味觉受体研究热点分析
2022-09-30张一纯陈艳红李利君
张一纯,陈艳红,2,3,李利君,2,3,倪 辉,2,3,*
(1.集美大学海洋食品与生物工程学院,福建 厦门 361021;2.福建省食品微生物与酶工程重点实验室,福建 厦门 361021;3.厦门市食品生物工程技术研究中心,福建 厦门 361021)
味觉是人们重要的感知反应,基本的味觉反应包括甜、酸、苦、咸、鲜5 种。近年来,一些科学家还发现了脂肪味和“加强味”的存在。错综复杂的味觉反应不仅能带给人们愉悦和美妙的感觉,还可以用来评估食物中化学物质的营养及安全特性。鲜味和甜味分别代表着食物富含蛋白质和能量;苦味和酸味可以帮助识别有害或腐败的食物;咸味有利于机体在饮食中保持电解质平衡。除此之外,味觉反应还与身体健康状况密切相关,很多疾病的发生都伴随着味觉感知的改变。
人们对味觉的感知依赖于味觉受体,食物风味成分和味觉受体相互作用,向机体发出信号,从而产生对食物的味觉响应。1999年人们发现了第一个味觉受体(taste receptor family 1 member 1,T1R1)基因,2001年美国科学家在人体第4对染色体中鉴别出一个与甜味感知相关的基因,2006年又报道人类基因组共编码了25 种苦味受体T2Rs。近年来,随着分子生物学、细胞生物学和神经生物学等技术手段突飞猛进的发展,人们对味觉受体的认知快速增长,味觉感知途径、呈味物质和受体的相互作用机制研究越来越深入,味觉受体领域正逐渐成为研究热点。
在Web of Science核心数据库中以味觉受体为主题词进行检索共得到1980—2020年间7 973 篇文献。本综述根据这些文献,对味觉受体热点关键词、期刊、来源、研究手段、受体类型、研究人员等进行分析概述,为开展味觉受体相关研究提供文献综述参考。
1 热点关键词及学术期刊分析
如图1A所示,味觉受体领域发表文献记录数(number of records,Recs)最多的前20 位常用词汇分别是苦味(bitter)、甜味(sweet)、酸味(acid)和鲜味(umami)等。来源包括大鼠(rats/rat)、人类(human)、小鼠(mice/mouse)和果蝇()。热点研究手段主要体现在细胞(cell)、表达(expression)、蛋白质(protein)、诱导(induced)、基因(gene)、信号(signaling)、神经元(neurons)、分子(molecular)等常用词汇。如图1B所示,受体感知味道的种类、来源及研究手段的Recs占比分别为29%、26%和41%。这说明用细胞学和蛋白质表达技术对鼠类和人类的苦味受体和甜味受体进行研究是当前的热点。文献本地引用次数(total local cite score,TLCS)表示在被处理的所有文献中的被引次数,图2显示了味觉受体领域发表文献Recs和TLCS排名均在前7 位的期刊,分别是、、、、、,和,其中包括、、和等著名期刊,均为中科院分区2区以上期刊,通过阅读这些期刊可以获取味觉受体领域的前沿信息。
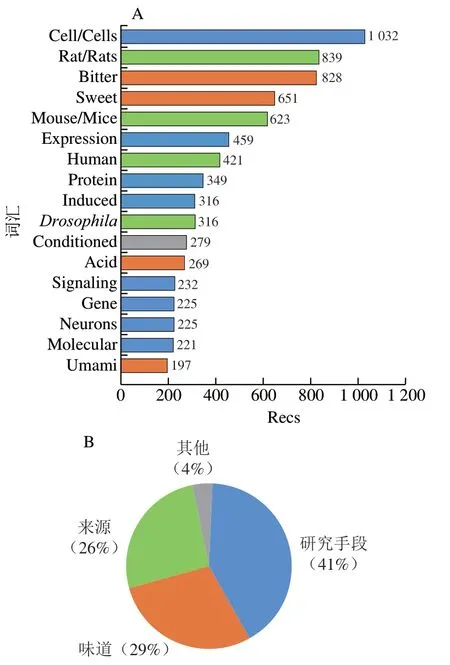
图1 味觉受体领域文献前20 位词汇Recs(A)和归类占比(B)Fig. 1 Top 20 lexical Recs (A) and their classification proportions (B)in the literature in the field of taste receptors
