不同高粱F2群体的株型相关性状的遗传分析
2022-09-29张旷野王佳旭柯福来张飞朱凯李志华段有厚邹剑秋王艳秋
张旷野,王佳旭,柯福来,张飞,朱凯,李志华,段有厚,邹剑秋,王艳秋
(辽宁省农业科学院 高粱研究所,辽宁 沈阳 110161)
高粱是全球第五大粮食作物,虽然高粱已不再是我国目前的主粮作物,但其仍然是我国农业生产中不可或缺的一部分,在干旱和半干旱地区的农作物生产体系中具有不可替代的优势[1-2]。高粱株型是产量的重要影响因素,同时为了适应机械化、智能化的农业发展方向,目前育种家们已经逐渐从主要关注个体产量性状(诸如高秆、大穗、高生物量)转变为注重群体产量性状(诸如矮秆、整齐度)及相关遗传育种理论研究转变[3]。高粱是典型的C4 作物,产量受群体的光捕获能力和受光姿态的影响,群体的截光能力和受光姿态与光合产物的积累密切相关[4]。高粱株型的改良可以通过调节个体的空间构型,提高群体的光合产物积累,从而提高产量[5]。
高粱的理想株型应该具备中矮杆,分布均匀且上冲的叶片,大而上冲的穗,强壮的根系和茎秆以及适宜的柄伸长度等几方面主要的特征。其目的就是使群体的光合效率和有机物合成水平达到动态平衡,减少群体内竞争而导致不必要的物质损耗,使其更多的形成籽粒产量,并且更加适宜机械化作业,提高经济效益[6-8]。近年来研究者们已经对高粱株型相关性状开展了遗传研究,表1 整理了相关性状的遗传模型研究进展[9-14]。株高方面,董林洁等人认为株高受一对主基因控制[13],而白晓倩和李延玲等的研究都发现株高受两对主基因控制,但是李延玲等认为株高符合加性-显性混合遗传模型,白晓倩等人则认为是株高受两对加性-显性-上位性主基因控制[10-11],逯晓萍等人认为株高遗传受2 对加性-显性主基因+加性-显性多基因控制[12]。叶片数的遗传研究结果也存在一定的差异,白晓倩等、卢华雨等和逯晓萍等认为叶片数主要受两对主基因控制[9,12,14],但是在董林洁的研究中叶片数未检测到受主基因控制[13]。穗长方面,白晓倩等和李延玲等认为穗长符合2MG-EEAD 模型,受2 对主基因控制[9,11]。柄伸长度、穗柄长度和叶面积等性状方面的遗传研究结果较少,且一致性较差。虽然高粱株型相关性状的研究表明,这些性状的遗传机制复杂,因研究材料,方法和环境的不同,结果也存在一定差异,但这些研究同样为育种工作提供了重要的参考依据。
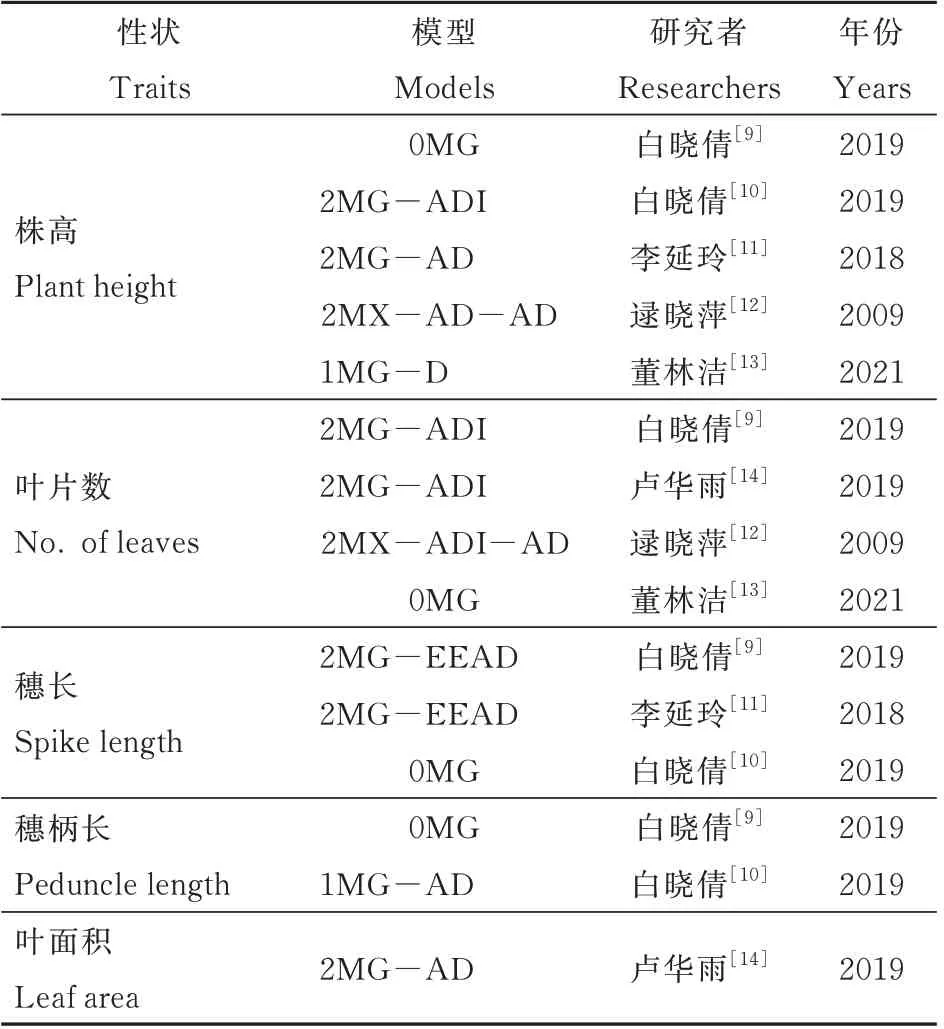
