褐飞虱两个糖转运蛋白的功能分析及调控海藻糖代谢的影响
2022-09-22罗雨嘉邱玲玉刘永康庞晓青王世贵徐彩娣
罗雨嘉,邱玲玉,刘永康,庞晓青,姚 琼,王世贵,唐 斌,徐彩娣,3*
(1.杭州师范大学生命与环境科学学院,杭州 311121;2.广东省农业科学院植物保护研究所/广东省植物保护新技术重点试验室,广州 510640;3.杭州师范大学经亨颐教育学院,杭州 311121)
水稻OryzasativaL.作为我国的主要粮食作物几乎每年消费产量达2万吨以上,根据报道估计,在各种水稻病虫害中仅因褐飞虱产生的损失就占四成以上(徐春春等,2018;刘杰等,2021),这严重危害我国粮食的生产与安全。褐飞虱Nilaparvatalugens单食水稻,其刺吸式口器直接插入水稻维管束的韧皮部中吸取汁液,汁液中含有大量糖分,这是褐飞虱能量物质的来源(Sōgawa,1982)。有研究表明,褐飞虱在吸食水稻的过程中不能直接利用水稻内的蔗糖,而是通过糖转运蛋白介导蔗糖进入飞虱体内(Priceetal.,2007;Kikutaetal.,2015),但具体机制尚未明确。
海藻糖是一种非还原性二糖,广泛存在于许多原核生物和真核生物中,包括细菌、真菌、昆虫、植物等(Elbeinetal.,2003;Lunnetal.,2014),它可以作为能量和糖的来源(Benaroudjetal.,2001;Boninietal.,2004;Lunnetal.,2014),在昆虫中被称为“昆虫血糖”,是大多数昆虫主要的血淋巴糖(Thompson,2003;Avonceetal.,2006)。与存在海藻糖的许多生物一样,昆虫的海藻糖也是通过TPP/TPS途径合成,即在昆虫脂肪体中海藻糖的合成依赖于海藻糖合成酶(Trehalose-6-phosphate synthase,简称TPS)和海藻糖磷酸化酶(Trehalose-6-phophate phosphatase,简称TPP)的催化,并且受唯一的水解酶——海藻糖酶(Trehalase,简称TRE)的作用水解为两分子的葡萄糖(Beckeretal.,1996;Shuklaetal.,2015),最后通过特定的跨膜转运蛋白运输至血淋巴中,由此维稳着体内碳水化合物代谢的平衡(Leyriaetal.,2021)。正如Leyria所描述,海藻糖与葡萄糖一样也不能直接穿透细胞膜(Leyriaetal.,2021),生命体对于这些糖类物质的利用都必须经过一个膜转运的过程,其中一种方式就是需要借助糖转运蛋白(Sugar transporter,简称ST)的运输进出细胞(Chenetal.,2015)。
糖转运蛋白是血糖水平动态平衡的关键,例如哺乳动物受激素调节通过葡萄糖转运蛋白(Glucose transporter,简称GLUT)GLUT4维持血糖水平,由胰岛素信号通路发出信号控制GLUT4易位至细胞膜,在细胞膜上将血液中多余的葡萄糖运输进细胞(Huang and Czech,2007);而在植物中SWEET转运蛋白已被确定为促进蔗糖在多种组织中扩散到质体中的蛋白,Durand等人研究发现SWEET基因表达量的上调可能会使蔗糖到胞外基质的外排增加,从而适应干旱环境(Leetal.,2015;Durandetal.,2016;Juliusetal.,2017)。此外,昆虫的海藻糖转运蛋白(Trehalose transporter,简称TRET)能够协助新合成的海藻糖从脂肪体内转移到循环的血淋巴中,被其他组织吸收,并且根据海藻糖的浓度梯度和生理需要以可逆的双向方式起作用(Thompson,2003;Leyriaetal.,2021),细胞内海藻糖的水平与昆虫抵抗各种环境胁迫存在相关性(Benoitetal.,2009;Cornetteetal.,2010)。由此可见,糖转运蛋白的协助对于糖代谢的正常运作来说必不可少,它严密调控着血糖浓度从而维持其生命活动,在昆虫中海藻糖转运蛋白则是由执行该功能的糖转运蛋白之一。
综上,糖转运蛋白可为控制害虫提供新的潜在目标。但是,目前关于昆虫糖转运蛋白特别是海藻糖转运蛋白的功能研究和信息相对较少,仅少数学者明确研究过昆虫海藻糖转运蛋白的功能(Kikawadaetal.,2007;Kanamorietal.,2010;Liuetal.,2013;Kikutaetal.,2015),且已有的研究大多数集中在海藻糖的合成和分解途径(Shuklaetal.,2015;Tangetal.,2018)。随着全基因组测序的完成,褐飞虱逐渐成为研究基因功能的靶标昆虫。因此,阐明昆虫血淋巴中糖的摄取机制,了解能量物质的获取途径,从而可确定控制害虫的新目标。本研究通过对褐飞虱两个糖转运蛋白基因进行功能分析,进一步利用RNAi技术探究其调控海藻糖代谢的生理作用,鉴定该基因是否为海藻糖转运蛋白候选基因,从而评估其作为控制害虫潜在基因的可能性,为将来寻找防治害虫新靶标提供理论依据。