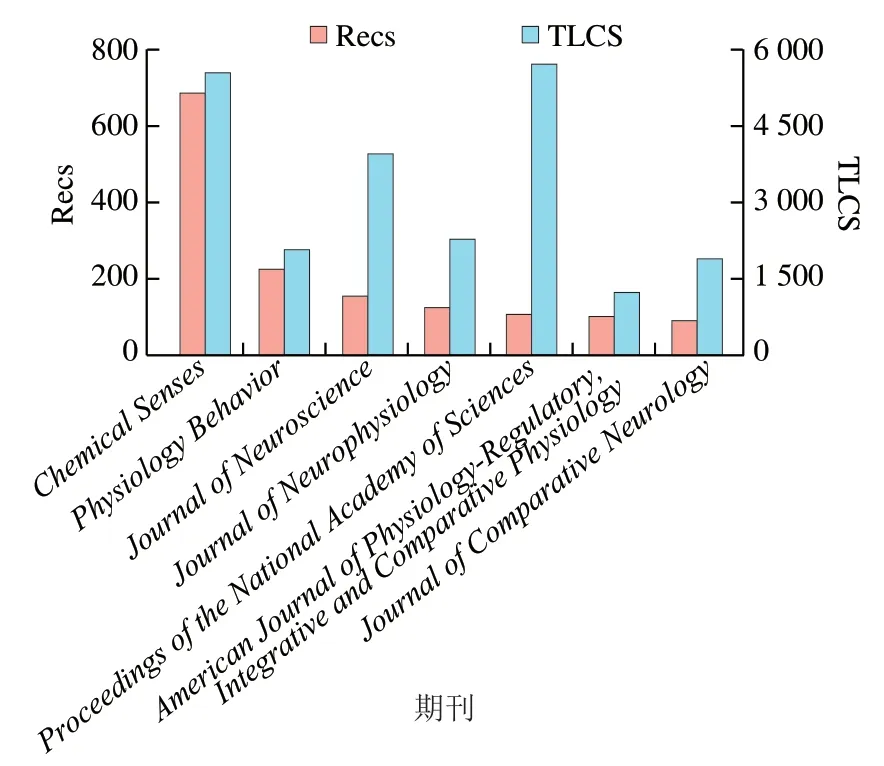
图2 味觉受体领域的重要期刊Fig. 2 Important journals in the field of taste receptors
2 热点来源分析
味觉受体来源的热点词汇分析表明,大鼠(rat/rats)和小鼠(mice/mouse)的味觉受体是科研工作中最常用的研究对象,其次是人类(human)和果蝇()。鼠类(包括大鼠和小鼠)和人类基因序列相似且操作简单,是最常用的实验材料;果蝇不仅和哺乳动物有相似的进食偏好,还可作为一个强大的遗传模型,应用复杂的分子遗传技术对其味觉的分子、生理和行为方面进行研究。科研人员利用鼠类味觉受体作为实验对象模拟患者出现的生理症状,用于研发治疗药物。大鼠实验证明,黄芪多糖可以改善高脂饮食损害的肠道甜味受体的信号传导,通过调整2型糖尿病(diabetes mellitus type 2,T2DM)大鼠肠道中的葡萄糖转运蛋白和甜味受体/胰高血糖素样肽/胰高血糖素样肽1受体途径表达,从而减轻T2DM大鼠的症状。除此之外,通过鼠类实验也验证了味觉受体和其他激素之间也存在关联性,例如唾液脂联素可以影响胃中的味觉受体抑制饥饿素的分泌等。人类味觉受体的基因型常用于探索预防和治疗肥胖等现代文明病的方法。例如在肥胖问题上,最新的一项研究表明肥胖与味蕾中基因表达量的改变有关,特别是肥胖病人中II型味觉细胞基因数减少;苦味受体T2R38 rs10246939与韩国人的饮食摄入量相关,具有TT基因型的女性会摄入更多的水果且肥胖风险也增加了1.75 倍。总的来说,味觉受体和肥胖具有关联性,深入研究味觉受体对肥胖症的预防和治疗具有重要意义。最近的研究发现,果蝇的味觉器官分布在全身各处,并且除了味觉受体基因家族的成员,很多化学感受受体也都参与了味觉的感知,包括离子型受体(ionotropic receptor,Ir)、囊袋(pickpocket,ppk)和瞬时受体电位(transient receptor potential,Trp)基因家族编码的受体,这些不同受体共同表达的模式可能是感知多种味道的基础。近年来,科研人员对果蝇味觉系统与控制摄食、交配和产卵之间的联系进行了研究,更加明确了味觉系统调节昆虫行为的方式。总之,鼠类味觉受体易操作并且与人类受体相似,是大多数味觉受体研究的实验对象。果蝇的味觉受体和哺乳动物的截然不同,通过分析果蝇的味觉受体可以为防治害虫提供更好的策略。然而,不同来源的味觉受体之间仍然存在较大差异,例如人工甜味剂如阿斯巴甜、纽甜、甜蜜素、新橙皮苷二氢查耳酮和甜味蛋白等甜味物质均不会被鼠类动物感知。为了更准确探索人类味觉受体,还需要对人类本身的味觉受体进行深入的研究。
3 热点研究手段分析
图1显示了8 个与味觉受体热点研究手段相关的词汇,其中表达(expression)、蛋白(protein)、诱导(induced)、基因(gene)和分子(molecular)属于分子生物学和结构生物学的研究手段,细胞(cell/cells)属于细胞生物学手段,除此之外,还包括了信号(signaling)和神经元(neurons)有关神经生物学研究手段。