表1 株型相关性状遗传模型研究分析Table 1 Analysis of the genetic models of plant type-related traits
高粱株型是产量形成的关键影响因素之一。目前,高粱品种普遍为叶片较多且肥大平展,且由于株高的降低,节间长度下降,导致叶片分布不合理,容易造成群体郁闭[15-16]。本研究旨在通过分析不同的高粱F2群体叶面积、叶片数、株高、穗长、柄伸长度和穗柄长度的遗传模型,探究各性状的遗传方式,并通过一阶遗传参数和二阶遗传参数估计遗传效应,为高粱的株型改良提供理论依据。
1 材料与方法
1.1 试验材料与种植方式
本研究供试材料为4 个高粱F2分离群体,其中群体1 两亲本为保持系7050B 和Tx2925B,群体内共包含单株255 株;群体2 两亲本为保持系01-26B和7009B,群体内单株数量为217 株;群体3 两亲本为恢复系7037 和矮四,群体样本容量为92 株;群体4 为甜高粱恢复系LTR108 和LR625,群体内包含单株192 株。
试验于2020 年5 月在辽宁省农业科学院试验田(123.56°E,41.82°N)种植8 个亲本,并进行去雄杂交工作,获得F1代种子后。于2020 年10 月于辽宁省农业科学院农作物海南育种中心(108.78°E,18.61°N)F1代种子自交繁种,于2021 年获得F2代群体种子。2021 年5 月于辽宁省农业科学院试验田(123.56°E,41.82°N)种植4 个F2分离群体以及各自亲本,行距0.6 m,株距0.2 m,其中亲本各种植4 行。前茬作物为野生大豆。土壤类型为沙壤土。常规栽培管理。
1.2 表型调查方法
叶面积(Leaf Area,LA)调查方法为于授粉结束后10 天利用直尺测定亲本及F2群体的全株叶长,叶宽(叶片最宽处宽度),通过计算得到叶面积(叶面积=叶宽*叶长*0.75[17])将平均值做为表型数据,其中群体4 未调查全株叶面积。
全株叶片数(Number of leaves,LN)调查方法为分别在五叶期,十叶期,以及抽穗期用红色油漆对第五,第十和第十五片叶进行标记,授粉结束后统计叶片数。
株高(Plant Height,PH)调查方法为利用塔尺测量地表到穗顶的高度。穗长(Spike Length,SL)调查方法为穗顶部至穗基部的长度。柄伸长度(Exsertion Length,EL)指穗基部到旗叶基部的长度。穗柄长度(Peduncle Length,PL)为穗基部到节间长度。穗长,柄伸长度和穗柄长度均使用卷尺进行测量,3 个性状示意图如图1 所示。