1 材料与方法
1.1 褐飞虱糖转运蛋白基因的生物信息学分析
从美国国际生物技术信息中心NCBI数据库(https://www.ncbi.nlm.nih.gov/)获得褐飞虱两条糖转运蛋白的基因序列,以FASTA格式保存。使用NCBI上自带的Blast-N和Blast-X工具将这两条基因的cDNA序列与GenBank中的其他糖转运蛋白的核酸序列以及氨基酸序列进行比较。通过软件Clustal X对这两个蛋白的氨基酸序列与其他昆虫糖转运蛋白氨基酸基因序列进行多序列比对,并用Mega 5.1中的邻接法构建系统发育树,设置Bootstrap法的重复值为1 000(Henaoetal.,2014)。
利用瑞士生物信息研究所(SIB)维护的蛋白组学分析平台ExPASy(https://web.expasy.org/protparam/)预测本研究所使用的两条基因序列翻译为蛋白后所表现的物理化学性质,这些性质包括相对分子质量(Molecular weight)、理论等电点(Theoretical pI)、原子组成(Atomic composition)不稳定指数(Instability index)等。然后利用在线预测工具SMART(http://smart.embl-heidelberg.de/)预测蛋白结构域的构成和功能,使用NCBI Conserved Domains(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)对蛋白质的保守结构域进行预测。对于蛋白质二级结构的预测使用在线预测工具PSIPRED(http://bioinf.cs.ucl.ac.uk/psipred/),再将氨基酸序列提交到SWISS-MODEL网站(https://swissmodel.expasy.org/interactive)进行蛋白质三级结构建模。最后利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)对该蛋白进行亚细胞定位。
1.2 供试昆虫
供试昆虫褐飞虱为杭州师范大学动物适应与进化重点实验室饲养种群,供试水稻为感虫水稻TN1(Taichung Native1),褐飞虱置于人工气候室饲养,环境条件设置为:温度25±1℃,光周期L ∶D=16 h ∶8 h,相对湿度70%±5%。
1.3 总RNA的抽提与dsRNA的合成
采用Trizol法提取褐飞虱总RNA,并用1%琼脂糖凝胶电泳和Nano Drop2000分光光度计分别检测所提取总RNA的质量以及浓度。根据Prime Script©RT reagent Kit With g DNA Eraser试剂盒的说明书,合成cDNA第一链,并放于-20℃冰箱保存备用。采用Primer5软件设计合成特异性引物进行PCR扩增,产物经过切胶回收与纯化后连接于p MD 18-T载体进行T克隆,挑取单一菌落进行菌落PCR,选取阳性菌液送往Invitrogen(上海)公司测序。以返回的质粒为模板使用含T7启动子的引物进行交叉PCR,并按照T7 Ribo MAXTMExpress RNAi System试剂盒的使用方法进行双链RNA(dsRNA)的合成。采用相同的方法合成绿色荧光蛋白基因(GFP)的dsRNA作为对照组,相关引物序列具体见表1。使用Nano Drop2000分光光度计检测dsRNA的浓度,并加入DEPC处理水将其稀释为终浓度50 μg/μL,最后放入-80℃冰箱保存备用。

表1 dsRNA合成引物及荧光定量PCR引物序列Table 1 Primers sequences used for dsRNA synthesis and fluorogenic quantitative PCR
1.4 褐飞虱的显微注射
选取发育至5龄第1天的褐飞虱若虫用于RNAi显微注射试验。
首先将dsRNA溶液导入玻璃毛细管针中,设定显微注射仪参数为:注射压强1 000 pah,补偿压80 pah,并根据注射入标准毛细管的体积(5龄褐飞虱的注射体积为100 nL/头)调整注射时间。然后用毛笔挑取CO2麻醉后的褐飞虱使其腹面朝上,放置在提前准备好的琼脂糖胶台的凹槽中,在莱卡EZ4解剖镜的观察下将针刺入褐飞虱胸部第二和第三对足之间并注入dsRNA。把处理后的飞虱放入含新鲜水稻苗的玻璃管中,48 h后挑取存活的褐飞虱用于荧光定量分析、糖含量检测以及海藻糖酶活性测定。挑取3个平行的样品,每个平行10头褐飞虱用于检测RNAi后基因表达水平;挑取3个平行的样品,每个平行20头褐飞虱用于测定海藻糖酶活和糖含量。
