HA组(<0.05)。就CREB而言,Con组CREB mRNA表达显著低于HA及HA组(<0.05),HA组显著低于HA组(<0.05),其余各组比较不存在统计学意义(>0.05)。就ERK1/2/p-ERK1/2 mRNA表达而言,HA及Con组ERK1/2 mRNA表达显著低于HA及HA组(<0.05),HA组显著高于HA组(<0.05),其余各组比较不存在统计学意义(>0.05),见表6。

表6 PI3K/Akt/GSK-3β/CREB通路相关mRNA表达表
2.5 PI3K/Akt/GSK-3β/CREB信号通路蛋白表达
通过Western Blot检测受试者血清PI3K/Akt/GSK-3/CREB通路蛋白表达,结果见图2。就AKT/p-AKT蛋白而言,随着高原适应时间延长,高原适应受试者AKT/p-AKT蛋白表达呈下降趋势,HA组、Con组AKT及p-AKT蛋白表达均显著低于HA组及HA组(<0.05),Con组与HA组不存在显著差异,HA与HA组比较亦无统计学意义(>0.05)。就PI3K/p-PI3K蛋白表达而言,HA组、Con组PI3K蛋白表达均显著低于HA组及HA组(<0.05),其余各测试组不存在显著差异(>0.05);各测试组p-PI3K蛋白表达均不存在统计学差异(>0.05)。就GSK-3/p-GKS-3蛋白表达而言,Con组GSK-3蛋白表达显著低于HA组及HA组(<0.05),HA组显著低于HA组(<0.05);Con组及HA组p-GKS-3蛋白表达显著高于HA组,其余各组比较不存在统计学意义(>0.05)。就CREB/p-CREB蛋白表达而言,HA组、HA组CREB蛋白显著高于HA组及Con组(<0.05),其余各组比较不存在统计学意义(>0.05);各测试组p-CREB蛋白表达均不存在统计学差异(>0.05)。就ERK1/2/p-ERK1/2蛋白表达而言,随高原适应时间延长,受试者ERK1/2/p-ERK1/2蛋白表达均呈现下降趋势,HA组ERK1/2蛋白显著低于其余各测试组(<0.05),其余各组间比较不存在显著差异(>0.05);HA组及HA组p-ERK1/2蛋白表达显著高于HA组及Con组(<0.05)。
2.6 PI3K/AKT/GSK-3β/CREB通路相关蛋白磷酸化水平
PI3K/AKT/GSK-3/CREB通路相关蛋白磷酸化水平检测结果见图3。就AKT/p-AKT比值而言,Con组显著低于HA、HA及HA组(<0.05),且高原适应测试组间不存在显著差异(>0.05)。就PI3K/p-PI3K比值而言,HA组及Con组均显著低于HA组及HA组(<0.05),HA组显著高于HA组(< 0.05)。就GSK-3/p-GKS-3比值而言,HA组及Con组均显著低于HA组及HA组(<0.05),HA1组显著高于HA2组(<0.05)。就CREB/p-CREB比值而言,HA组及HA组均显著高于HA组及Con组(<0.05),其余组间比较不存在显著差异(>0.05)。就ERK1/2/p-ERK1/2比值而言,HA组及HA组均显著高于HA组及Con组(<0.05),其余组间比较不存在显著差异(>0.05)。
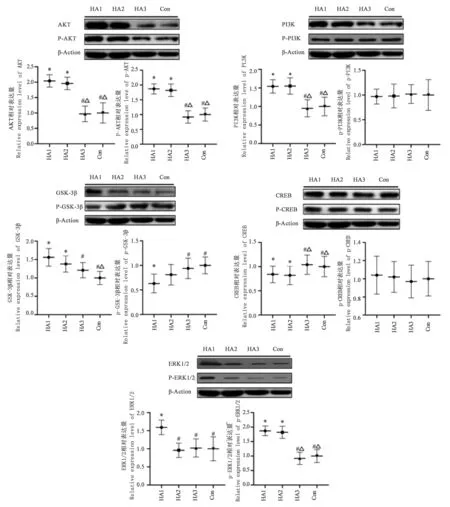
图2 PI3K/Akt/GSK-3β/CREB通路相关蛋白表达图

图3 PI3K/Akt/GSK-3β/CREB通路相关蛋白磷酸化水平图
3 讨 论
3.1 不同高原适应时相受试者运动心率反应
3.2 不同高原适应时相受试者运动氧化应激特征
3.3 不同高原适应时相受试者运动后炎症反应特征
本研究结果表明,世居高原受试者血清IL-6、TNF-低于移居高原受试者,世居高原受试者血清IL-10、INF-高于移居高原受试者,且移居高原受试者上述指标的变化特征存在时间依赖性。大量的研究表明,急性运动可引起众多致炎因子表达,其中INF-、TNF-表达最为明显,大量的致炎因子引起的炎性反应使炎性反应加重。研究发现,CD4+CD28+ Foxp3+ Treg细胞参与了运动疲劳及其恢复过程的免疫应答。运动应激下,人体及动物体内都观察到CD4+CD25+ Treg细胞数量激增现象。王成美等认为,单次大强度运动可导致机体血清TNF-明显升高。Boyle等报道,2~5 h跑测验和5 km赛跑后均可见血浆TNF-升高。现阶段,虽然血清IL-6对低氧环境反应的生理机制尚不明确,但一般认为,急性缺氧状态下机体血清IL-6可能是炎症或应激反应时相蛋白的重要介质,且血清IL-1、IL-1、TNF-和C反应蛋白水平呈现激增现象。研究证实,炎性因子可能通过激活花生油烯酸生成途径诱导ROC释放增加,从而激发机体氧化应激效应。