微小核糖核酸495及微小核糖核酸200b基因多态性与大动脉粥样硬化型卒中发病年龄的关联分析
2022-10-03秦山梅沈冲汤武装林颖王昭君张治中刘新峰
秦山梅 沈冲 汤武装 林颖 王昭君 张治中 刘新峰
卒中已成为我国居民死亡的第3位病因[1]。70%以上的卒中是缺血性卒中,其中以大动脉粥样硬化型(large artery atherosclerosis,LAA)最常见,且该型更易受遗传因素的影响[2]。有研究证实,基因多态性与LAA缺血性卒中密切相关[3]。
微RNA(micro-ribonucleic acid,miRNA)可通过部分抑制信使RNA或降解靶向的信使RNA,使靶基因转录后发生基因沉默[4]。在缺血性卒中发生发展的过程中,存在大量特异性miRNA差异性表达, miRNA可能成为缺血性卒中诊断及对高危人群评估的生物学标志物。研究显示,miRNA-495及miRNA-200b参与了动脉粥样硬化、血小板聚集与活化、新生血管形成等病理生理过程[5-8]。miRNA-495及miRNA-200b的单核苷酸多态性(single nucleotide polymorphisms,SNPs)会导致相应的miRNA及其靶基因失调,进而影响缺血性卒中的发生发展[9]。目前,对于miRNA-495及miRNA-200b的SNPs与缺血性卒中发病年龄的相关性尚未明确,因此,本研究拟对miRNA-495及miRNA-200b的基因多态性与LAA缺血性卒中发病年龄的关联进行探讨,以期为缺血性卒中高危人群的病因预防及早期筛查提供依据。
1 对象与方法
1.1 对象
连续回顾性纳入2009年5月至2017年12月江苏省宜兴市人民医院神经内科LAA缺血性卒中住院患者697例,其中男429例,女268例;年龄39~82岁,平均(66.0±9.8)岁。本研究方案经东部战区总医院伦理委员会批准(伦理号:2017NZGKJ-041),患者或其家属签署了诊疗知情同意书。
纳入标准:(1)年龄≥18岁;(2)首次缺血性卒中,根据急性卒中Org 10172治疗试验(trial of Org10172 in acute stroke treatment,TOAST)分型诊断为LAA型[10],并经影像学证实。排除标准:(1)既往卒中、短暂性脑缺血发作史;(2)病历资料不完整;(3)合并有其他系统严重疾病。
1.2 研究方法
记录患者的基线资料,包括人口学特征(年龄、性别)、心脑血管疾病危险因素(高血压病、高脂血症、糖尿病)、不良嗜好(吸烟、饮酒史)。高血压病、高脂血症、糖尿病的评价分别参照国内相关指南的标准[11-13]。吸烟史及饮酒史通过查阅病历资料获得。
1.3 易感基因多态性检测及基因分型
通过NCBI dbSNP数据库及既往文献[9,14],筛选出与LAA相关的2个基因(miRNA-495及miRNA-200b)中的2个SNPs位点,分别为rs2281611及rs7549819。
