黄大茶水提物改善高脂小鼠脂肪组织的脂肪酸代谢
2022-09-30陆凤娟沈钰珠董荣荣周德杰
陆凤娟,许 娜,*,沈钰珠,董荣荣,汪 闽,周德杰,储 俊
(1.安徽农业大学茶与食品科技学院,茶树生物学与资源利用国家重点实验室,安徽 合肥 230036;2.安徽中医药大学,新安医学教育部重点实验室,安徽 合肥 230038)
皖西黄大茶主产于安徽霍山,属轻度发酵的黄茶。与绿茶采用嫩叶原料不同,黄大茶加工原料为一芽三、四、五叶的带茎鲜叶。独特的闷黄和拉老火(高温烘焙)工艺,造就了黄大茶“黄叶黄汤”的品质特征,形成独特的烘焙香、焦糖香和“锅巴香”,类似于咖啡/可可的烘焙香气。本课题组前期开展黄大茶、绿茶、红茶和黑茶的对比研究,结果显示黄大茶对高脂饮食诱导小鼠的降血糖作用优于其他3 种茶。在糖尿病模型(C57BLKsJ-/)小鼠中,黄大茶也显示出显著降低餐后血糖的功效。此外,在C57BL/6小鼠模型中,黄大茶能改善巨噬细胞相关的慢性炎症及代谢综合征。
肥胖是一种慢性代谢性疾病,常伴有2型糖尿病、脂肪肝等疾病。脂肪组织功能障碍和异位脂肪沉积是决定个体患肥胖症及代谢综合征的重要因素。在肥胖等病理条件下,脂肪组织功能失调(受脂肪细胞扩张限制,其细胞体积达到临界阈值),过剩的能量超过脂肪组织的最大储存能力,导致在异位组织(肝脏、骨骼肌和心脏)中积聚,而脂肪酸的异位积累是造成非酒精性脂肪肝的主要因素。在肝脏中从血浆摄取和从头生物合成的脂肪酸,通过细胞内的氧化和与极低密度脂蛋白结合的形式分泌到血浆中。肝脏是脂肪酸代谢的核心器官,尽管通过肝脏产生脂肪酸的途径通量很高,但在正常情况下,肝脏仅存储少量的脂肪酸。
脂肪组织是脂肪酸储存的主要部位,也是脂肪酸代谢的另一个重要器官。本课题组前期在研究/小鼠时发现,黄大茶能够显著抑制肝脏脂肪酸的合成,促进脂解途径,从而抑制肝脏脂肪变性。在3T3-L1细胞模型中也发现黄大茶干预能够降低脂肪细胞的脂质积累。然而黄大茶对小鼠脂肪组织中脂肪酸合成与分解的影响仍未明确。本研究采用高脂饮食诱导C57BL/6肥胖小鼠模型,探究黄大茶水提物(large-leaf yellow tea aqueous extracts,YT)对脂肪组织中脂肪酸代谢的调控机制,以期为开发黄大茶相关的降糖、降脂产品提供理论依据和应用参考。
1 材料与方法
1.1 动物、材料与试剂
SPF级雄性C57BL/6小鼠(4 周龄)购自北京维通利华实验动物技术有限公司(生产许可证号:SCXK(京)2012-0001)。
黄大茶购自安徽省霍山抱儿钟秀茶业有限公司。