部分研究显示,移居3 400 m高原6个月,安静状态下及定量负荷运动后机体炎症反应低于初上同海拔高原人群。但其具体机制并不明了,可能诱因如下:(1)热休克蛋白70(Heat shock protein70,HSP70)为中介的协同免疫效应增强。研究证实,周期性低氧预适应可能导致受试者HSP70表达上升,该现象可能介导NF-B p65、TNF-、IL-1表达,从而减轻低氧环境下的炎症亢进反应。(2)线粒体活性氧诱导低氧诱导因子HIF-1通路激活。虽然缺氧条件下线粒体适应确切机制还不清楚,但近年来随着线粒体动态过程、缺氧诱导因子1(hypoxia- inducible factor 1,HIF-1)和线粒体自噬受体等不同角度的相关研究。研究证实,线粒体活性氧可有效激活HIF-1活性,且线粒体功能在生理耐受范围内参与调节机体免疫反应。
3.4 PI3K/Akt/GSK-3β/CREB通路参与调控高原运动氧化应激、炎症反应的机制
最新的研究表明,氧化应激和炎症反应被认为是反映机体高原适应程度的关键因素。研究证实,急、慢性运动可能导致机体氧化活性水平发生适应性变化,不同环境、类型、持续时间和频率的运动对氧化应激有不同的影响。例如,部分研究显示,低氧环境下抗阻训练或有氧运动都可能诱导机体呈现氧化应激现象。Mller等研究证实,常氧和低氧(急进4 559 m高原3d)条件下力竭运动导致受试者DNA链断裂和氧化性DNA损伤现象加剧,活性氧诱发DNA链断裂和氧化性DNA损伤,线粒体呼吸阻断或缺氧诱导炎症反应。在以往的研究中,关于移居高原对人体或动物运动氧化应激损伤的研究较少,Wu等发现高原适应3年后力竭运动即刻受试者抗氧化能力较初上高原者显著提高,且机体脂质过氧化水平也呈现下降趋势,由此可见低氧适应可能是减轻高原运动氧化应激损伤的重要因素。上述研结论佐证了本研究结果。综上所述,课题组认为,PI3K/Akt/GSK-3/CERB通路可能是高原适应的潜在分子靶点。PI3K/Akt通路在细胞内外内刺激响应中发挥重要作用,并参与构成细胞内稳态细胞信号转导网络。此外该经典通路还参与细胞增殖、细胞代谢、蛋白质折叠及细胞自噬过程。PI3K的活化诱导质膜产生PTEN,这导致Akt及其激酶、依赖于PI的PDK1激酶重新募集到质膜上。Akt磷酸化糖原合成酶激酶3(GSK-3)的关键位点Ser9活性被抑制。GSK-3的功能亢进可能诱导NF-B等炎症因子水平上升,这一过程在氧化应激反应中起关键作用,并损害部分转录因子的转录活性,如cAMP反应元件结合蛋白(CREB)。既往研究证实,在短期低氧应激刺激下,Akt磷酸化一系列下游靶标,使GSK-3磷酸化水平下降并激活表达,CREB表达上扬,该种条件下从事中、高强度运动,Akt活化受到明显增强,磷酸化水平明显上升,GSK3失活也明显受抑制,本研究显示了类似的结果。部分研究显示,低氧适应现象强化了cAMP-CREB的级联效应,CREB被认为是潜在的缺氧适应靶点,通过CREB磷酸化(p-CREB)改善氧化应激效应。PI3K是受体酪氨酸激酶(RPTKs)和G蛋白复合受体的主要下游分子,激活的PI3K催化第二信使PIP3磷酸化及Akt生成。Akt通过其下游酶或转录因子(如糖原合成酶激酶-3,GSK-3(含GSK-3、GSK-3))实现自身激活,在调节细胞增殖、分化、凋亡、葡萄糖运输和代谢方面发挥重要作用。现阶段研究显示,GSK-3是催化蛋白磷酸化的重要酶;另一方面,GSK-3可通过磷酸化转录因子激活NF-B信号通路,但GSK-3不通过NF-B信号通路影响细胞增殖及代谢。这可能是低氧适应减轻机体高海拔运动氧化应激的途径之一。
4 结论与局限
不同周期高原低氧适应过程中,随着高原适应时间延长,受试者Conconi运动心率、氧化应激及炎症反应呈下降趋势,且上述指标趋于高原世居人群基本特征;AKT/p-AKT、PI3K、GSK-3、CREB/p-CREB及ERK1/2/p-ERK1/2 mRNA呈现上升趋势,p-PI3K及p-GSK-3呈下降趋势;此外,随高原适应时间延长,AKT、p-AKT、PI3K、GSK-3、ERK1/2及p-ERK1/2蛋白表达呈下降趋势,p-GSK-3蛋白表达呈上升趋势,且存在较为显著的时间剂量依赖现象;P-PI3K、CREB及P-CREB蛋白表达可能对高原低氧环境不敏感。
同时,研究尚存在以下不足之处:(1)囿于样本量纳入有限,上述研究结果尚待大数据研究的进一步验证;(2)受试者年龄、性别较为单一,缺乏全年龄段的横向研究数据。后续研究依赖于对高原适应的多年龄段多指标测试,以期系统、客观地评价高原适应人群运动氧化应激变化规律。