目前,科研人员主要利用分子生物学手段使味觉受体表达并对其进行结构解析。哺乳动物表达系统可以使蛋白正确折叠并进行各种翻译后修饰,保证味觉受体完整行使其功能。其中甜味受体和鲜味受体具有两个亚基,要进行两个亚基的共表达。此外,科研人员将味觉受体成功地在细菌表达系统中表达,大量生产重组蛋白。解析蛋白质等生物大分子及其配合物的三维结构信息的主流方法包括X射线衍射晶体学、核磁共振以及冷冻电子显微镜技术,这3 种方法各有优点,在高分子结构分析中具有重要意义。在这些技术中,X射线衍射晶体学仍然是最广泛用于结构测定的技术。科研人员通过等温滴定量热法、荧光共振能量转移和小角度X射线散射法分析了青鳉鱼T1R2/T1R3,表明配体结合后异二聚体的构象发生转化。接着通过捕蝇夹结构域(venus flytrap domain,VFTM)与抗原结合片段(fragment antigen binding,Fab)(由识别T1R2 VFTM的抗体制备)共结晶,又解析出分辨率为2.2~2.6 Å的青鳉鱼T1R2/T1R3配体识别结构域,为理解味觉受体的化学感知提供了结构基础。近年来,许多科学家通过对内分泌细胞构建模型进行体外研究,常用的方法包括荧光共聚焦显微镜、单细胞显微切割捕获系统、免疫印迹法等,发现了甜味受体激动剂可以分泌与食欲相关的激素,如胰高血糖素样肽1、葡萄糖依赖性促胰岛素激素和直肠黏膜酪酪肽,并且甜味受体也被认为与调节胰岛素的分泌有关;我国上海交通大学近年来也发表了相关文献阐述了葡萄糖代谢与甜味受体的表达有关;美国斯克利普斯研究所提出苦味受体的激活导致肠内分泌激素释放,改善了代谢综合征的症状,表明味觉受体与人类健康息息相关。
科研人员利用神经生物学主要对味觉受体的传导过程及如何被感知进行了研究。当舌头味蕾上的味觉受体接受到味觉刺激,信息通过颅神经VII、IX和X从舌传递到脑干。如图3所示,依次通过孤束核(nucleus of solitary tract,NST)、脑桥臂旁核(parabrachial nucleus,Pbn)和腹后内侧丘脑(parvicellular portion of the ventroposteromedial nucleus of the thalamus,VPMpc)的小细胞部分,然后传递给初级味觉皮层岛皮质(insular cortex,IC)。研究表明,外周味觉系统中的味觉信息以标记线的方式编码,因此每种味觉模式都有独特的味觉受体和相应通路。其他研究者阐述了一种交叉纤维编码的方式,即包括大量感觉传入纤维的广泛共激活,相同纤维的不同组合构成了不同味觉品质的编码。电生理研究、双光子钙成像研究显示,一些味觉神经元只负责一种味觉,而其他神经元对多种味觉刺激有广泛的响应。东京大学重点研究了甜味信号的传导过程,通过细胞消融、体内钙成像和光磁实验表征了Pbn中表达富含AT序列特异性结合蛋白2(special AT-rich sequencebinding protein 2,SATB2)的神经元在选择性地将甜味信号传递到味觉丘脑中起重要作用;并对表达甜味受体突变体的所有细胞系进行了一系列细胞实验,分析了与抑制剂的相互作用,基于这些结果,进行了对接模拟、能量最小化,阐明了人类甜味受体抑制剂的具体结构机制。Yoshida等通过荧光共聚焦显微镜鉴定味蕾细胞,再由高阻抗膜片钳放大器记录电信号,证明了苦味敏感的味觉细胞具有异质性,这种异质性可能有助于区分苦味化合物。Lu Bo等通过化学刺激和电生理信号记录解释了温度对小鼠滋味感知的影响,其中温度对甜味和鲜味刺激的响应伴有瞬时感受阳离子通道家族M成员5(transient receptor potential cation channel, subfamily M, member 5,TrpM5)的参与,而对盐和酸的温度调节响应并不是通过上皮细胞钠离子通道和TrpM5的机制进行的,苦味刺激则受温度的影响较小。