图1 穗长,柄伸长度和穗柄长度性状示意图Fig.1 Diagram of spike length,exsertion length and peduncle length
1.3 数据分析方法
利用SPSS24.0 软件对亲本以及F2分离群体进行基本表型分析,包括亲本间的T 检验,群体变异系数、峰度、偏度以及正态分布检验等。遗传模型分析根据植物数量性状遗传体系的分离分析方法,利用章元明等人开发SEA 软件包中SEA-F2功能对不同群体的株型相关性状进行遗传模型分析,参数设置为Threshold=0.0001。在得到所有模型后,对不同模型进行极大对数似然估计,并利用 IECM(Iterated Expectation and Conditional Maximization)法计算得到极大似然函数值(Log_Max_likelihood)和AIC 值(Akaike Information Criterion),从中选择AIC 值最小的3 个模型作为候选模型,之后对3 个候选模型进行合适性检验,包括均匀性检验(U12、U2
2、U32)、Smimov 检验n W2和Kolmogorov 检验Dn,选择显著性差异个数最少的模型做为最适模型,并通过计算得到一阶遗传参数和二阶遗传参数[18-25]。
2 结果与分析
2.1 四个群体亲本差异比较
7050B 和Tx2925B 在叶面积,株高,穗长和柄伸长度四个性状间存在显著性差异。其中7050B的全株平均叶面积,株高和穗长显著大于Tx2925B,而柄伸长度则显著小于Tx2925B。群体2 两亲本01-26B 和7009B 在叶面积,叶片数,株高和穗长方面存在显著性差异,其中01-26B 的叶面积和穗长显著大于7009B,而叶片数和株高则显著小于7009B。7037 和矮四在叶面积,叶片数,株高,穗长和柄伸长度5 个性状间差异明显。其中7037 的叶面积,叶片数和穗长均显著小于矮四,而株高和柄伸长度则显著大于矮四。LTR108 和LR625 只在叶片数和株高方面存在明显差异。LTR108 的叶片数显著大于LR625,而株高则显著小于LR625(表2)。
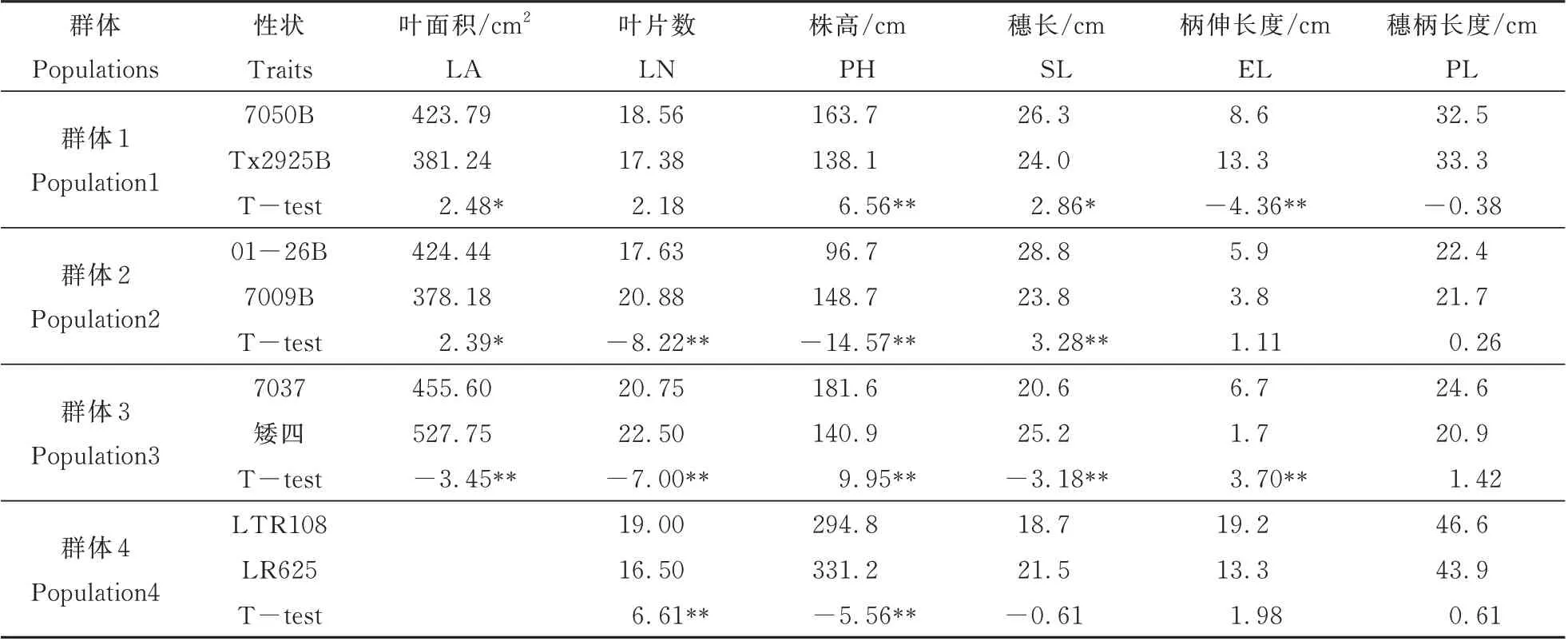
表2 4 个群体的亲本株型相关性状比较Table 2 Comparison of parental plant type related traits of four populations
2.2 F2群体表型分析