Mayer’s苏木素、伊红 上海索莱宝生物公司;TRIzol试剂、SYBR Green混合物 南京诺唯赞生物科技有限公司;逆转录试剂盒 北京全式金生物技术有限公司;实时荧光定量PCR引物 上海生工生物技术有限公司;-actin、腺苷酸活化蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)1α、磷酸化AMPK1α(p-AMPK1α)、乙酰辅酶A羧化酶(acetyl CoA-carboxylase,ACC)、磷酸化ACC(p-ACC)单克隆抗体 艾博抗(上海)贸易有限公司;RIPA裂解液、BCA蛋白浓度测定试剂盒 上海碧云天生物技术有限公司;增强型化学发光(enhanced chemiluminescenc,ECL)显色液 武汉博士德生物工程有限公司;其他化学试剂为分析纯。
1.2 仪器与设备
石蜡自动切片机、光学显微镜 德国Leica公司;荧光定量聚合酶链式反应(polymerase chain reaction,PCR)仪、酶标仪、蛋白电泳仪、凝胶成像仪 美国Bio-Rad公司;高效液相色谱仪 美国Agilent公司。
1.3 方法
1.3.1 YT的制备
按照本课题组之前报道的方法制备茶水冻干粉,主要步骤如下:取干燥茶叶100 g粉碎,按照1∶30(/)添加煮沸的蒸馏水,自然冷却至70 ℃后,将其转移到70 ℃的旋转蒸发仪中处理15 min,重复以上步骤2 次,合并冷凝液及旋蒸后过滤的上清液,冷冻干燥制得YT冻干粉。每100 g干茶叶约获得YT 23.6 g。经高效液相色谱检测YT含有茶多糖9.7 mg/g、可溶性糖19.1 mg/g、茶多酚109.9 mg/g,主要活性单体没食子儿茶素(gallocatechin,GC)7.46 mg/g、没食子儿茶素没食子酸酯(gallocatechin gallate,GCG)9.96 mg/g、表儿茶素(epicatechin,EC)1.41 mg/g、表儿茶素没食子酸酯(epicatechin gallate,ECG)4.98 mg/g、表没食子儿茶素(epigallocatechin,EGC)11.38 mg/g、表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)29.22 mg/g、没食子酸9.95 mg/g、咖啡碱63.91 mg/g、可可碱0.10 mg/g。
1.3.2 小鼠饲料的制备