在人类口腔中有5 000 个味蕾,它们都位于舌头上表面、上颚和会厌,被分为I型(类胶质)细胞、II型(味觉受体)细胞、III型(突触前)细胞、IV型(基底)细胞和V型(边缘)细胞,I型、II型和III型细胞的协同作用是接受味觉的基础。如图4所示,I型细胞是味蕾中最丰富的细胞,由胶质细胞退化得到,顶端是钠钾离子泵,形成与盐传导有关的离子电流。对于II型细胞,钙离子成像显示其能够响应甜味、苦味和鲜味刺激,能与多种有机分子结合,识别出甜、咸、苦等复杂分子造成的味觉。III型细胞能被水和氢离子激活,直接对酸味刺激和碳酸溶液作出响应。除此之外,味觉受体不仅在口腔表达,通过对其他组织细胞研究发现T1Rs和T2Rs广泛表达于全身,分布于口腔外的受体和味蕾细胞中的受体信号通路相似,但无法感知味觉,而是具有营养传感、气道免疫防御和代谢过程调节等作用。嗅觉受体也同样分布于全身,最近的一项实验表明哺乳动物的味觉细胞上存在功能性嗅觉受体的表达。
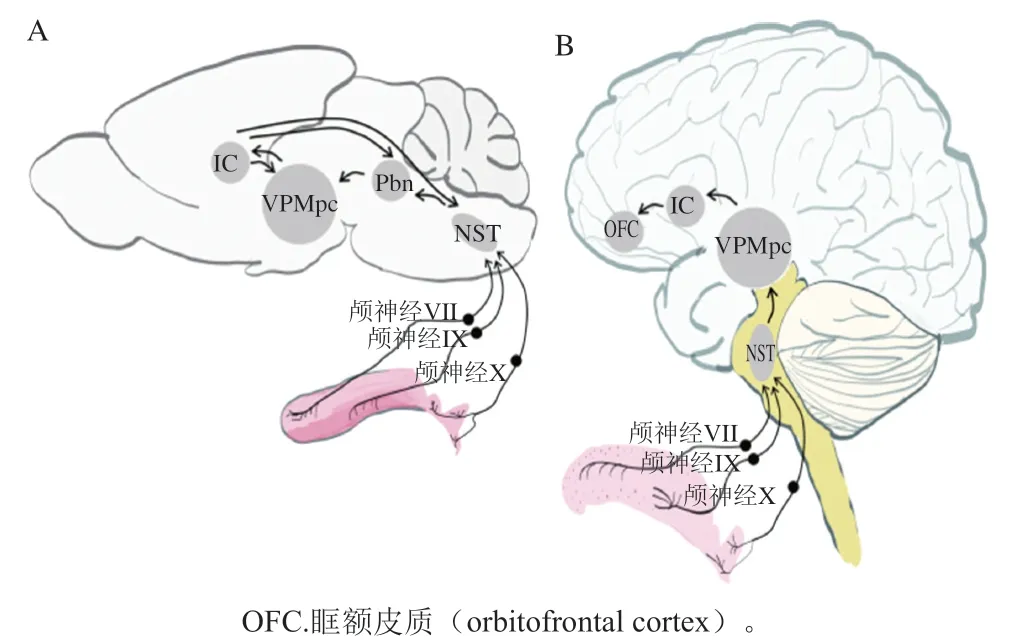
图3 啮齿动物(A)和哺乳动物(B)大脑感知甜味的味觉通路[32]Fig. 3 Taste pathways for sensing sweetness in rodent (A) and mammalian (B) brains[32]
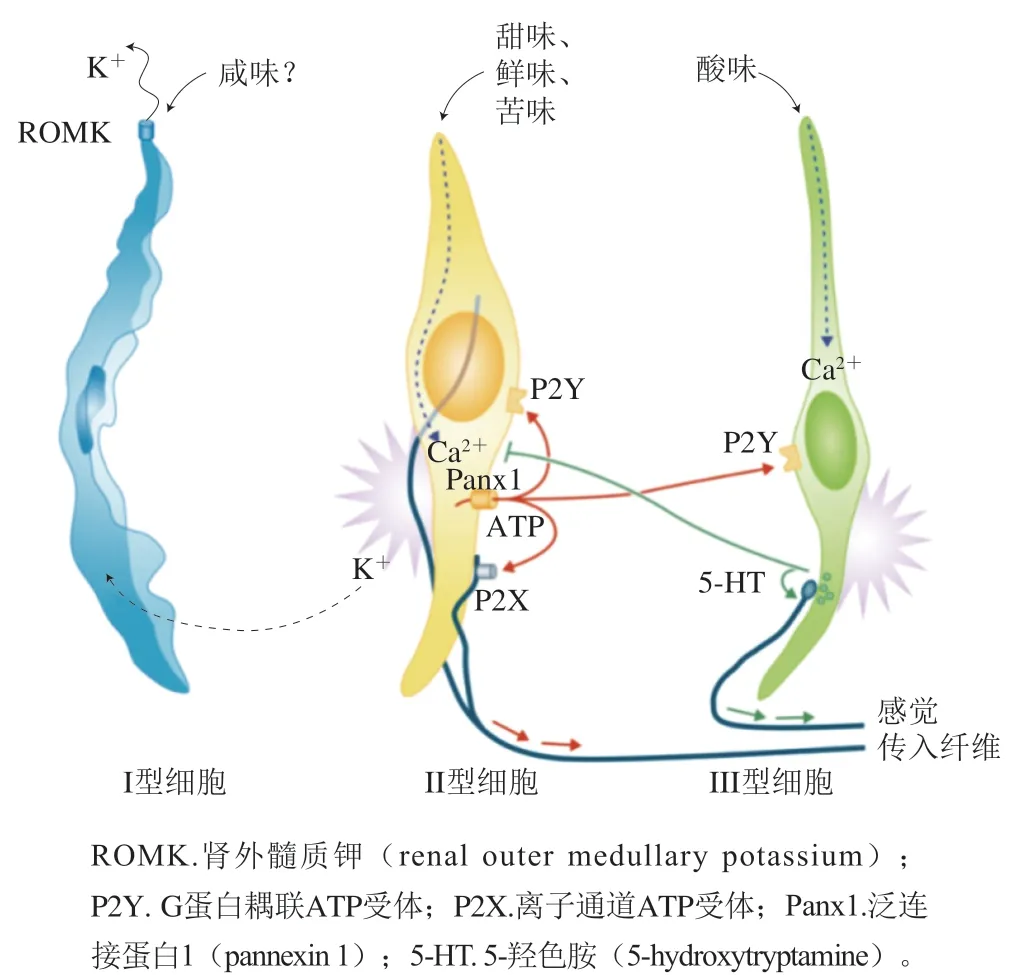
图4 三大类味觉细胞[44]Fig. 4 Three major classes of taste cells[44]
近年来,科学家们基于味觉受体的表达及神经传导领域的前沿技术,已经开始研发味觉受体方面的生物传感器。中国科学院侧重于研发体外生物电子舌来检测苦味或鲜味。莫奈尔化学感官中心和我国浙江大学生物传感器国家重点实验室共同提出将人苦味受体T2R16在HEK-293细胞中作为生物传感器的传感元件,可对水杨酸进行特异性检测。获取高纯度的蛋白质被认为是开发重复性强、性能稳定生物传感器的关键;同时,氧化铟锡(indium tin oxide,ITO)作为苦味传感器具有制备简单、廉价和灵敏的优点。将基于大肠杆菌表达系统的味觉受体与ITO构建的电化学传感器相结合,研发出的生物电子舌用于检测特定苦味物质,该方法具有简单和稳定的特点。此外,有些科研机构基于石墨烯效应晶体管构建双链生物电子舌,通过将人的鲜味和甜味受体在HEK-293细胞中表达,利用细胞松弛素B将其制成纳米囊泡,固定在微图案化的石墨烯表面,可用于同时检测鲜味和甜味。
总地来说,对味觉受体的探索已经进入分子生物学和神经生物学水平,但大多数受体的精确结构还未解析出,也难以区分不同特点的味觉神经刺激,目前主要采用计算机模拟和分子生物学实验结合等手段对受体结构进行探索。
4 热点受体研究进展分析
科研人员重点聚焦在苦味受体、甜味受体、酸味受体和鲜味受体上,其中苦味受体、甜味受体和鲜味受体均为G蛋白偶联受体(G protein coupled receptors,GPCRs),酸味则由Otopetrin-1(OTOP1)质子通道感知。