入院后次日清晨采集所有患者的空腹外周静脉血5 ml,置于乙二胺四乙酸抗凝管中,离心后取下层红细胞,-80 ℃冰箱保存。由上海天昊生物科技有限公司采用SNPscan法对rs2281611、rs7549819进行基因分型。为评价基因分型的可靠性,随机挑选部分样本进行重复验证,检测结果符合率为100.0%。
依据rs2281611、rs7549819多态性基因分型,累计突变等位基因的个数,即患者基因型中所含2个SNPs突变等位基因(miRNA-200b C及miRNA-495 G)的个数之和。根据基因型中突变等位基因个数和,将患者分为3组,突变等位基因总数分别为0个(不携带突变等位基因)、1~2个、3~4个。
为了避免不同年龄段可能对结果带来的影响,本研究先对年龄进行了分层分析,miRNA-200b rs7549819在59岁患者中不同基因型交叉最多(Log-rankχ2=5.546,P=0.019;图1),并结合世界卫生组织的年龄划分标准[15],故本研究将LAA缺血性卒中患者分为中青年(年龄<59岁)、老年(年龄≥59岁)。
1.4 统计学分析

2 结果
2.1 基线资料比较
697例LAA缺血性卒中患者中,无高血压病史者平均发病年龄低于高血压病史者,有吸烟史及饮酒史患者平均发病年龄低于无吸烟史及饮酒史者,差异均有统计学意义(均P<0.05);其余基线资料平均发病年龄的差异均无统计学意义(均P>0.05)。见表1。
2.2 miRNA-495 rs2281611和miRNA-200b rs7549819多态性与LAA缺血性卒中发病年龄的关联
在miRNA-495 rs2281611的共显性模型中,TT、TG、GG型患者平均发病年龄的差异有统计学意义(F=4.224,P=0.015),与TT型和TG型患者相比,GG型患者平均发病年龄更早,两两比较的差异均有统计学意义(P值分别为0.036、0.004),TT型和TG型平均发病年龄的差异无统计学意义(P=0.639);在其隐性模型中,GG型患者平均发病年龄早于TT+TG型(P=0.004);在其显性模型中,TT型与TG+GG型平均发病年龄的差异无统计学意义(P>0.05)。miRNA-200b rs7549819各遗传模型中患者平均发病年龄的差异均无统计学意义(均P>0.05)。在共显性和隐性模型中,携带miRNA-495 rs2281611 GG型患者发生LAA缺血性卒中的发病风险更高(共显性:HR=1.40,95%CI:1.12~1.74,P=0.003;隐性:HR=1.35,95%CI:1.13~1.62,P=0.001)。见表2。对miRNA-200b rs7549819隐性模型分析显示,在年龄≥59岁的老年患者中,与TT+TC型相比,CC型LAA缺血性卒中发病风险增加了38.0%(HR=1.38,95%CI:1.08~1.77,P=0.009),且CC型患者发病年龄会更早(Log-rankχ2=9.579,P=0.002);中青年患者中CC型携带者LAA缺血性卒中发病风险则呈降低趋势,但差异无统计学意义(HR=0.73,95%CI:0.45~1.18,P=0.193),且与CC型患者发病年龄的差异无统计学意义(Log-rankχ2=1.651,P=0.199)。见图2。