对照组饮食(LFD)配方基于美国实验动物饲料公司Research Diets(http://www.researchdiets.com)D12450B,能量16.11 kJ/g,其中脂肪占能10%。高脂饮食(HFD)配方基于Research Diets D12451,能量19.79 kJ/g,其中脂肪占能45%。饲料由南通特洛菲饲料有限公司合成,详细成分组成见表1。前期研究在高脂饮食饲料中分别添加终质量分数为2.5%、0.5%的YT,结果显示两种剂量对C57BL/6雄性小鼠无肝毒性及任何其他不良反应。与高脂饮食饲喂的小鼠相比,HFD+2.5% YT组小鼠肝脏和脂肪组织中的脂质沉积均显著降低,组织和循环系统中的炎症水平得到改善。因此,本研究继续沿用此有效剂量。
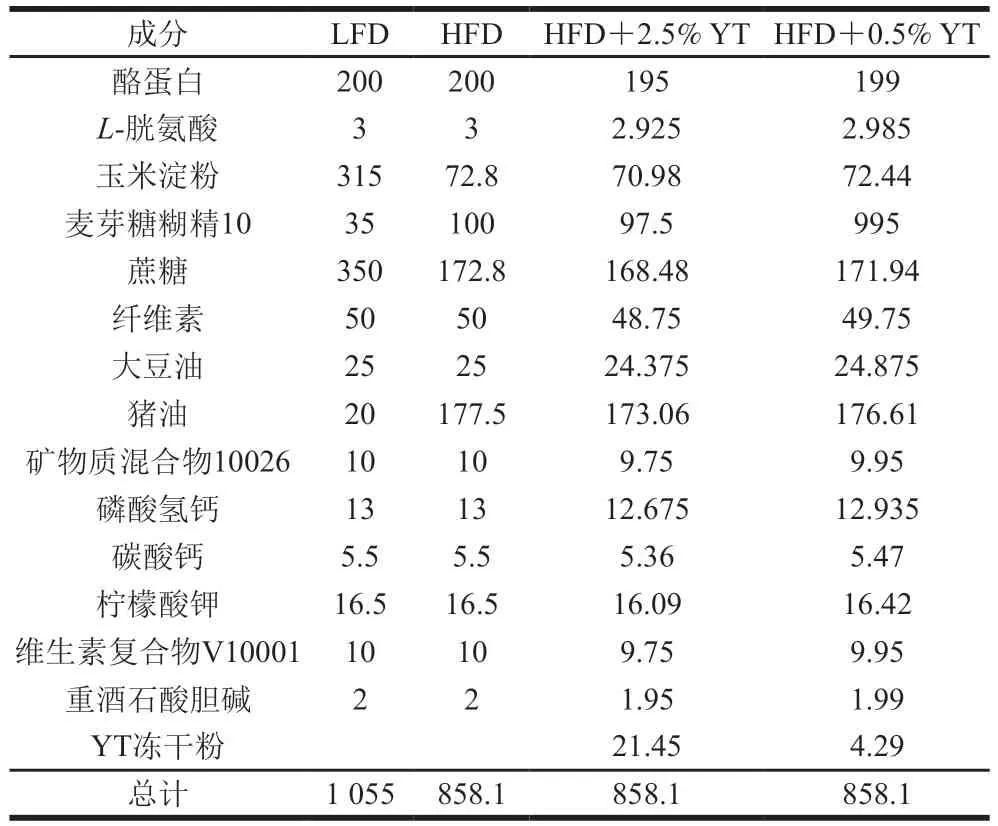
表1 实验动物饮食配方[6]Table 1 Composition of diets used in the experiment[6] g
1.3.3 小鼠分组处理和样本采集
小鼠在安徽农业大学SPF级动物房中适应1 周后,随机分4 组(每组6 只):LFD(对照饮食),HFD(高脂模型饮食)、HFD+2.5% YT(高剂量茶饮食)和HFD+0.5% YT(低剂量茶饮食)。保持12 h光照/12 h黑暗周期,允许小鼠自由进食、饮水。所有小鼠实验方案经安徽农业大学动物伦理委员会批准(编号:AHAU2016001)。
每周监测小鼠体质量、饮水量和食物摄入量。于第12周结束时禁食过夜,CO处死小鼠。解剖小鼠,分别取棕色脂肪、皮下脂肪和附睾脂肪组织,拍照、称质量。将组织一部分用体积分数4%多聚甲醛溶液固定备用;另一部分液氮速冻,-80 ℃保存。
1.3.4 组织病理学分析
取固定后组织,按照常规方法脱水、包埋、制备蜡块、切片、苏木精-伊红染色。染色片于光学显微镜下观察组织形态,拍照记录并用Image-Pro Plus 6.0软件统计细胞面积。
1.3.5 PCR检测脂肪生成、分解相关基因表达量
TRIzol法提取组织总RNA,检测RNA的浓度及质量,反转录获得cDNA,-80 ℃保存。采用10 μL PCR体系:2×PowerUp SYBR Green Master Mix 5 μL、上游引物(10 nmol/L)0.2 μL、下游引物(10 nmol/L)0.2 μL、cDNA模板1 μL、ddHO补充至10 μL。以-为内参基因,采用ΔΔCt阈值循环法计算基因表达水平。引物序列如表2所示。
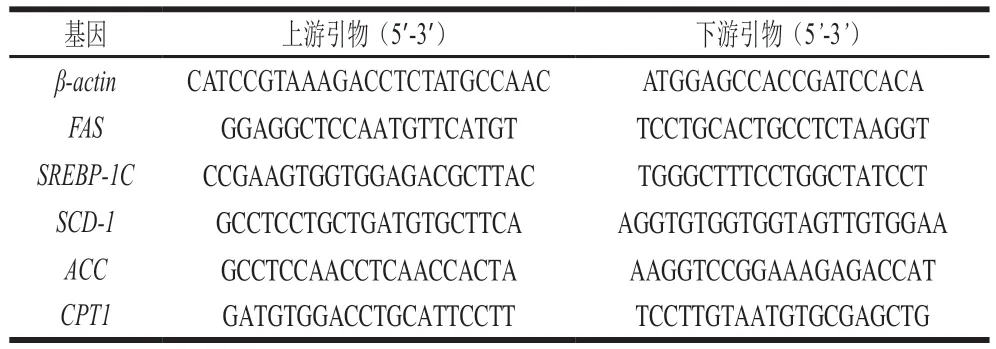
表2 引物序列Table 2 Primer sequences used for PCR amplification
1.3.6 Western blot检测
