不同龄期的大草蛉对桃蚜的捕食作用
2022-09-22刘兴旺李国慧彭秀丽
陈 斌,张 伟,刘兴旺,黄 英,王 玲,李国慧,彭秀丽
(1.四川省高县农业技术推广站,四川宜宾 645150;2.宜宾学院农林与食品工程学部,宜宾 644000;3.四川省绵阳市植保植检站,四川绵阳 621000;4.四川省广元市利州区农业农村局,四川广元 628017;5.四川省北川县植保植检站,四川绵阳 622750;6.四川省夹江县农业农村局,四川乐山 614100)
桃蚜Myzuspercicae属半翅目蚜科,是严重危害桃Prunuspersica、梨Pyrusspp.、李Prunussalicina、甘蓝Brassicaoleracea、萝卜Raphanussativus、辣椒Capsicumannuum和烟草Nicotianatabacum等果蔬经济作物的重要害虫,也是黄瓜花叶病毒(Cucumbermosaicvirus,CMV)、马铃薯Y病毒(PotatovirusY,PVY)等115种植物病毒的重要传播媒介,其分泌的蜜露还能引起果树煤烟病(赵荣乐等,2003;王化兵,2015;易齐等,2020)。桃蚜在我国大部地区均有发生,在华北地区年发生10余代(司生云等,2012),在长江中下游地区年发生20~30代,在上海、江苏、四川等气候适宜区发生尤为严重。目前,防治桃蚜的主要措施是化学农药,而长期连续多次、高剂量使用化学农药防治造成桃蚜抗药性迅速上升,农药残留、污染环境、对人畜造成毒害等问题越来越突出,迫切需要寻找新的治理措施(姚洪渭等,2002;宫亚军等,2011)。生物防治是解决桃蚜生态控制的重要措施,保护和利用天敌昆虫是其中重要的组成部分。近年来,随着农药减量控害概念的提出,研究和利用昆虫天敌控制害虫成为研究热点,30年来我国利用天敌昆虫防治农业害虫取得了可喜的成果(杨怀文,2015a;2015b)。我国天敌资源丰富,据有关文献报道我国昆虫天敌仅寄蝇约有450种,捕食螨约200种,捕食性农田蜘蛛有265种,仅广西壮族自治区捕食性昆虫有313种(杨怀文,2015a;2015b)。
大草蛉Chrysopapallens(Rambur)属脉翅目草蛉科,是农业生产上重要的天敌昆虫之一(Wyckhuysetal.,2013),因其食量大、分布广、数量多、对害虫控制效果好,早在上世纪80年代就引起我国生防工作者重视并对其开展研究(牟吉元等,1980;赵敬钊,1988)。近十年来,有关大草蛉对害虫捕食能力的研究发现,大草蛉成虫及幼虫可以捕食蚜虫(赵琴等,2008;程丽媛等,2014)、粉虱及卵(刘爽等,2011;王然等,2016;唐天成等,2018)、蓟马(唐良德等,2017)、叶螨(张欣等,2012)、鳞翅目害虫卵及幼虫(李姝等,2019;徐庆宣等,2019;曹雯星等,2020),大草蛉甚至还可捕食多异瓢虫Adoniavariegata、七星瓢虫Coccinellaseptempunctata等一些捕食性天敌昆虫的低龄幼虫(阿力甫·那思尔等,2014,2015)。