表2 miRNA-495 rs2281611和miRNA-200b rs7549819多态性与697例大动脉粥样硬化型缺血性卒中发病年龄的关联分析

rs7549819 T>C基因型例数[例(%)]发病年龄(x-±s,岁)检验值P值HR值e95% CIP值共显性0.712c0.4910.161 TT282(40.5)66.0±10.01.00-- TC318(45.6)66.3±10.00.950.81~1.120.570 CC097(13.9)64.9±08.81.190.95~1.510.136显性0.085d0.932 TT282(25.3)66.0±10.01.00-- TC+CC415(74.7)66.0±09.71.000.86~1.170.982隐性1.152d0.250 TT+TC600(77.0)66.2±10.01.00-- CC097(23.0)64.9±08.8 1.230.99~1.520.068

2.3 miRNA-495 rs2281611和miRNA-200b rs7549819突变等位基因数量与LAA缺血性卒中发病年龄的关联
697例患者总体情况显示,随着携带累计突变等位基因数量的增加,LAA缺血性卒中患者平均发病年龄呈下降趋势,携带0、1~2、3~4个累计突变等位基因LAA缺血性卒中患者发病年龄的差异有统计学意义(F=4.108,P=0.017);与携带3~4个突变等位基因患者比较,携带0、1~2个突变等位基因LAA缺血性卒中患者发病年龄的差异均有统计学意义(P值分别为0.016、0.012),携带0个与1~2个突变等位基因患者发病年龄的差异无统计学意义(P=0.331)。与不携带突变等位基因患者相比,携带3~4个累计突变等位基因患者LAA缺血性卒中发病风险增加了41.0%(P=0.029)。对不同性别、年龄段LAA缺血性卒中患者发病年龄进行分析显示,携带3~4个累计突变等位基因的女性及老年患者LAA缺血性卒中发病风险显著增加(均P<0.05),但该结果在男性及中青年患者中差异无统计学意义(均P>0.05)。见表3。
3 讨论
本研究探讨了miRNA-495 rs2281611、miRNA-200b rs7549819多态性与LAA缺血性卒中发病年龄之间的关联,结果显示,携带miRNA-495 rs2281611 GG型患者的LAA缺血性卒中发病年龄更早,而miRNA-200b rs7549819 CC型与老年患者LAA缺血性卒中发病年龄提前相关,同时,这2个SNPs突变等位基因累计个数是影响LAA缺血性卒中发病年龄的独立危险因素,携带3~4个突变等位基因患者的LAA缺血性卒中发病风险显著增加,该结果在女性及老年患者中更显著。