1.5 RNAi后褐飞虱体内海藻糖代谢通路相关基因表达量测定
取不同处理组显微注射48 h后的褐飞虱抽提总RNA,参照试剂盒反转录得到cDNA,并根据SYBR©Premix Ex TaqTM试剂盒的方法进行qRT-PCR检测,具体定量引物见表1。荧光定量PCR程序:95℃预变性10 s,95℃解链5 s,59℃退火并延伸30 s,40个循环。每个处理3个模板,每个模板重复3次。目的基因的相对拷贝数根据2-ΔΔCT方法进行分析(Livaketal.,2001)。
1.6 总糖原、海藻糖、葡萄糖含量以及海藻糖酶活性测定
取注射后48 h的材料分为对照组与实验组,向其加入200 μL PBS,经过组织破碎仪(程序:55 Hz,120 s)充分研磨后放入超声破碎仪进行超声破碎0.5 h,往破碎后的样品再加入800 μL PBS补足1 mL,4℃,1 000 g离心20 min。取2份350 μL上清,1份用于总蛋白、总糖原和海藻糖含量的测定;1份进行4℃,20 800 g超离心1 h,超离心后取300 μL上清液用于测定葡萄糖、蛋白质含量以及可溶型海藻糖酶活性,沉淀于300 μL的PBS悬浮后用于测定葡萄糖、蛋白质含量和膜结合型海藻糖酶活性,具体操作步骤参照Zhang等的方法(Zhangetal.,2017)。
1.7 数据分析
应用Excel软件整理数据,并使用SPSS软件和Sigma Plot10.0进行统计分析和数据作图,数据用平均值+标准误(SE)表示,采用非配对T检验(unpaired t-test)和单因素方差分析(One-Way ANOVA)中的LSD方法进行差异显著性检验(P<0.05为差异显著,用*表示;P<0.01为差异极显著,用**表示)。
2 结果与分析
2.1 两个褐飞虱NlTret1基因序列功能分析
登录号为XP_022187716.2和XP_022188572.1的核苷酸序列分别有1 353 bp和1 488 bp的开放阅读框,编码具有450和495个氨基酸残基的蛋白质,将这两个蛋白序列分别命名为NlTret1和NlTret1 X1。进行理化分析,预测这两个蛋白分子量大小为49.984 kDa和53.059 kDa,理论等电点pI为6.53和7.46,其中NlTret1带负电荷氨基酸残基总数(Asp+Glu)为34,带正电荷氨基酸残基总数(Arg+Lys)为33,NlTret1 X1带负电荷氨基酸残基总数(Asp+Glu)为30,带正电荷氨基酸残基总数(Arg+Lys)为31。因此推测NlTret1属于酸性氨基酸,NlTret1 X1属于碱性氨基酸。它们的不稳定系数为34.63和34.38,脂肪族氨基酸指数为115.24和114.48,亲水平均系数分别为0.565和0.713,综合以上结果,表明NlTret1和NlTret1 X1都属于疏水性氨基酸。另由SMART预测NlTret1和NlTret1 X1的跨膜结构域模型分别包含10个跨膜结构域和12个跨膜结构域且定位于细胞膜,属于MFS超家族,并都具有特异性糖转运蛋白的功能结构域,说明这两个蛋白均为固定在细胞膜上的糖转运蛋白。从二级结构和三级结构预测结果可看出,NlTret1和NlTret1 X1主要由α-螺旋和无规卷曲组成(图1)。
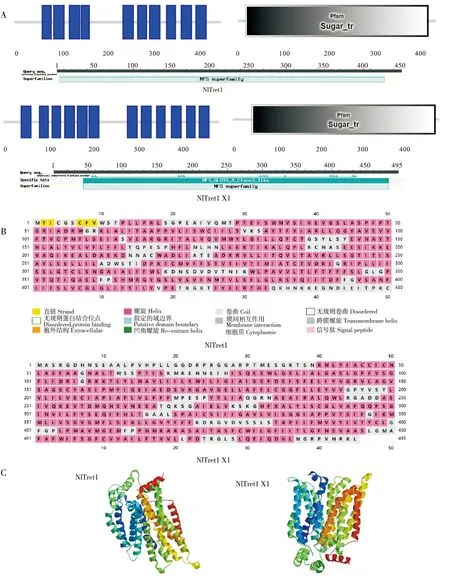
图1 NlTret1和NlTret1 X1的生物信息学分析Fig.1 Bioinformatics analysis of NLTRET1 and NLTRET1 X1注:A,NlTret1与NlTret1 X1跨膜结构域及保守结构域预测;B,NlTret1与NlTret1 X1的二级结构域预测;C,NlTret1与NlTret1 X1三级结构预测。Note:A,NlTret1 and NlTret1 X1 transmembrane domain prediction and conservative domain prediction;B,Prediction of the secondary domains of NlTret1 and NlTret1 X1;C,NlTret1 and NlTret1 X1 tertiary structure prediction.