如图5所示,GPCRs有相同的信号转导通路,配体与GPCRs结合以后,GPCRs构象发生变化激活异源三聚体细胞内G蛋白复合物,G蛋白的-和-亚基从活化的-亚基上释放,接着G蛋白的-和-亚基激活磷脂酶C-β2(phospholipase C-β2,PLCβ2)介导的肌醇1,4,5-三羟甲基氨基甲烷磷酸盐(inositol 1,4,5-trihydroxymethyl aminomethane phosphate,PIP2)生成甘油二酯(diacylglycerol,DAG)和1,4,5-三磷酸肌醇(inositol 1,4,5-triphosphate,IP3),IP3作用于IP3受体(IP3 receptor,IP3R),使内质网释放细胞内储存的Ca,激活TrpM4/5通道,引发Na内流和细胞去极化,随后细胞膜去极化激活电压门控钠通道(voltage-gated sodium channels,VGSC),最后激活钙稳态调节器1/3(calcium homeostasis modulator 1/3,CALHM1/3)通道释放ATP,导致位于感觉纤维中P2X2/P2X3的激活,触发动作电位,向味觉皮层发出信号以进行感知。
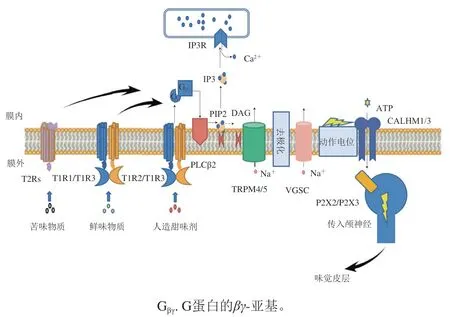
图5 舌头上的味觉受体细胞中甜味、鲜味、苦味的信号通路Fig. 5 Signaling pathways of sweet, umami, bitter in taste receptor cells in the tongue
苦味是5 种基本味道中最复杂的一种,苦味受体属于GPCRs的A亚型,由味觉受体第2家族(T2Rs)所介导,以单体形式存在。与其他味觉GPCRs相比,TAS2R家族是最多样化的并与多种激动剂结合。近年来,德国人类营养研究所发现了有助于感知二价锰离子或二价亚铁离子盐的苦味受体TAS2R7;根据同源建模和配体对接实验对人T2R14进行点突变增强了对不同激动剂的选择性。除此之外,该机构还提出人T2R14、T2R46在感知中药的苦味上发挥了主导作用,并证明琥珀酸衍生化作用可使T2R14苦味受体对愈创木酚甘油醚的响应消失。现在普遍认为,苦味受体存在于呼吸道、肠道、生殖系统和泌尿道,不仅可以用于检测有毒物质,而且可作为免疫“哨兵”防御致病性的侵略。宾夕法尼亚大学提出苦味受体T2R38功能的变化与慢性鼻窦炎的疾病状态和疾病严重程度相关;T2R14作为上呼吸道感染的潜在治疗靶点,其激动剂黄酮物质可能具有局部治疗的临床潜力。Ayabe等报道了啤酒中的异--酸和成熟啤酒花酸可能与肠道分泌细胞中的苦味受体结合,从而改善认知功能。此外,研究表明和基因可能与人类的长寿有关。对于苦味受体而言,有效的药物开发很大程度上取决于对配体和受体结构-功能关系的研究,目前仍不清楚苦味受体的真实结构,主要通过同源建模、对接模拟、功能性实验和突变体结合来获得。除此之外,近年来我国的科学家也将目光投向了研究苦味受体和物种进化的关系上,中国科学院提出大熊猫和小熊猫由于饮食习惯造成了苦味基因的特异性进化,武汉大学则探究出功能性苦味受体的数量与禽类饮食中大量潜在毒素水平之间存在正相关。这些报道为研究鸟类和哺乳动物感觉进化的分子基础以及对生态环境的适应过程提供了新思路。