取200 mg组织,液氮充分研磨,RIPA裂解液提取总蛋白,BCA蛋白浓度测定试剂盒检测蛋白浓度,-80 ℃保存。等量蛋白上样,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)(60 V、60 min;90 V、120 min),100%甲醇激活聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜转膜。质量分数5%脱脂奶粉室温封闭2 h,一抗孵育过夜(1×TBST稀释抗体)、二抗孵育2 h,ECL显色,曝片并拍照,通过ImageJ软件检测蛋白条带的灰度对蛋白表达量进行分析。
1.4 数据处理与分析
采用GraphPad Prism 7软件进行数据统计分析,结果以平均值±标准误差表示。采用单因素方差分析评估各组之间的差异显著性。
2 结果与分析
2.1 YT干预对小鼠体质量及体脂水平的影响
由表3可知,各组小鼠初始体质量无差异;饮食干预12 周后,与LFD组相比,HFD组小鼠体质量高度显著增加(<0.001),说明成功建立饮食诱导的肥胖模型。HFD+2.5% YT组小鼠体质量高度显著低于HFD组(<0.001),说明YT在不影响小鼠膳食习惯的前提下可以促进小鼠减肥;但其摄食量和饮水量的显著增加,说明小鼠体质量的减轻不是因为能量摄入的减少。而HFD+0.5% YT组与HFD组小鼠体质量无显著差异,说明低剂量YT效果不显著,而高剂量的YT具有显著改善效果。
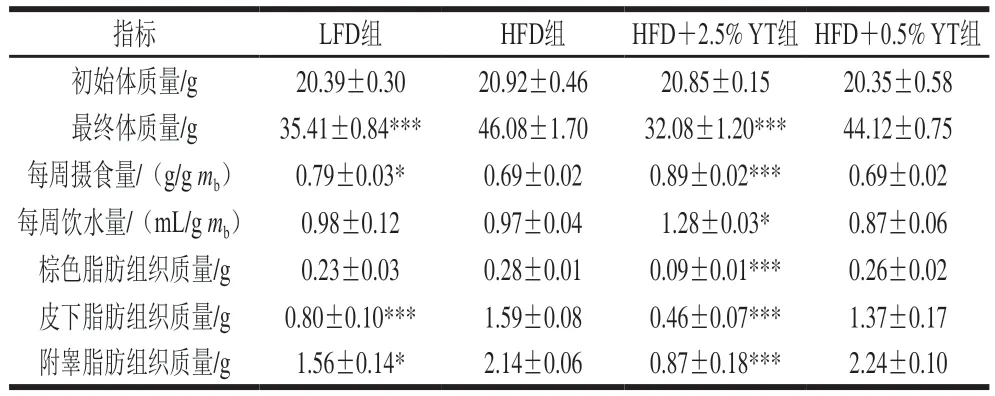
表3 YT对小鼠生理指标的影响Table 3 Effect of YT on physiological indexes of mice
观察喂养12 周的小鼠表型(图1),HFD+2.5% YT组相比HFD组较瘦,与LFD组体型相似,说明高剂量YT饮食干预能明显抵抗高脂诱导的肥胖。同时前期数据显示,HFD组小鼠血清总胆固醇、低密度脂蛋白胆固醇、甘油三酯水平显著升高,而2.5% YT、0.5% YT处理能不同程度降低血脂水平,表明YT具有降低血脂的作用。
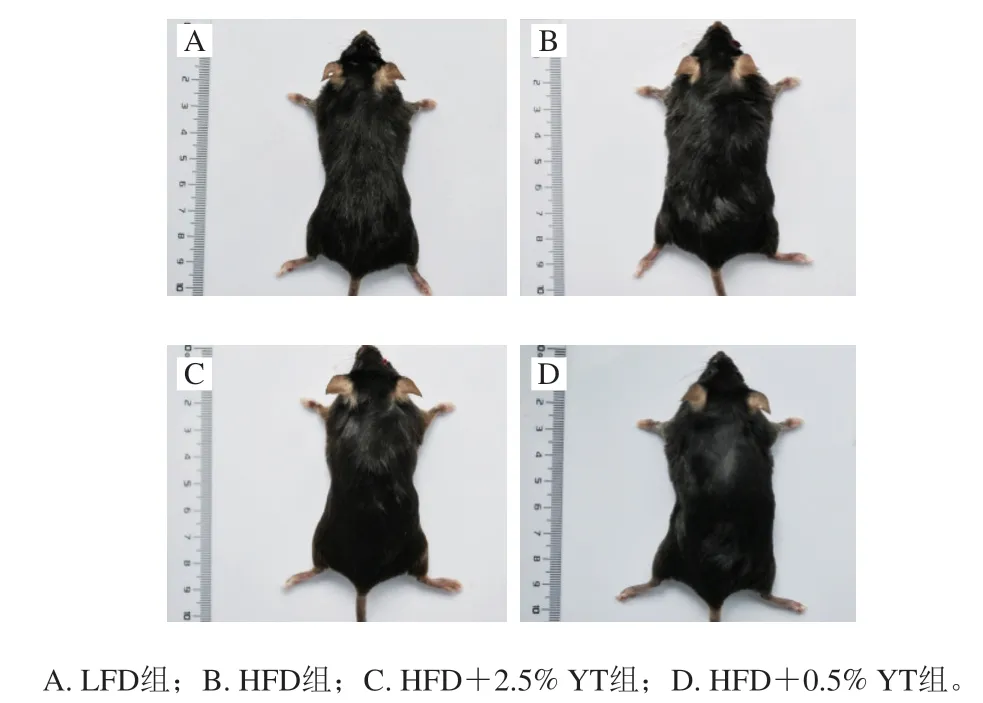
图1 YT干预12 周后的小鼠形态Fig. 1 Morphology of mice after 12-week YT treatment
对各组小鼠3 种典型的脂肪组织质量进行分析,发现HFD组比LFD组小鼠的皮下脂肪(<0.001)、附睾脂肪组织(<0.05)质量显著增加,棕色脂肪组织质量也有上升的趋势,但未达到显著水平(表3)。与HFD组相比,HFD+2.5% YT组3 种脂肪组织质量均高度显著降低(<0.001)(表3),表明YT能有效降低小鼠的体脂水平。
2.2 YT干预对脂肪组织脂质沉积的影响