4 个群体的叶面积、叶片数、株高、穗长、柄伸长度和穗柄长度的表型分析结果如表3 和图2 所示。群体1 和群体2 的叶面积平均值均大于两亲本,群体3 的叶面积平均值介于两亲本之间,接近于矮四的叶面积,3 个群体的叶面积变异系数为11.88%~15.43%,且均有超亲个体存在,正态分布检验表明3 个群体的叶面积均服从正态分布。在叶片数方面,群体1 和群体4 的叶片数平均值均大于两亲本,群体2 的群体叶片数平均值小于两亲本,群体3 则介于两亲本之间,四个群体的叶片数变异系数较小,均在10%以下,除群体4 中没有超低亲个体的存在,其他群体的叶片数中均存在超亲个体,正态分布检验结果表明4 个群体的叶片数均不符合正态分布。4 个群体的株高平均值均介于各自亲本之间,变异系数在15.39%~31.67%之间,且均有超亲个体存在,群体内变异丰富,其中群体2 和群体3 的株高符合正态分布,群体1 和群体4 则不符合正态分布。4 个群体的穗长平均值除群体2 小于两亲本之外,其余均介于各自两亲本之间,4 个群体内株高变异丰富,均存在超亲个体,夏皮洛-威尔克检验结果表明,群体3 和群体4 符合正态分布,而群体1 和群体2 则不符合正态分布。4 个群体的柄伸长度平均值均介于各自亲本之间,柄伸长度的变异系数较大,均在60%以上,同样存在超亲个体,且均不符合正态分布。4 个群体中,群体2 和群体3 的穗柄长度平均值均大于各自亲本,群体4 则小于亲本,群体1 介于两亲本之间,4 个群体的穗柄长度变异系数为23.04%~35.59%,群体内变异丰富,均存在超亲个体,正态分布检验结果表明,群体3 和群体4 符合正态分布,其余两群体的穗柄长度则不符合正态分布。
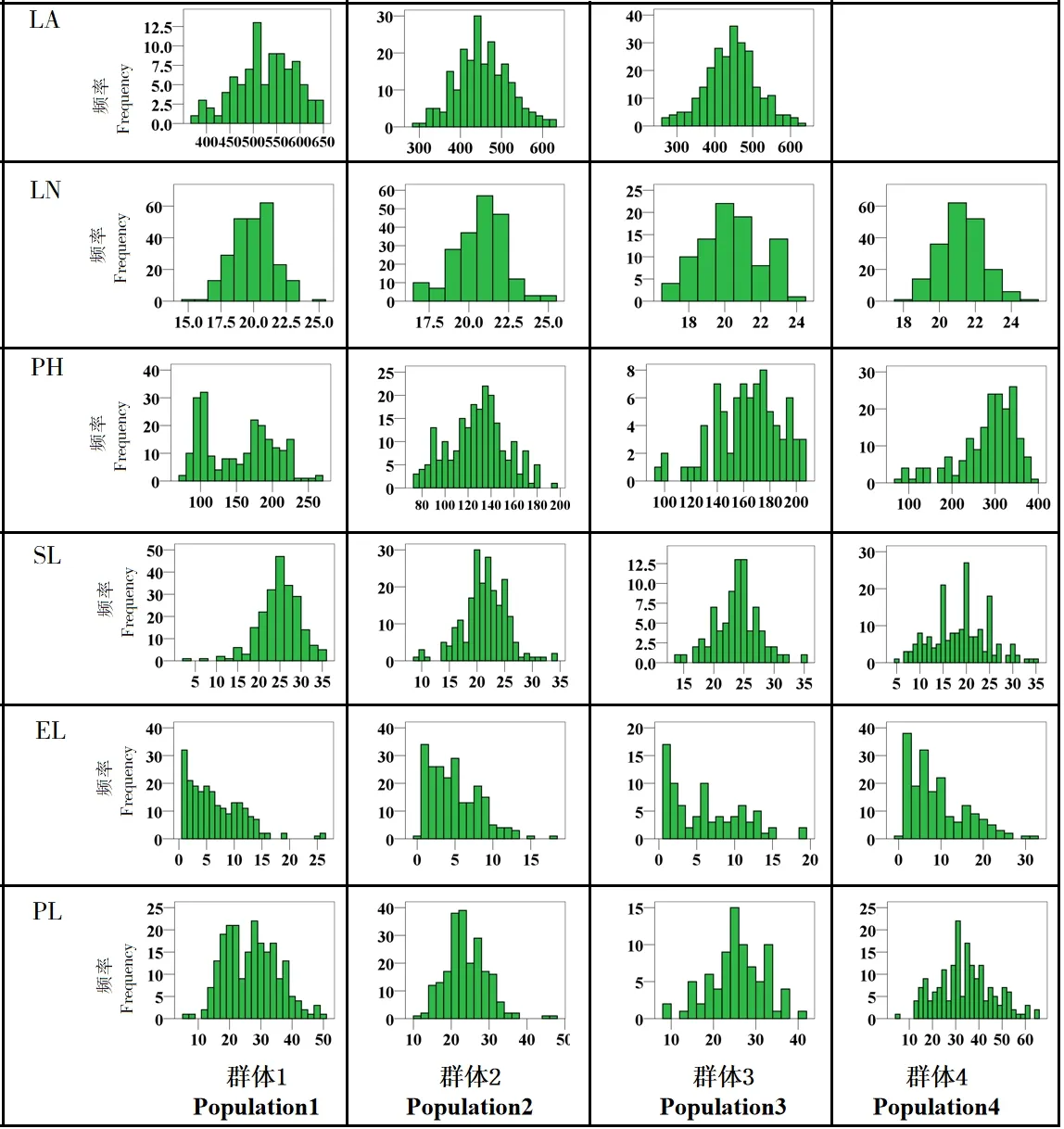
图2 株型相关性状频率分布直方图Fig.2 Frequency distribution histogram of plant type related traits
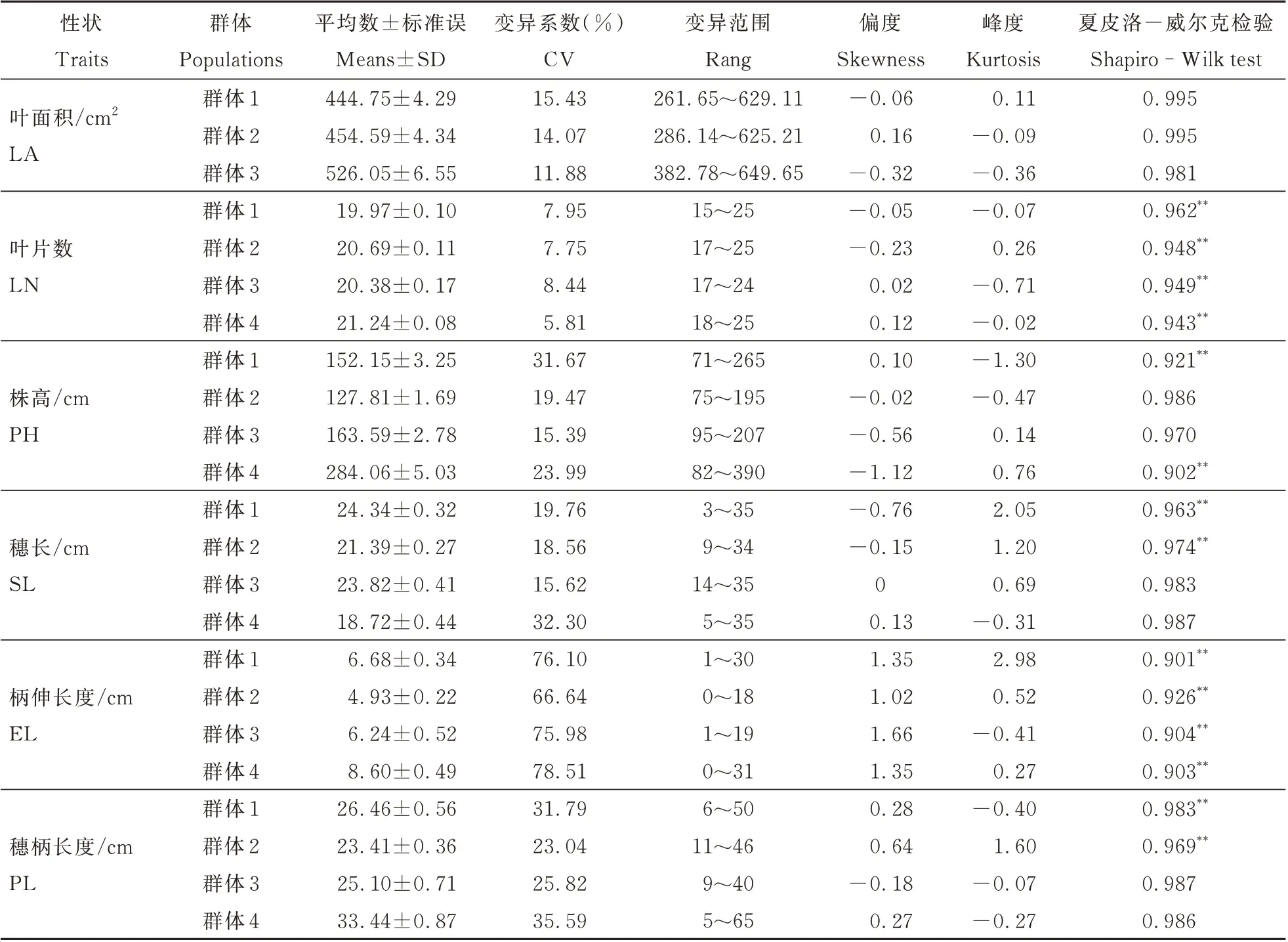
表3 四个群体株型相关性状的表型分析Table 3 Phenotypic analysis of plant type-related traits in four populations
2.3 遗传模型分析
2.3.1 最适遗传模型筛选
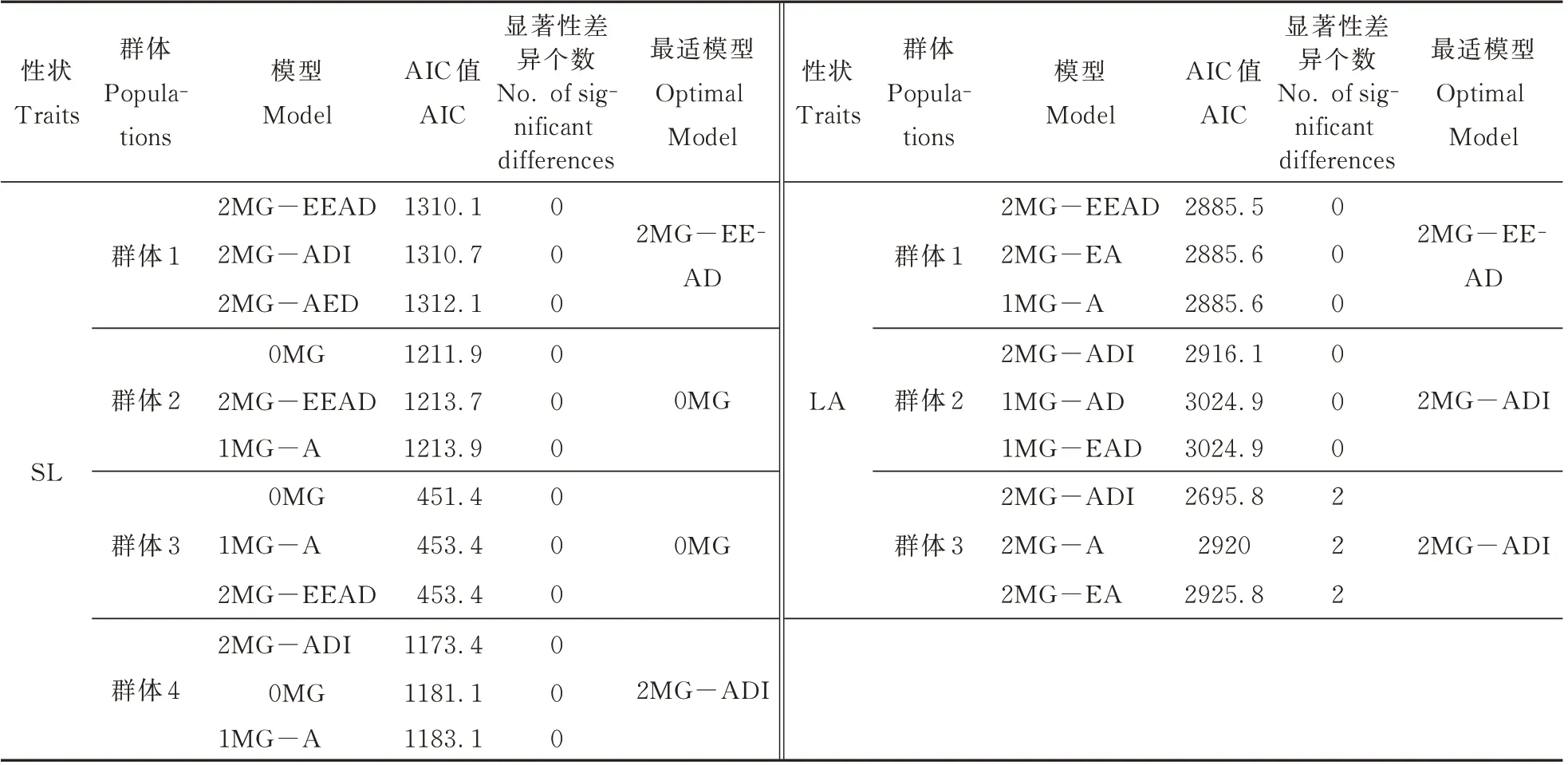