甜味受体T1R2/T1R3和鲜味受体T1R1/T1R3属于GPCRs的C亚型,味觉受体第1家族(T1Rs),以异二聚体形式存在,如图6所示,T1R2/T1R3的结构包括VFTM、半胱氨酸富集区(cysteine rich domain,CRD)、跨膜区(transmembrane domain,TMD)和胞内结构域(intracellular domain,ID),T1R1/T1R3与之类似。甜味受体和鲜味受体两者共享一个亚基T1R3,与T1R3结合的甜味剂——甜蜜素虽然不能激活鲜味受体,但能增强谷氨酸盐诱导活性,是T1R1/R1R3的正变构调节剂,甜味抑制剂Lactisole则是T1R1/T1R3的负变构调节剂。除了人工甜味剂的传导途径外,甜味还涉及不同的信号传导途径(图7)。蔗糖等糖类物质与T1R2/T1R3结合,使G蛋白的-亚基(G)活化,从而激活腺苷酸环化酶(adenylyl cyclase,AC),产生3’,5’-环腺苷酸(cyclic adenosine monophosphate,cAMP),导致胞内cAMP浓度上升,cAMP直接激活蛋白激酶A(protein kinase A,PKA),引起K通道的磷酸化,导致离子通道关闭,抑制K外流,引起膜去极化和神经递质释放。甜味受体已经被发现在大脑、胃肠道、肾脏和脂肪等组织中均有表达,受体不仅影响着代谢过程,如胰岛素分泌、糖代谢和脂肪代谢,还与包括T2DM在内的慢性炎症疾病有关。慢性代谢疾病患者具有较高的与饮食行为相关的等位基因频率,他们对富含甜味的食物更加喜爱。随着人们健康意识的提高和工作性质的转换,低热量、无热量的甜味剂在食品领域得到广泛使用。然而多数甜味剂甜味不纯正,呈现速度和持续时间与蔗糖相比有差异,存在不良“异味”,导致产生不愉悦的口感。为了满足消费者对减糖和口感的双重需求,科研人员运用构建嵌合体、计算机模拟来探究甜味受体与配体相互作用从而设计新型甜味剂。鲜味受体方面的热点聚焦在鲜味肽/鲜味增强肽的呈味规律和变构调节。除T1R1/T1R3外,目前报道的代谢型谷氨酸受体(metabotropic glutamate receptors,mGluRs)和细胞外钙受体(extracellular calcium sensing receptor,CaSR)等也可以感知鲜味且均为GPCRs。不同的鲜味剂之间具有协同作用,例如在含谷氨酸的食物中添加游离核苷酸如5’-磷酸鸟苷后,鲜味会增加数倍。除此之外,鲜味剂还与其他味觉感知受体具有相互作用,例如鲜味肽通过人类苦味受体抑制苦味。2020年,Spaggiari等发表了一篇研究鲜味、甜味和苦味受体的计算机分子建模技术的完整概述,该方法已用于味觉感受器建模、模拟促味剂-受体结合方式以及筛选潜在促味剂等方面。近年来研究人员提出了几种机器学习模型,用定量构效模型来预测分子的甜度。最新的一篇文献报道了通过结构鉴定和分子对接从天然库中筛选潜在的甜味剂。目前,主要通过建立可靠的突变实验、构建嵌合体、数学建模、分子对接、电生理技术、动物味觉模型来深入研究有关受体分子的作用机制。然而甜味受体和鲜味受体的结构和微观作用规律以及呈味规律还无法准确的阐明,因此,采用冷冻电子显微镜等技术解析甜味受体的精细结构是本领域的研究方向。
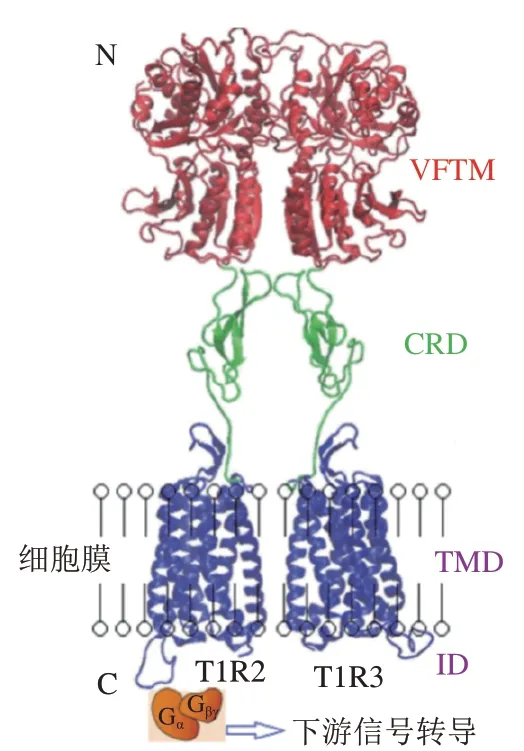
图6 甜味受体T1R2/T1R3的结构示意图[27]Fig. 6 Schematic diagram of sweet taste receptors[27]
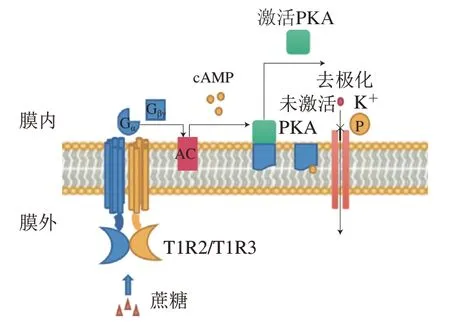
图7 甜味受体感知蔗糖的信号传导通路Fig. 7 Signaling pathways of sucrose sensing by sweet taste receptor