脂肪组织主要由脂肪细胞组成,而脂肪细胞包括传统意义上的棕色脂肪细胞和白色脂肪细胞,以及近期发现并被关注的浅棕色脂肪细胞。为分析YT对组织中脂质沉积的影响,取小鼠代表性棕色和白色脂肪垫进行组织学分析。如图2A所示,棕色脂肪组织主要由棕色脂肪细胞组成,其细胞具有多腔室结构,内含大量小脂滴,是脂质氧化产生热量的主要场所。HFD诱导后,苏木精-伊红染色结果显示小鼠棕色脂肪细胞由多腔室转变为含有巨大脂滴的单室白色脂肪细胞,说明大量白色脂肪细胞侵入组织;而YT干预明显恢复了其多腔室结构的组织形态,说明其阻止了高脂饮食诱导的棕色脂肪“白色化”(图2A)。
白色脂肪组织由单腔室的白色脂肪细胞组成,每个细胞内仅含有单个大脂滴,是主要负责储存脂肪的细胞。皮下脂肪组织和附睾脂肪组织是两种典型的白色脂肪组织。由图2B可知,HFD相比LFD组小鼠,皮下和附睾脂肪组织细胞面积高度显著增加(<0.001),苏木精-伊红染色分析结果显示细胞内脂滴变大,说明脂质沉积明显增加。而0.5% YT、2.5% YT处理均能减小白色脂肪(皮下脂肪和附睾脂肪)细胞的面积,阻止白色脂肪细胞扩张,减少白色脂肪细胞内脂质积累,进而阻止了脂肪组织膨胀及肥胖的发展进程。
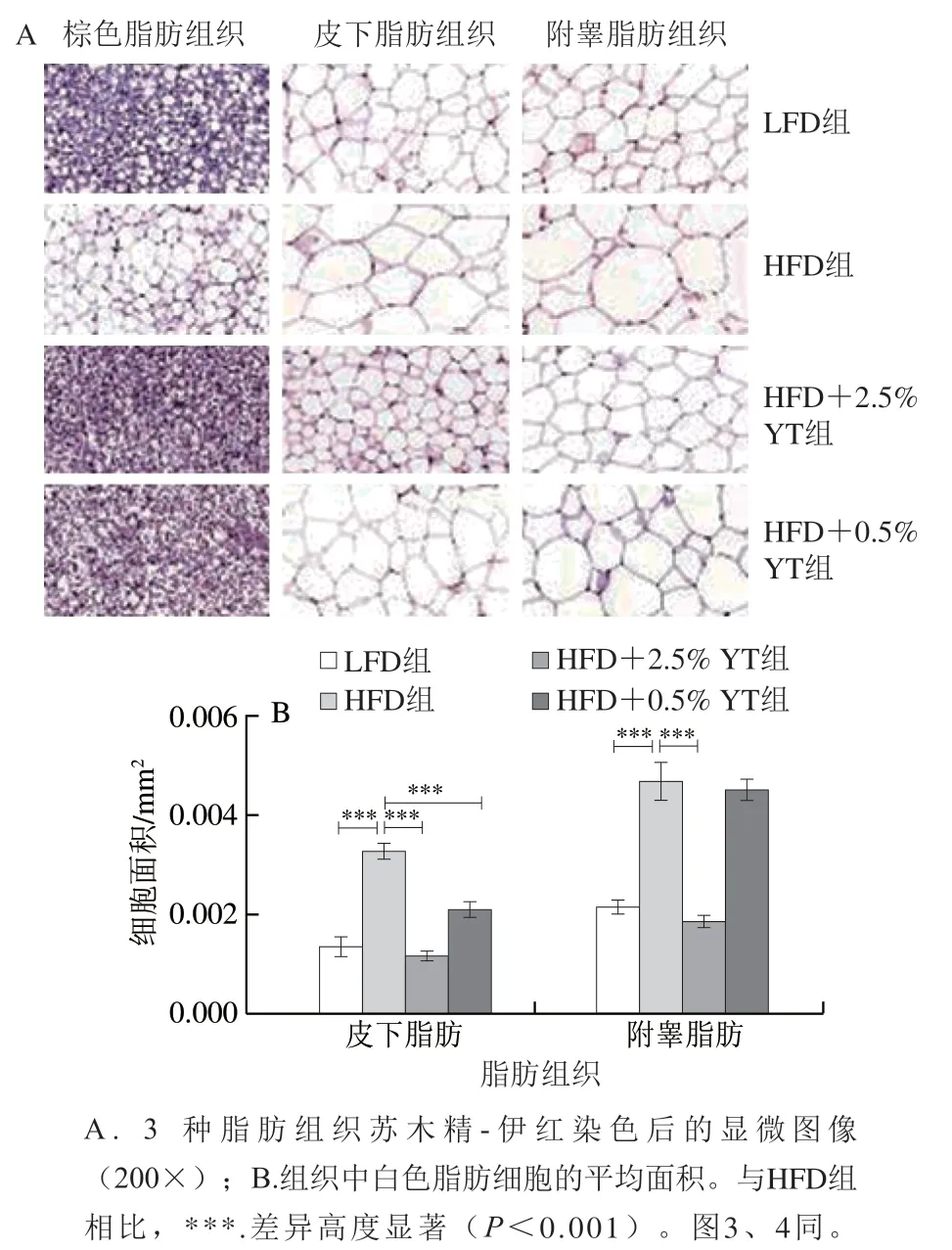
图2 小鼠脂肪组织的形态学分析Fig. 2 Morphological analysis of adipose tissues in mice
2.3 YT干预对小鼠脂肪组织脂肪酸代谢的影响
为进一步探究YT对高脂饮食小鼠脂肪酸代谢的作用,分析对比了各组小鼠脂肪组织脂肪酸代谢相关基因的表达水平。SREBP-1C、FAS、ACC和SCD-1是参与调控脂肪酸生成途径的关键因子。由图3可知,与LFD组相比,HFD组和mRNA表达量在3 种脂肪组织中均呈现下降趋势;而YT干预后,它们的表达水平上升,并呈剂量依赖性。与LFD组相比,HFD组和mRNA表达量也呈下降趋势,且在皮下和附睾脂肪组织中极显著下降(<0.01),而YT干预后,这些脂肪酸生成相关因子的mRNA表达量在3 种脂肪组织中呈不同程度的上升趋势。以上结果提示,YT可能通过上调脂肪酸生成相关基因而调控下游的脂肪酸代谢,进而增加组织储存脂肪的能力;同时的表达量增加有利于维持脂质去饱和,促进脂肪组织的脂肪储存,从而缓解其他组织受到脂质毒性损伤。
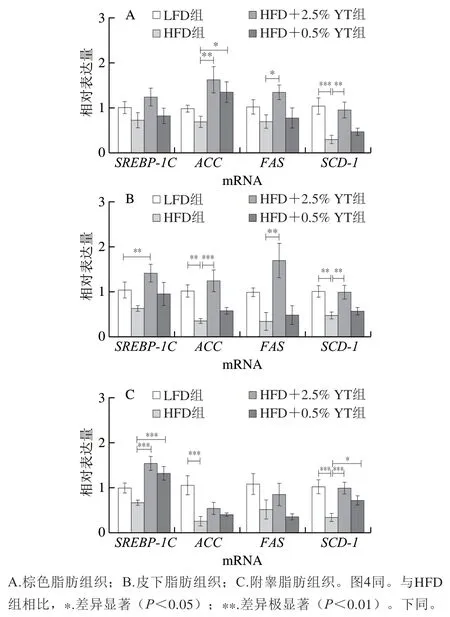
图3 YT对小鼠脂肪组织中脂肪生成相关因子的调控Fig. 3 Effect of YT on the expression of adipogenesis-related genes in adipose tissue of mice