大草蛉繁殖力高,且人工繁育技术已较成熟(党国瑞,2013;潘鹏旭,2017),卵即使长时间低温保存,也有较高孵化率(Zhangetal.,2019),有利于规模化繁殖。鉴于大草蛉广阔的生物防治应用前景,为明确大草蛉对桃蚜的控害效果,本文利用功能反应模型测定了大草蛉对桃蚜的捕食作用,并分析了种内干扰对捕食作用的影响,以期为开展桃蚜生物防治提供参考。
1 材料与方法
1.1 供试虫源
桃蚜和大草蛉采自四川宜宾高县田间。桃蚜采回后用甘蓝苗扩繁,大草蛉从田间采回后用桃蚜作为饲料进行饲养(王海建,2015),饲养条件均为温度25℃ ± 1℃、相对湿度70% ± 10%、光周期14 L ∶10 D。通过对蜕皮次数的观察,得到1龄、2龄、3龄幼虫和成虫(王海建,2015)。
1.2 试验方法
1.2.1不同龄期大草蛉对桃蚜的捕食功能反应
试验在光照培养箱内进行,试验条件为温度25℃、光照时间14 h、相对湿度70%。将参试的大草蛉雌成虫(交配后1 d)和1~3龄幼虫保存在4℃~5℃冷藏箱内,饥饿处理24 h。参照王海建(2015)的方法,在直径为25 cm的大烧杯放入新鲜甘蓝叶片,叶片下垫蘸水的脱脂棉保湿,接入1日龄桃蚜若蚜后,引入1日龄大草蛉1头。分别测定大草蛉雌成虫对桃蚜捕食功能反应时,桃蚜密度设置为50、100、150、200、250、300头。分别测定大草蛉1龄、2龄和3龄幼虫对桃蚜捕食功能反应时,桃蚜密度均设为20、30、40、50、60、70头。每杯引入大草蛉1头,各处理均设8次重复,并设仅放桃蚜的等量密度对照组一个。24 h后观察并记录各处理剩余的桃蚜数量,并以对照组自然死亡桃蚜数量进行校正,建立捕食功能反应模型(王海建,2015)。用Holling Ⅱ 圆盘方程Na=a′·N·Tt/( 1+a′·N·Th)拟合大草蛉成虫和不同龄期幼虫对桃蚜的捕食功能反应,式中Na为被捕食的桃蚜数量,a′为瞬时发现率,N为桃蚜密度,Th为处置时间,Tt为试验持续时间(试验为24 h,即1 d,取Tt=1);参照吴坤君等的方法计算功能反应模型参数,并对所建立方程进行χ2检验(李桂亭,2002;吴坤君等,2004)。
1.2.2种内干扰对不同龄期大草蛉捕食率的影响