2.2 两个褐飞虱NlTret1蛋白的进化树分析
从不同昆虫种类进行的系统发育树表明,两个蛋白聚集在相关的系统发育分支中,NlTret1与玉米蚜Rhopalosiphummaidis、高粱蚜Melanaphissacchari、温带臭虫Cimexlectularius和褐翅椿象Halyomorphahalys聚集在一支;NlTret1 X1与棉蚜Aphisgossypii、玉米蚜Rhopalosiphummaidis、高粱蚜Melanaphissacchari、烟蚜Myzuspersicae、蔗黄伪毛蚜Siphaflava和棕榈蓟马Thripspalmi集中在一支。NlTret1与NlTret1 X1蛋白都与其他昆虫的Tret1与Tret1 X1蛋白聚集在一支,并与上述多数互为半翅目的昆虫同源相近(图2)。

图2 NlTret1(XP_022187716.2)与NlTret1 X1(XP_022188572.1)基于氨基酸序列与其他昆虫Tret构建的进化发育树Fig.2 Evolution and development tree constructed by NlTret1 (XP_022187716.2) and NlTret1 X1(XP_022188572.1) based on amino acid sequence and other insect Trets注:Tret蛋白物种来源及其登录号:棉蚜Aphis gossypii,AgTret1X1(XP_027838003.1);玉米蚜Rhopalosiphum maidis,RmTret1-likeX1(XP_026808273.1)、RmTret1-like(XP_026819645.1);高粱蚜Melanaphis sacchari,MsTret1-likeX1(XP_025205476.1)、MsTret1-like(XP_025208158.1);烟蚜Myzus persicae,MpTret1-2X1(XP_022182394.1);蔗黄伪毛蚜Sipha flava,SfTret1-likeX1(XP_025417238.1);棕榈蓟马Thrips palmi,TpTret1-2X1(XP_034240573.1);白蜡窄吉丁Agrilus planipennis,ApTret1-likeX1(XP_018333082.1);黄翅菜叶蜂Athalia rosae,ArTret1-2X1(XP_012255542.1);丽蝇蛹集金小蜂Nasonia vitripennis,NvTret1-2X1(XP_008207267.1);红松蜂Osmia bicornis,ObTret1-2(XP_029042530.1)、ObTret1-like(XP_029049346.1);韭菜迟眼蕈蚊Bradysia coprophila,BcTret1-2(XP_037026138.1);油橄榄果实蝇Bactrocera oleae,BoTret1X1(XP_014096828.1);昆士兰实蝇Bactrocera tryoni,BtTret1X1(XP_039957802.1);桔小实蝇Bactrocera dorsalis,BdTret1X1(XP_011202667.1);家蚕Bombyx mori,BmTret1(XP_004932337.1);甘蓝尺蠖Trichoplusia ni,TnTret1-like(XP_026731345.1);菜粉蝶Pieris rapae,PrTret1-like(XP_022117370.1);君主斑蝶Danaus plexippus,DpTret1-like(XP_032527054.1);阿芬眼蝶Maniola hyperantus,MhTret1-like(XP_034833046.1);斑点木蝶Pararge aegeria,PaTret1-like(XP_039761312.1);温带臭虫Cimex lectularius,ClTret1-like(XP_014258243.1);褐翅椿象Halyomorpha halys,HhTret1(XP_014277273.1);多胚跳小蜂Copidosoma floridanum,CfTret1-like(XP_014215782.1);果园壁蜂Osmia lignaria,OlTret1-like(XP_034178871.1);西方蜜蜂Apis mellifera,AmTret1(XP_016771112.2);大蜜蜂Apis dorsata,AdTret1-like(XP_031366370.1);小蜜蜂Apis florea,AfTret1-like(XP_003695906.1).