根据筛选规则筛选出各群体各性状的最适遗传模型,筛选结果如附表1 所示。群体1、群体2 和群体3 叶面积最适遗传模型分别为2MG-EEAD(两对等加性-等显性主基因)、2MG-ADI(两对加性-显性-上位性主基因遗传模型)和2MG-ADI,即3 个群体的叶面积都是受2 对主基因控制。4 个群体的叶片数均符合2MG-ADI 模型。4 个群体的株高最适遗传模型分别为1MG-AD(一对加性-显性主基因遗传模型)、2MG-A(两对加性基因)、2MG-EEAD 和2MG-ADI,其中群体1 株高受1 对主基因控制,而其余3 个群体的株高受2 对主基因控制。群体2 和群体3 中的穗长并未检测到主基因存在,主要受微效基因影响,群体1 和群体4 的穗长则分别符合2MG-EEAD 和2MG-ADI 遗传模型。群体2 和群体4 的柄伸长度均符合2MGAD,即两对加性-显性主基因遗传模型,而群体1和群体3 则符合2MG-ADI 模型。4 个群体的穗柄长度最适模型均为0MG,即均未检测到主基因存在。

附表1 最适模型筛选Appendix 1 Screening of the optimal model
续表
2.3.2 遗传参数分析
4 个群体的株型相关性状最适模型的遗传参数如表4 所示(群体2 和群体3 的穗长以及4 个群体的穗柄长度均为0MG,故没有列出)。

表4 4 个群体株型相关性状的遗传参数分析Table 4 Genetic parameters analysis of plant type related traits in four populations
(1) 叶面积遗传参数分析
群体1 的叶面积最适模型为两对等加性-等显性主基因,所以两对主基因的加性和显性效应均为12.33,表现为正向完全显性,主基因遗传率较小,仅为5.24%;群体2 的叶面积两对主基因的加性效应(180.7 和116.25)均为正向效应,显性效应则表现为一正一负(-15.02 和64.65),第1 对主基因表现为负向部分显性,显性度(ha/da)为-0.08,第2 对主基因为正向部分显性(显性度=0.56),因为2 对主基因的加性效应(|da|+|db|),显性效应(|ha|+|hb|)和上位性效应(|i|+|jab|+|jba|+|l|)分别为296.95,79.67 和185.27,所以受加性效应影响较大;群体3 的叶面积两对主基因的加性效应(253.15 和41.63)均为正向效应,显性效应同样表现为一正一负(-379.73 和4.61),第1 对主基因表现为负向超显性(显性度=-1.50),第2 对主基因为正向部分显性(显性度=0.11),因为两对主基因的加性,显性和上位性分别为294.78、384.34 和253.16,所以受显性效应影响较大。群体2 和群体3 的主基因遗传率较大分别为99.56%和99.55%。