此外,本实验还考察了脂肪酸分解途径的两个关键调控因子和,结果显示,高脂饮食抑制脂肪酸分解途径,而YT高度显著提高和在皮下和附睾脂肪组织中的表达(<0.001),但在棕色脂肪组织中的上调未达显著性水平(图4)。说明YT同时也增强了脂肪组织中的脂肪分解途径,可抵抗和改善高脂饮食导致的脂代谢紊乱;其中YT在棕色脂肪组织中作用微弱,是因为棕色脂肪组织并非负责脂质的存储而是主要负责机体产热代谢。

图4 YT对小鼠脂肪组织脂肪分解相关因子的调控Fig. 4 Effect of YT on the expression of lipolysis-related genes in adipose tissue of mice
2.4 YT干预对脂肪组织AMPK/ACC信号通路的影响
AMPK被称为“细胞能量调节器”,AMPK信号通路是调节细胞能量状态的中心环节。ACC是脂肪酸代谢的限速酶,参与脂肪酸的氧化及合成。在机体应激时,AMPK发生磷酸化并激活下游靶分子ACC,通过增强脂肪酸的β-氧化,减少脂肪酸的合成而改善脂质代谢。大量研究报道,茶叶以协同的方式抑制糖异生和脂肪生成,并通过激活AMPK促进脂肪酸分解代谢,而儿茶素EGCG通过激活AMPK调控葡萄糖生成与分解、脂肪酸合成与分解代谢。因此考察在脂肪组织中YT是否影响AMPK/ACC信号通路,相关蛋白表达情况如图5~7所示。
HFD组小鼠相比于LFD组小鼠,AMPK蛋白的磷酸化水平在皮下脂肪组织中极显著下调(<0.01)(图6C),而在棕色脂肪组织和附睾脂肪组织中未显著改变(>0.05)(图5C和图7C);YT干预后,与HFD相比,HFD+0.5% YT和HFD+2.5% YT组均一定程度上调棕色脂肪组织和附睾脂肪组织中p-AMPK1α/AMPK1α水平,但在皮下脂肪组织中呈下调趋势。同时HFD组的3 种脂肪组织中p-ACC/ACC水平显著或极显著下降(<0.05、<0.01),HFD+0.5% YT和HFD+2.5% YT组均具有抵抗HFD的作用,并上调p-ACC/ACC水平(图5B、图6B和图7B)。本研究结果表明,YT可能通过促进AMPK、ACC磷酸化,增强脂肪酸氧化途径,从而降低脂肪酸在脂肪组织中的积累。

图5 YT干预下小鼠棕色脂肪组织中AMPK、ACC的磷酸化水平Fig. 5 Effects of YT on phosphorylation of AMPK and ACC in brown adipose tissue
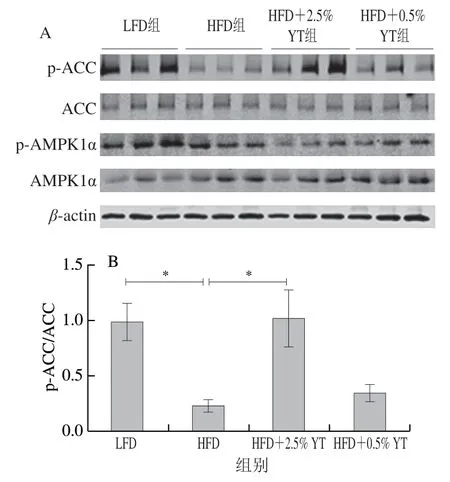