2.3 RNAi后褐飞虱两个NlTret1基因沉默效率
为了鉴定RNAi的注射效果,利用qRT-PCR检测NlTret1和NlTret1X1在mRNA表达水平上变化情况。干扰NlTret1和NlTret1X1的48 h后本基因表达量都极显著下调(P<0.01),其中NlTret1与对照组相比靶标基因的表达量下调了88.1%,NlTret1X1下调了95%,这说明合成的dsRNA可以成功抑制靶标基因的表达水平(图3)。

图3 dsRNA注射48 h后褐飞虱NlTret1和NlTret1 X1基因相对表达量水平Fig.3 Relative expression levels of NlTret1 and NlTret1 X1 genes 48 h after dsRNA injection注:采用非配对T检验进行数据分析,误差用平均值的标准误差表示。“*”表示在P<0.05上差异显著,“**”表示在P<0.01差异极显著。使用绿色荧光蛋白GFP作为对照。Note:Unpaired t-test was used for data analysis,the error was expressed as the standard error of the mean value.“*”indicated a significant difference when P<0.05,and “**”indicated a extremely significant difference when P<0.01.GFP,green fluorescent protein was used as control.
2.4 RNAi后褐飞虱海藻糖代谢关键基因相对表达量变化
荧光定量PCR检测的结果显示,相比于对照组褐飞虱海藻糖酶基因NlTRE在RNAi后mRNA水平都有不同程度变化,在注射dsNlTret1和dsNlTret1X1的48 h后NlTRE1-1基因表现为超低水平表达(P<0.01),相反地NlTRE1-2基因则表现为高水平的表达(P<0.01);NlTRE1-3基因在注射dsNlTret1后表达量呈极显著上调,而注射dsNlTret1X1后则呈极显著下调现象。同时,检测到3个海藻糖-6-磷酸酯酶基因(NlTPS)表达量水平也不一致,注射两种靶标基因的dsRNA后NlTPS2和NlTPS3基因分别表现为极显著下调和极显著上调,NlTPS1基因则是在干扰NlTret1后为上调而在干扰NlTret1X1后为下调(图4)。

图4 dsTret1和dsTret1 X1注射48 h后褐飞虱体内海藻糖代谢通路3个NlTRE(TRE1-1、TRE1-2、TRE2)与3个NlTPS(TPS1、TPS2、TPS3)基因相对表达量水平Fig.4 Relative expression levels of three NlTREs (TRE1-1,TRE1-2,and TRE2) and three NlTPSs (TPS1,TPS2,and TPS3) genes in trehalose metabolic pathway in BPH after 48 h of dsTret1 and dsTret1 X1 injection注:采用单因素方差分析进行数据分析,误差用平均值的标准误差表示,星号代表与对照组比较的结果。“*”表示在P<0.05上差异显著,“**”表示在P<0.01差异极显著。使用绿色荧光蛋白GFP作为对照,下同。Note:One-way ANOVA was used for data analysis,the error was expressed as the standard error of the mean value,and asterisk represented the results compared to the control group.“*”indicated a significant difference when P<0.05,and “**”indicated a extremely significant difference when P<0.01.GFP,green fluorescent protein was used as control.The same as below.