酸味一直被认为是5 种基本味觉之一,该味觉检测途径的发现历经坎坷。早期,科学家提出了、、等候选基因可能与酸味感知有关,它们(特别是)在酸敏感离子通道或在酸敏感味觉受体细胞中富集表达。科学家一直致力于确认这种受体以及其他可能的酸味受体的作用机制和功能;并发现即使缺乏假定的酸味受体细胞,动物对酸仍然表现出强烈的厌恶,这为确定酸味受体的功能基因带来了很多困难。因此,酸味受体的研究进展缓慢,直到2019年才发现了真正的酸味检测通道OTOP1。近年来,科学家解析了斑马鱼OTOP1的结构,阐明了其转导机制,发现强酸和弱酸的传导机制完全不同,强酸的H通过对Zn敏感的H通道进入细胞,而弱酸以未解离的中性分子进入细胞再解离形成H。如图8所示,H进入细胞引起细胞内的酸化,阻断了K2.1 K通道,细胞膜去极化,电压门控Na通道开放,引起一系列动作电位开放电压门控Ca通道,神经递质产生神经冲动引起酸味的感知。目前在破解传入大脑途中处理信息的神经回路方面已经取得了一些进展,但其传导通路中仍有许多环节有待研究。
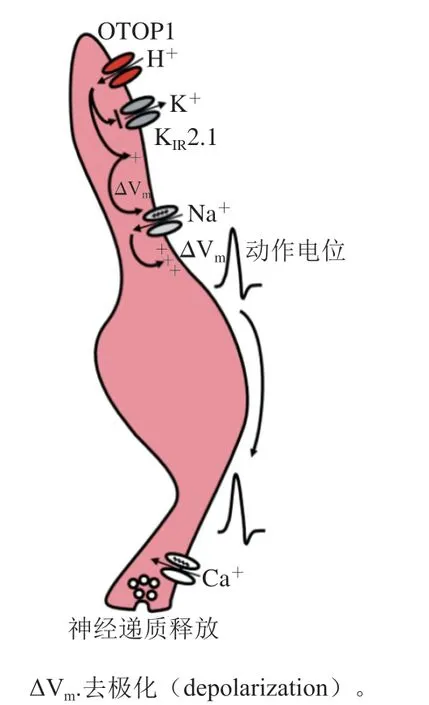
图8 酸味受体的信号传导通路[105]Fig. 8 Signaling pathways of sour taste receptors[105]
总而言之,味觉受体的结构特征和相关机制都尚未明确,该领域还需要更多的科学支持和技术支持。新型滋味物质的设计、味觉的感知机制、受体和健康的关系及个体感知的差异方面将不断吸引着科学家的关注,是未来的探索方向。
5 热点科研机构及科学家分析
如图9所示,Recs排在前5 名的国家分别是美国、日本、德国、英国和中国;其中美国的Recs占全部Recs的44%,遥遥领先其他国家。TLCS排名前10的科研机构中有9 个来自于美国(图10)。其中加州大学圣迭戈分校发表的文献被引次数最高,该机构很早就致力于研究哺乳动物口腔感知鲜味、甜味和苦味的机制,发现了第一个味觉受体T1R1,并利用外源表达证明了味觉蛋白T1R2和T1R3结合感知甜味、苦味的编码独立于甜味和鲜味等基础内容。莫奈尔化学感官中心提出苦味受体TS2R38的多态性编码对包含N—C=S片段的苯硫脲和丙硫氧嘧啶化合物表现出不同的响应程度,这种多态性可能来源于进化压力;苦味受体不仅可以防止有害食品的摄入,T2R38还在上呼吸道中表达并发挥“哨兵”的功能;该机构还提出甜味、苦味和鲜味传入大脑的方式是通过CALHM1离子通道释放ATP,最终激活传入神经通路。美国国立卫生研究院牙科与颌面研究中心发表的文章对味觉受体研究有着深远的影响,推动了味觉受体领域的发展,该中心的研究团队很早就开始研究哺乳动物味觉受体的结构,相关文献均发表在1999—2018年,2002年其发现了甜味和鲜味受体有一个共同的亚基;2004年发现了在甜味受体上存在多种配体结合位点,证明了T1R3和T1R2的不同功能。科罗拉多大学主要致力于研究味觉传导机制,发现味觉传导过程包括ATP信号传导和钙离子传导,同时发现II型细胞是苦味、甜味和鲜味味觉转导的主要转导者,可能通过光滑内质网下池与神经系统通信,以取代传统的突触。迈阿密大学提出受体细胞在接受味觉刺激后通过Panx1分泌ATP,并进一步刺激其他味觉细胞释放血清素,这一发现阐明了味蕾细胞间信号处理的方式,在味觉受体信号传导方面作出了巨大贡献;2015年该机构还提出脂肪组织产生的瘦素能够特异性作用于甜味,对苦味诱发的递质释放没有影响。
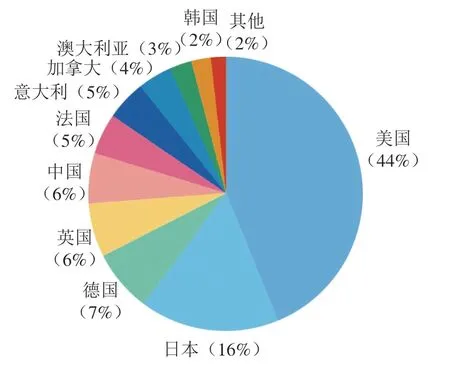