图6 YT干预下小鼠皮下脂肪组织中AMPK、ACC的磷酸化水平Fig. 6 Effect of YT on phosphorylation of AMPK and ACC in subcutaneous adipose tissue
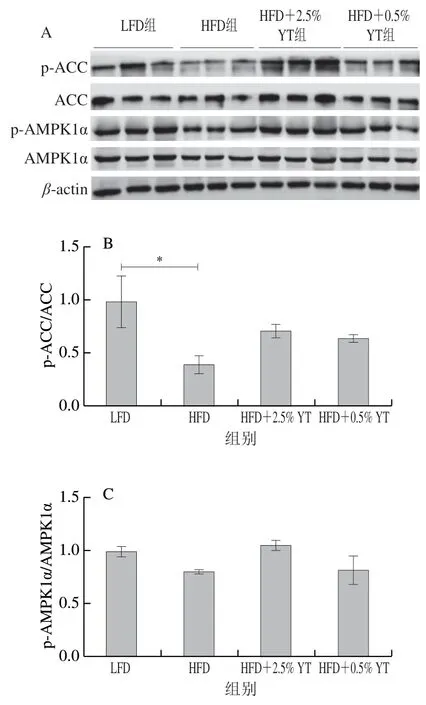
图7 YT干预下小鼠附睾脂肪组织中AMPK、ACC的磷酸化水平Fig. 7 Effect of YT on phosphorylation of AMPK and ACC in epididymal adipose tissue
3 讨 论
在肥胖的发展过程中,过多的脂肪热量超过脂肪组织储存的缓冲能力,导致在肝脏等其他组织中发生“溢出”,即“异位脂肪沉积”。脂肪组织的扩张包括肥大和增生。肥大是指脂肪细胞体积的增长,与脂肪酸和促炎症细胞因子释放量增加、免疫细胞募集程度和脂联素水平降低、胰岛素敏感性受损等有害现象有关;增生是指脂肪细胞数量增长,它被认为是储存健康脂肪的一种机制,与脂联素水平增加、脂肪酸和促炎症细胞因子释放减少、免疫细胞募集程度增加、胰岛素敏感性改善等有益现象有关。与细胞肥大相比,脂肪细胞增生使新分化的脂肪细胞更多地储存脂质,对肥胖相关的代谢并发症起到改善作用。本研究表明,YT干预能有效抑制由高脂饮食导致的小鼠肥胖,控制脂肪组织扩张及脂肪细胞肥大(表3和图1、2),进而有效抵抗小鼠肥胖的发展。
脂肪组织对脂肪酸的储存能力可以防止其他组织产生脂毒性,但肥胖患者脂肪组织功能出现缺陷而损害了其缓冲能力。因此,尽管在早期肥胖中,脂肪酸合成途径在脂肪组织中上调,以快速储存脂肪;但在长期肥胖发展中,相关基因表达量降低,是由于后期的适应性过程目的是限制脂肪组织的进一步膨胀肥大及功能失调。这种逆转的可能解释是,一旦达到脂肪细胞储存能力的界限,在自然抑制性反馈过程之后,细胞就会降低其合成脂肪酸的能力。Soukas等以肥胖症/小鼠模型,研究发现相对于瘦小鼠,肥胖小鼠脂肪组织中的脂肪酸生成关键因子被下调;Moraes等也发现在饮食诱导的肥胖模型中,与脂肪酸代谢相关的基因表达下调。本实验研究结果与以上报道结果相一致,在高脂饮食组,3 种脂肪组织中脂肪生成相关基因、、和表达量均下调;而YT干预整体上显著上调这些基因的表达水平(图3)。因此YT干预能够提高高脂饮食下脂肪组织脂肪酸的储存能力,进而减轻其他组织的脂毒性。