2.5 RNAi后褐飞虱体内海藻糖酶活和糖含量变化
dsRNA能显著抑制靶基因,进一步检测到dsNlTret1和dsNlTret1X1对可溶性海藻糖酶活和膜结合海藻糖酶活也都有极显著的抑制效果(P<0.01)。糖原与海藻糖含量在干扰NlTret1基因48 h后检测到较对照组含量表现为极显著下降,而dsNlTret1对葡萄糖含量则无显著影响。干扰NlTret1X1基因的结果与之相反,糖原与海藻糖含量并无显著的变化,葡萄糖含量与对照组相比却极显著上升(图5~图6)。

图5 dsTret1和dsTret1 X1注射48 h后褐飞虱可溶性海藻糖酶与膜结合海藻糖酶酶活Fig.5 Soluble trehalase and membrane-bound trehalase activity of the brown planthopper after 48 h of dsTret1 and dsTret1 X1 injection

图6 dsTret1和dsTret1 X1注射48 h后褐飞虱糖原、海藻糖与葡萄糖含量Fig.6 Glycogen,trehalose and glucose concentrations of brown planthopper after 48 h of dsTret1 and dsTret1X1 injection
3 结论与讨论
主要的促进子超家族(MFS superfamily)是最大的转运蛋白超家族之一,目前包含82个家族,每个家族特定于一组化合物(Reddyetal.,2012;Nio-Gonzálezetal.,2019),在MFS超家族中,糖由糖转运蛋白St运输。虽然在几乎所有的GLUT超家族成员中都可以看到序列相似性,包括TRET1,但它们的生化特性差异仍十分大(Uldry and Thorens,2004)。另外,TRET1直系同源物之间的动力学特征也不同(Kanamorietal.,2010),这说明除了保守区域中的氨基酸残基外,其他氨基酸残基也可能负责每个转运蛋白的特异性。本研究用生物信息学的方法对NlTret1和NlTret1 X1蛋白进行分析,通过三级结构预测、跨膜结构域分析以及亚细胞定位表明NlTret1和NlTret1 X1蛋白是与细胞膜结合的跨膜蛋白(图1),拥有特异性转运糖类物质的结构域,因此NlTret1和NlTret1 X1为典型的糖转运蛋白,但它们的二级结构和保守结构域外的氨基酸残基存在差异,推测其糖运输功能可能会因此有所不同。
糖转运蛋白是负责把糖类物质运输到各种组织中的工具,在昆虫细胞内海藻糖转运蛋白则是转运海藻糖的主要蛋白并且对适应环境胁迫、平衡营养状态起着重要的作用(Thompsonetal.,2003)。本研究在证实了RNAi效果显著的前提下检测NlTRE与NlTPS的基因表达量,沉默NlTret1和NlTret1X1后褐飞虱3个海藻糖酶基因与3个海藻糖合成酶基因都显示了不同程度的超表达水平,这与Zhao等人敲除NlTRE结果几乎一致(Zhaoetal.,2016),表明海藻糖转运蛋白可影响TRE1和TRE2以及3个TPS之间的表达。不同的可溶性海藻糖酶(Soluble trehalase,简称TRE1)基因存在着不同分工(陈坚毅等,2017),在注射dsRNA后NlTRE1-1极显著降低(图4-A),这说明褐飞虱海藻糖转运蛋白NlTret1和NlTret1 X1对TRE1-1有着相同的调控功能,NlTRE1-1表达量减少后NlTRE1-2表达量显著增加(图4-B),一个基因的表达敲低时可激活其他基因的补充功能,这揭示了基因之间的相互作用。沉默NlTret1后NlTPS1与NlTPS3表达量显著增加(图4-D、图4-F)消耗糖原(图6-A)应积累海藻糖(於卫东等,2020),但似乎NlTret1可严重影响到NlTRE1-1基因的表达(图4-A),尽管最后降低了可溶性海藻糖酶活性与膜结合海藻糖酶活(图5),但是仍大量消耗海藻糖含量,最终结果表现为海藻糖含量的极显著降低(图6-B)。AgTret1缺陷型冈比亚按蚊Anophelesgambiae的血淋巴海藻糖浓度检测结果也与本实验结果一致,与对照组相比降低40%(Liuetal.,2013),抑制NlTret1表达可减少海藻糖从脂肪体到血淋巴的转运,这说明可通过NlTret1调整维持血淋巴的海藻糖浓度。相对来说,沉默NlTret1X1虽也使TRE与TPS基因产生不同程度的波动(图4),对海藻糖酶活的影响也是显著降低的(图5),但并不影响到糖原含量与海藻糖含量的变化,仅使葡萄糖浓度升高(图6-C)。Tret1可能属于GLUT家族的新成员,有研究表明嗜眠摇蚊Polypedilumvanderplanki的Tret1不仅可以转运海藻糖还可转运葡萄糖类似物。此外,尽管果蝇Drosophilamelanogaster的Tret1-2含有保守的Tret1氨基酸序列,但其并不具备运输海藻糖的能力(Kikawadaetal.,2007;Kanamorietal.,2010)。因此推测NlTret1与NlTret1 X1这两个蛋白分别执行不同的功能,在此之中NlTret1作为特异性转运海藻糖的蛋白可能性更大,而NlTret1 X1也许以运输葡萄糖为主,由实验结果来看可能还存在着功能互补的作用,但相关机制还需进一步研究与验证。