(2) 叶片数遗传参数分析
群体1 的叶片数两对主基因加性(6.27 和5.15)和显性效应(1.13 和3.6)均表现为正向效应,根据显性度来看,两对主基因均为正向部分显性(0.18 和0.70),加性效应起主要作用(加性效应11.42>上位性效应10.28>显性效应4.73);群体2 两对主基因的加性效应(7.27 和4.98)均为正向,第1 对主基因的显性效应为正向(0.39),表现为正向部分显性(显性度=0.05),另一对的显性效应为负向(-4.48),表现为负向部分显性(显性度=-0.90),2对主基因以上位性效应为主(上位性效应16.41>加性效应12.25>显性效应4.87);群体3 两对主基因加性效应均为正向(10.26 和1.08),显性效应均为负向(-15.39 和-0.03),第1 对主基因表现为负向超显性(显性度=-1.50),第2对主基因表现为负向部分显性(显性度=-0.03),显性效应起到较大作用(显性效应15.42>加性效应11.34>上位性效应10.32);群体4 两对主基因的加性效应(6.80 和5.27)均为正向,第1 对主基因的显性效应为正向(1.33),表现为正向部分显性,显性度=0.20,另一对的显性效应为负向(-4.77),表现为负向部分显性,显性度=-0.91),2 对主基因以上位性效应为主(上位性效应17.94>加性效应12.07>显性效应6.1)。4 个群体的叶面积主基因遗传率均在95%以上。
(3) 株高遗传参数分析
群体1 的株高符合一对加性-显性主基因遗传模型,主基因加性效应(56.71)和显性效应(15.51)均为正向,表现为正向部分显性(显性度=0.27),主基因遗传率为89.42%;群体2 的株高符合两对加性主基因遗传模型,两对主基因的加性效应均为正向(10.39 和35.53),主基因遗传率为91.77%;群体3 的株高最适模型为两对等加性-等显性主基因,所以两对主基因的加性和显性效应均为16.67,表现为正向完全显性,主基因遗传率为66.99%;群体4 两对主基因的加性效应(84.63 和31.65)均为正向,第1 对主基因的显性效应为负向(-16.86),表现为负向部分显性(显性度=-0.20),另一对的显性效应为正向(6.30),表现为正向部分显性(显性度=0.20),2 对主基因以上位性效应为主(上位性效应123.74>加性效应116.28>显性效应23.16),主基因遗传率为94.01%。