PGC-1α是介导多种细胞能量代谢和核受体的重要因子,它通过激活脂肪酸氧化、糖异生、线粒体呼吸的关键酶参与脂代谢。CPT-1是脂肪酸β氧化的限速酶,负责将脂肪酸转运进线粒体内进行氧化。Chen Liu等比较了6 种茶叶提取物,发现白茶、黄茶和乌龙茶提取物可通过显著上调肥胖小鼠的表达来增加能量消耗和脂肪酸氧化。另有研究报道显示,PGC-1α-CCAAT增强子结合蛋白β(CCAAT/enhancer binding protein β,CEBPB)相互作用正向调节,从而促进脂肪酸氧化。本研究发现,YT干预在两种白色脂肪组织(皮下和附睾脂肪组织)中,显著上调脂肪分解相关基因和的表达水平,进而促进了脂肪组织的脂肪酸氧化;但YT干预在棕色脂肪组织中未达到显著上调效果,这是由组织功能差异决定的,因为棕色脂肪不负责脂质的储存,而是负责机体的产热代谢。
AMPK通路是细胞一级能量平衡的主传感器和调节因子。由ACC催化生成的丙二酰辅酶A不仅可作为脂肪酸合成的前体,也是线粒体摄取脂肪酸所需关键酶CPT-1的内源性抑制剂。细胞内AMPK含量的增加,使ACC磷酸化从而失去活性,导致体内丙二酰辅酶A合成量减少,不仅抑制了脂肪酸的合成,同时也激活了CPT-1,从而促进了脂肪酸氧化作用。在高脂饮食诱导的肥胖小鼠模型中,槲皮素能通过AMPKα1/沉默信息调节因子1(silent information regulator 1,SIRT1)通路抑制脂肪组织巨噬细胞的浸润和炎症反应。熊果酸通过激活肝激酶B1(liver kinase B1,LKB1)/AMPK途径,促进ACC磷酸化和的高表达,进而抑制3T3-L1前脂肪细胞的分化和脂质积累。本研究结果显示,YT促进脂肪组织中的表达,同时促进AMPK和ACC的磷酸化;然而在皮下脂肪组织中仅ACC磷酸化水平增加,AMPK磷酸化水平无明显变化,可能是ACC的磷酸化反向抑制了AMPK1α的活性。
本研究未关注YT健康功效的化学基础。基于大量的研究报道,作为绿茶主要活性成分的EGCG可抑制肝脏糖异生和脂肪生成,并激活AMPK途径促进脂解,故认为EGCG也在YT干预中提供重要的健康效益。在黄大茶加工过程中,EGCG差向异构化形成的GCG,比EGCG在降低胆固醇和三酰甘油水平以及降低餐后血糖等方面有更加显著的效果,提示GCG可能是黄大茶中的另一重要活性物质。此外,黄大茶加工中的焙火工艺产生的一系列儿茶素聚合物,如茶黄素等也可能是另一类有重要贡献的活性物质。在进一步研究中,探索黄大茶主要活性化合物的具体功效是亟待解决的主要问题。
综上所述,YT饮食干预能抵抗高脂饮食诱导的小鼠体质量、脂肪组织质量的增加,并能有效降低脂肪组织中脂质积累量。组织形态分析显示,YT改善了因高脂饮食造成的脂肪细胞肥大及细胞损伤。黄大茶通过增加脂肪生成基因、、、表达而促进脂肪酸的合成,提高脂肪组织的脂肪酸储存能力;同时增加AMPK和ACC磷酸化水平,以及和表达而促进脂肪酸氧化。因此YT能显著改善高脂饮食诱导的脂代谢紊乱,从而减少其他组织的脂毒性损伤。