图9 以发文量Recs为指标排名前10的国家/地区及Recs比例Fig. 9 Top 10 countries/regions by the number of publications and their percentages in total Recs

图10 以文献TLCS为指标排名前10的研究机构Fig. 10 Top 10 research institutions by literature TLCS
以TLCS进行排序,排名前5的高产作者也均来自于美国(表1),其中Ryba N. J. P.、Hoon M. A.和Chandrashekar J.来自加州大学圣迭戈分校,共同发表了Mammalian sweet taste receptors、T2Rs function as bitter taste receptors、A novel family of mammalian taste receptors等影响力较大的文献,研究重点主要是哺乳动物甜味、鲜味和苦味的感知过程,排在第三名的霍华德·休斯医学研究所的Zuker C. S.也参与了上述几篇文献的实验研究。排名第一的作者Margolskee R. F.在阐述人类、鼠类的味觉感知机理以及诊断、治疗相关疾病方面做出巨大贡献,Margolskee R. F.研究了味觉蛋白T1R2、T1R3对甜味剂的响应和对胰岛素分泌的调控机理,这对预防和治疗吸收不良综合征和包括糖尿病和肥胖症在内的饮食相关疾病具有意义。总地来说,目前味觉受体领域的研究主要集中在美国的加州大学圣迭戈分校,Ryba N. J. P.、Hoon M. A.和Chandrashekar J.等科学家的研究处于领先位置。
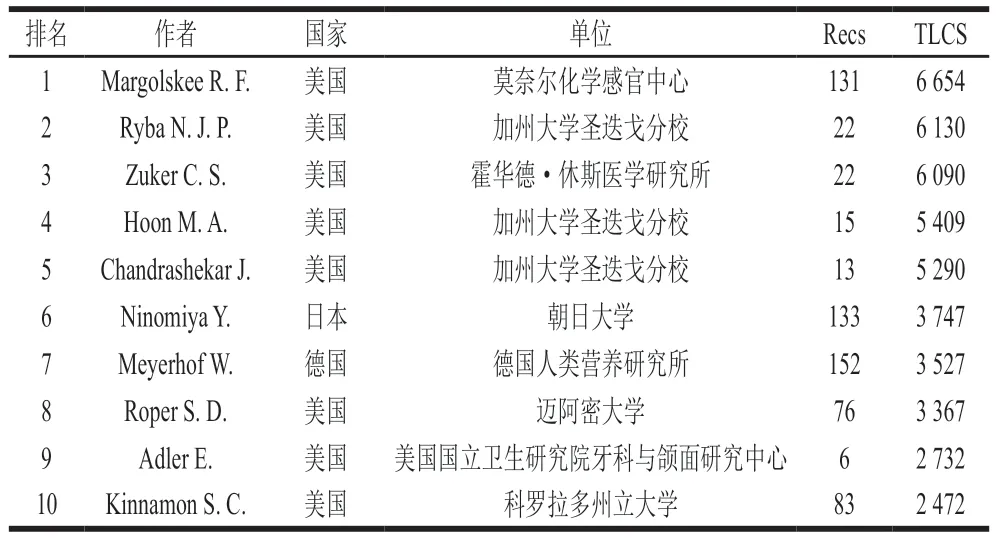
表1 Web of Science味觉受体领域文献TLCS排序前10 名作者分布Table 1 Top 10 authors by TLCS of literature on taste receptors in Web of Science
6 结 语
味觉受体不仅存在于口腔,还遍布全身各处。口腔外的组织表达的味觉受体无法感知味觉,而是具有营养传感、气道免疫防御和代谢过程调节等作用,被认为是重要的药物靶点,与呼吸道疾病、心脑血管疾病、糖尿病和肥胖等疾病相关,通过鼠类实验模拟T2DM患者,发现黄芪多糖对T2DM症状有改善作用,黄酮则对治疗上呼吸道感染疾病具有临床潜力,因此开发用于治疗疾病、减轻症状的小分子将成为一个非常活跃的研究领域。但由于获得单晶困难,大多数受体的精确结构还未解析出,也难以区分不同特点的味觉神经刺激,目前主要采用计算机模拟(包括同源建模、分子对接等手段)构建受体模型,将现有模型与实验数据进行比较,促进受体结构模型从低分辨率到高分辨率的过渡。虽然近年来我国在味觉受体尤其在生物传感器和受体进化机制方面的研究取得很大的进展,但在味觉受体领域做出主要研究贡献的国家还是美国,其中杰出的科学家包括加州大学圣迭戈分校的Ryba N. J. P.、Hoon M. A.和Chandrashekar J.等,我国需要与排在前列的国家、科研机构、科学家加大沟通交流。未来应深入加强味觉受体领域相互作用机制和信号传导途径方面的研究,攻克蛋白质表达纯化和结晶方面的难题,探究进化和多态性的分子机制,同时通过将味觉受体和化学材料等多种学科结合增强传感器稳定性。这些研究都有利于研制传感器在模拟人类味觉、药物筛选、研发新型增味剂和掩蔽剂、探索作为治疗疾病的药物靶点等方面的应用。