(4) 穗长遗传参数分析
群体1 的穗长最适模型为两对等加性-等显性主基因,所以2 对主基因的加性和显性效应均为2.46,表现为正向完全显性,主基因遗传率为39.02%;群体4 穗长两对主基因的加性效应(8.14和3.32)均为正向,显性效应均为负向(-6.01 和-1.92),均表现为负向部分显性(显性度分别为-0.74 和-0.58),两对主基因以上位性效应为主(上位性效应12.59>加性效应11.46>显性效应7.93),主基因遗传率为94.01%。
(5) 柄伸长度遗传参数分析
群体1 柄伸长度两对主基因的加性效应(7.20和3.29)均为正向,显性效应均为负向(-7.48 和-3.26),其中第1 对主基因表现为负向超显性(显性度=-1.04),第2 对主基因表现为负向部分显性(显性度=-0.99),两对主基因以上位性效应为主(上位性效应13.72>显性效应10.74>加性效应10.49),主基因遗传率为79.44%;群体2 两对主基因的加性效应(2.66 和2.36)均为正向,显性效应(-3.74 和-1.47)均为负向,其中第1 对主基因表现为负向超显性(显性度=-1.41),第2 对主基因表现为负向部分显性(显性度=-0.62),主基因遗传率为76.89%;群体3 柄伸长度两对主基因的加性效应(6.66 和1.98)均为正向,显性效应均为负向(-6.24 和-0.89),2对主基因均表现为负向部分显性(显性度分别为-0.94和-0.45),2对主基因以上位性效应为主(上位性效应12.62>加性效应8.64>显性效应7.13),主基因遗传率为96.39%;群体4 两对主基因的加性效应(6.29 和4.34)均为正向,显性效应(-8.86 和-2.24)均为负向,其中第1 对主基因表现为负向超显性(显性度=-1.41),第2 对主基因表现为负向部分显性(显性度=-0.52),主基因遗传率为87.44%。
3 讨论
本试验以4 个不同高粱F2分离群体为试材,对叶面积,叶片数,株高、穗长、柄伸长度和穗柄长度等多个株型相关性状进行田间调查,并分析遗传规律,发现这些性状均为典型的数量性状,且受两对主基因遗传的性状较多,包括叶面积,叶片数,柄伸长度以及部分群体的株高和穗长,为后续的基因定位和分子标记辅助选择育种奠定了理论基础。