试验在光照培养箱内进行,试验条件为温度25℃、光照时间14 h、相对湿度70%。参照李新兵等的方法(李新兵等,2019),在6个直径为25 cm的干燥洁净的培养皿放入新鲜甘蓝叶片,叶片下垫蘸水的脱脂棉保湿,以50、100、150、200、250 头/皿桃蚜分别与1、2、3、4、5 头/皿大草蛉成虫组合,8次重复,并设仅放桃蚜的等量密度对照组。24 h后观察并记录各组合桃蚜被捕食的数量,并以对照组桃蚜自然死亡数量进行校正。采用HasseⅡ干扰反应模型E=QP-m拟合种内干扰对大草蛉捕食率的影响,其中E为捕食率,Q为搜索常数,P为捕食者密度,m为干扰系数(Holling,1959)。将方程取对数后线性化,用最小二乘法求P、m值(李秋荣等,2020);并对所建立方程进行χ2检验(Hassell,1978)。
2 结果与分析
2.1 大草蛉成虫对桃蚜的捕食功能反应

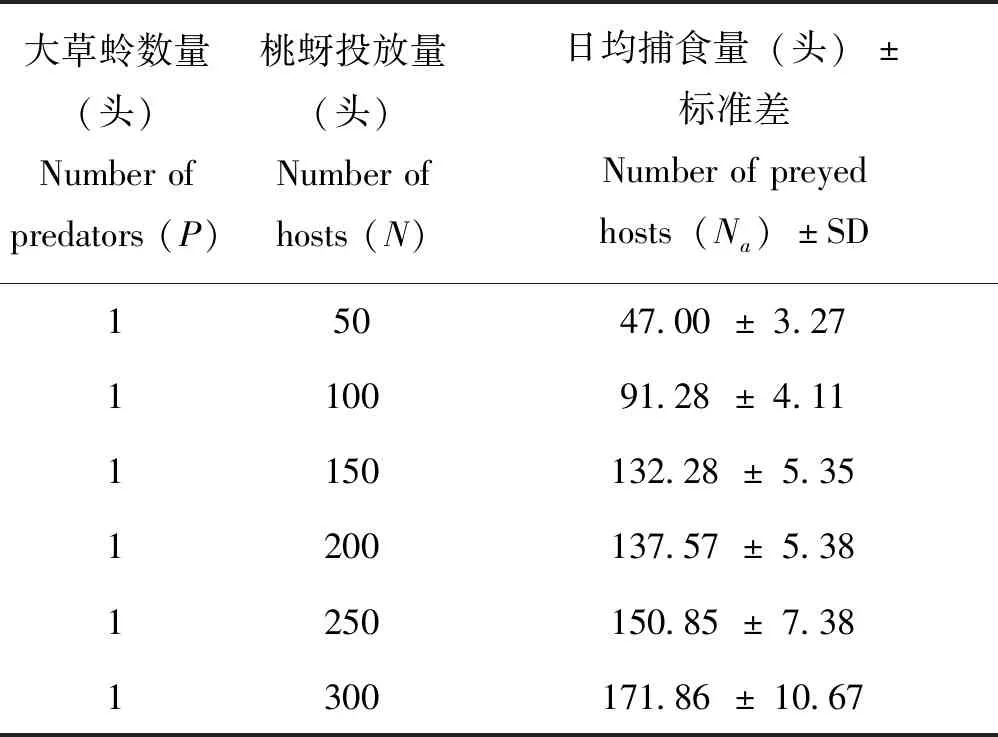
表1 大草蛉成虫对桃蚜的捕食功能反应Table 1 Predatory capacity of adult of Chrysopa pallens to Myzus persicae
2.2 大草蛉幼虫对桃蚜的捕食功能反应
2.3 种内干扰对大草蛉捕食率的影响


表2 大草蛉幼虫对桃蚜的捕食功能反应Table 2 Predatory capacity of larvae of Chrysopa pallens to Myzus persicae
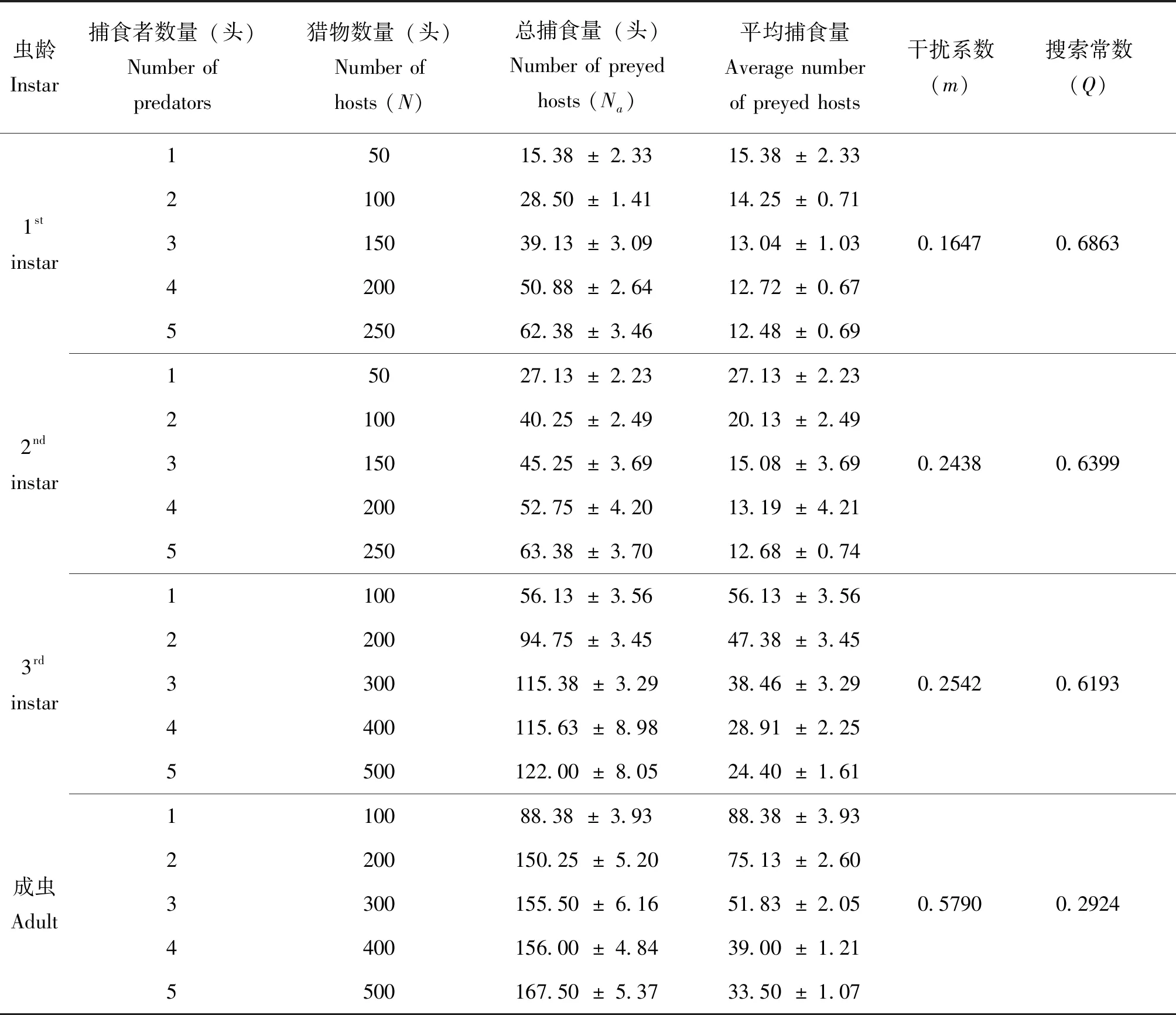
表3 种内干扰对大草蛉捕食作用的影响Table 3 Intraspecific interference on Chrysopa pallens predation rates
3 结论与讨论
近年来,天敌昆虫对桃蚜的捕食作用研究较多。大肩突瓢虫Synonychagrandis(Thunberg)(欧善生等,2008)、异色瓢虫Harmoniaaxyridis(李英梅等,2015)和四斑广盾瓢虫Platynaspismaculosa(陈燕等,2015)在实验室内对桃蚜均有较好的控制效果,理论日最大捕食量分别为277.4头、200.0头和81.3头,但均低于本研究中大草蛉成虫对桃蚜的理论日最大捕食量,表明大草蛉对桃蚜有较强的控制潜能。
捕食功能反应模型反映了天敌和害虫种群动态变化过程(Solomon,1949),Holling提出3个经典的功能反应模型,即Honlling Ⅰ型、Holling Ⅱ型和Honlling Ⅲ型(Honlling,1959)。Holling Ⅱ圆盘方程模型是最常见的捕食者-猎物功能反应模型,大量昆虫天敌对害虫的捕食效应都符合该模型。