大部分LAA缺血性卒中是由动脉粥样硬化斑块破裂及继发血栓形成所致,使脑血流灌注不足,从而引起相应缺血区的脑组织功能障碍。既往研究表明,miRNA-200b、miRNA-495在上述病理生理过程中参与了调控[6,16]。缺氧时,miRNA-200b可抑制内皮型一氧化氮合酶的表达,限制一氧化氮的生物利用度,内皮细胞代谢紊乱引起其功能障碍[5]。miRNA-200b表达水平升高会导致三磷酸腺苷结合盒转运体A1水平下调,加速了动脉血管壁细胞脂质积累,并抑制胆固醇逆转运,促进泡沫细胞形成,从而加速动脉粥样硬化的进展[16]。miRNA-495可抑制趋化因子配体2的表达,影响内皮细胞增殖和迁移通路,参与动脉粥样硬化的进程[17]。在血管再狭窄模型中,诱导miRNA-495表达沉默可抑制内膜增生,减少动脉血管壁细胞中巨噬细胞的数量,从而抑制动脉粥样硬化斑块的形成,并增加斑块的稳定性[6]。
血小板与动脉粥样硬化斑块破裂后继发的血栓形成密切相关[18]。miRNA-200b和miRNA-495可通过分别靶向蛋白激酶环磷酸腺苷依赖性Ⅱ型调节亚基β和Kelch样蛋白5,从而干扰血小板的激活和聚集[7]。与静息血小板相比,miRNA-495在受凝血酶刺激的血小板中表达水平上调[19]。miRNA-200b和miRNA-495失调会导致血小板表达水平及其反应性的改变,继而影响血栓形成的过程。
rs2281611、rs7549819的位点均位于基因启动子区,其发生突变能够直接影响miRNA-495和miRNA-200b基因的转录,并干扰突变位点相关转录因子的结合过程,基于上述miRNA-495和miRNA-200b在动脉粥样硬化进展[5-6,15-16]和继发血栓形成[7,18]等方面重要的调控作用,rs2281611、rs7549819位点突变可能会通过干扰相应miRNA及其靶基因的合成或调节过程,从而影响缺血性卒中的发生发展[9]。既往研究表明,老年人群大动脉粥样硬化负担随着年龄的增加而加重[20],而青年缺血性卒中的主要危险因素为吸烟、饮酒、腰臀比和心理社会因素等[21],且绝经后女性表现出更高的动脉粥样硬化斑块破裂的风险[22]。本研究结果与文献报道一致,提示rs2281611、rs7549819的多态性可能具有早期识别LAA缺血性卒中高风险人群的潜能。但rs2281611、rs7549819与LAA缺血性卒中患者发病年龄关系的具体生物学机制有待进一步探讨,本研究结果仍需后续的研究进行验证。
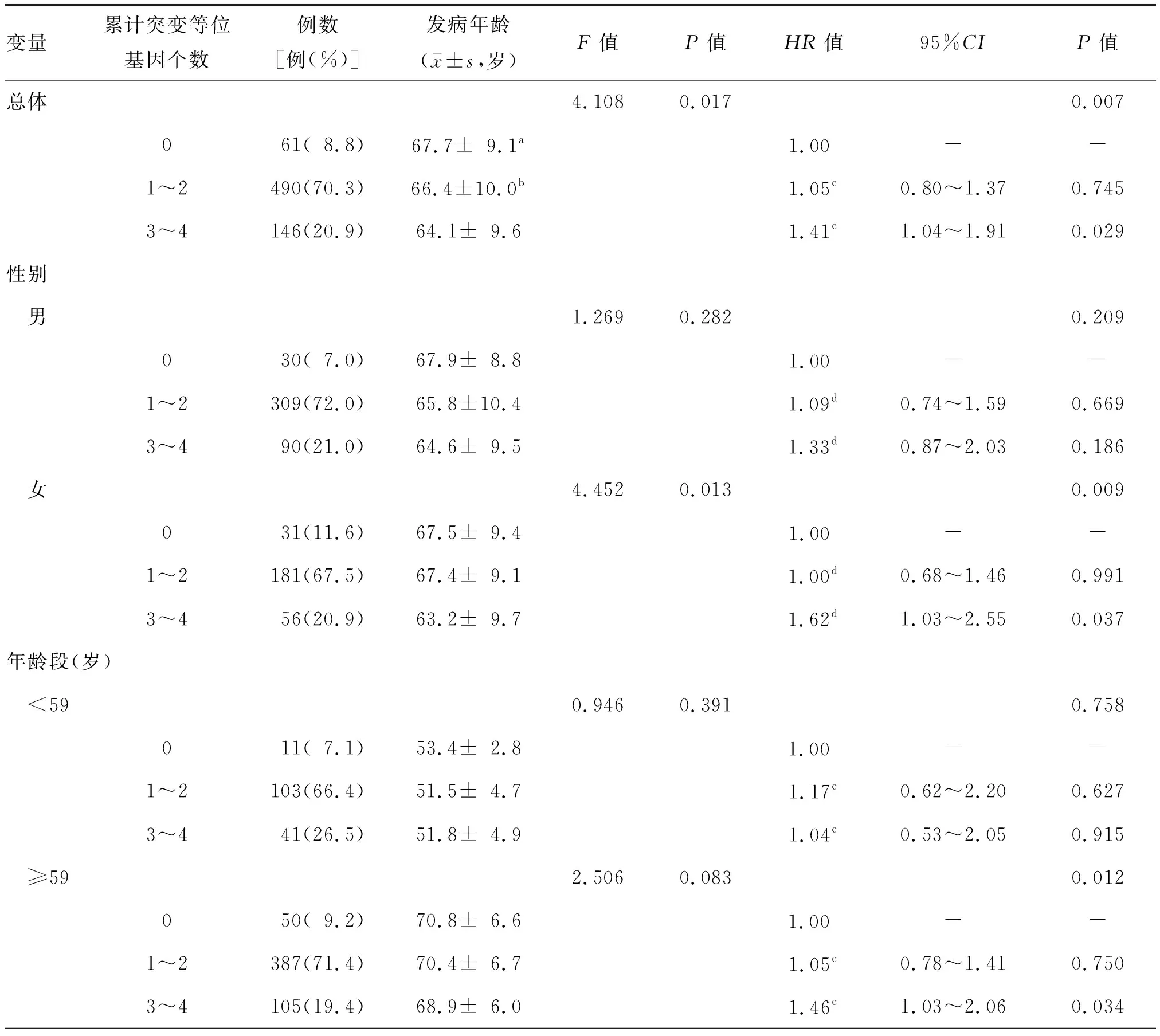
表3 miRNA-495 rs2281611和miRNA-200b rs7549819突变等位基因累计个数与697例大动脉粥样硬化型缺血性卒中发病年龄的关联分析
综上所述,miRNA-495 rs2281611、miRNA-200b rs7549819多态性及二者突变等位基因数量(3~4个)均与LAA缺血性卒中发病年龄提前相关,可能有助于早期识别LAA缺血性卒中高风险人群。