本研究结果表明高粱株型性状属于复杂的数量性状。其中4 个群体的叶片数均受两对加性-显性-上位性主基因控制,与白晓倩和卢华雨等的研究结果相吻合[9,14],且本研究中4 个群体的叶片数的主基因遗传率均在96%以上,说明高粱叶片数受环境影响较小,可以在早代对叶片数进行改良。株高方面,董林洁的研究中发现株高主要受一对主基因调控[13],与本研究中群体1 的结果相符。而在白晓倩,李延玲等,逯晓萍等的研究中株高受两对主基因调控,但是主基因的作用方式略有不同[10-12],这与本研究中群体2、3 和4 的分析结果相吻合。在株高的遗传率方面,虽然群体3 的遗传率为66.99%,但其余3 个群体的遗传率较高,在育种工作中同样可以考虑对株高进行早代选择。穗长方面,群体2 和3 中未检测到主基因存在,与白晓倩等的研究结果相吻合[10],群体1 符合2MG-EEAD 模型,与白晓倩等,李延玲等的研究结果相吻合[9,11],群体1 和群体4 的穗长遗传率相差较大,同时考虑到有主基因不存在的情况,因此在亲本血缘关系不明确的情况下,不建议对穗长进行早代选择。与之类似的还有叶面积,根据3 个群体叶面积的遗传率,同样不建议对其进行早代选择。穗柄长方面,本研究中的4 个群体中均为检测到主基因的存在,与白晓倩等的研究结果相吻合[9],但是卢华雨等的研究结果存出入[26],造成这种情况的原因可能是所用群体亲本的遗传背景存在差异、试验群体大不同以及种植环境差异较大等,仍需要更深入的研究与探讨。而柄伸长度方面的遗传研究,国内外尚未见报道,在本研究中,柄伸长度均受两对主基因调控,而且遗传率均比较高,因此可以对柄伸长度进行早代选择。
本研究以4 个不同的F2群体做为试材,对六个株型相关性状进行了遗传分析。虽然因为高粱F1代种子获取难度较大,没有获得有效的F1代各性状的表型数据,导致本研究中无法对多基因的遗传效应进行分析,但是本研究明确了各株型相关性状的主基因遗传方式和遗传率的大小,为高粱的理想株型育种提供了重要的理论依据和指导建议。
4 结论
通过对四个高粱F2分离群体的叶面积,叶片数,株高、穗长、柄伸长度和穗柄长度等性状的遗传模型分析,发现7050B×Tx2925B 群体、01-26B×7009B 群体和7037×矮四群体叶面积,4 个群体的叶片数和柄伸长度,01-26B×7009B 群体,7037×矮四群体和LTR108×LR625 群体株高,7050B×Tx2925B 群 体 和LTR108×LR625 群 体的穗长均受两对主基因控制。7050B×Tx2925B群体的株高则受1 对主基因控制,01-26B×7009B群体和7037×矮四群体的穗长和四个群体的穗柄长度中均未检测到主基因的存在。叶片数,株高和柄伸长度等性状的主基因遗传率较大,可以在早代进行改良和选择。