日本通草蛉Chrysoperlanipponensis、大草蛉、叶色草蛉Chrysopaphyllochroma、丽草蛉Chrysopaformosa、黄玛草蛉Malladabasalis、白面叉草蛉Pseudomalladaalbofrontata等草蛉科天敌对害虫的功能反应绝大部分都符合Holling Ⅱ圆盘方程模型(赖艳等,2020)。本研究结果显示大草蛉成虫和幼虫对桃蚜的捕食功能反应均符合Holling Ⅱ圆盘方程模型,即随着桃蚜投放量的增加,大草蛉捕食量呈加速增长趋势,但当桃蚜投放量超过 150头后,大草蛉成虫捕食量增长速度趋缓,1龄、2龄和3龄幼虫捕食量增长情况也呈相似趋势。该结果与大草蛉对夹竹桃蚜Aphisnerii(赵琴等,2008)、麦二叉蚜Schizaphisgraminum(李桂亭等,2002)、桃粉大尾蚜Hylopterusamygdali(孙丽娟等,2013)、牛蒡长管蚜Uroleucongobonis(孙丽娟等,2013) 、豆蚜Aphiscraccivora(唐良德等,2017)和桃粉蚜Hyalopterusarundimis(张安盛等,2004)等蚜虫的功能反应一致。而大草蛉对烟粉虱Bemisiatabaci(刘爽等,2011)、西花蓟马Frankliniellaoccidentalis(Sarkaretal.,2019)等比蚜虫个体更小的害虫的捕食功能反应则符合Honlling Ⅲ型,这可能与大草蛉对不同猎物取食偏好(Sarkaretal.,2019)以及猎自身的逃逸能力有关(Zhangetal.,2010)。
瞬时发现率a′与处置时间Th的比值,反映了天敌对猎物的消耗效率,a′/Th比值越大说明天敌对害虫的控制能力越强(周集中等,1986)。大草蛉1龄幼虫、2龄幼虫、3龄幼虫和成虫a′/Th比值分别为20.6、107.7、184.5和379.6,Holling Ⅱ圆盘方程模型拟合的理论日最大捕食量也随着虫龄的增加而增加,表明大草蛉成虫对桃蚜的捕食能力最强。该结果与大草蛉对桃粉大尾蚜Hyalopterusamygdali(Blanchard)、绣线菊蚜Aphiscitricolavan der Goot和牛蒡长管蚜Uroleucongobonis(Matsumura)的捕食效果一致(孙丽娟等,2013)。大草蛉成虫期对上述3种蚜虫的捕食量显著高于幼虫期,其中对绣线菊蚜的理论日最大捕食量可达766.0头。尽管大草蛉成虫捕食量大,但在生防应用中普遍存在定殖性差的问题(武鸿鹄等,2014),在生产中应用大草蛉控制桃蚜为害时,应优先考虑释放幼虫。
大量室内研究表明,捕食者密度的增加,捕食者之间将产生相互干扰,导致捕食效应下降,这一现象被称为种内干扰(Butleretal.,1979;王保平等,2001)。大草蛉捕食草地贪夜蛾Spodopterafrugiperda低龄幼虫(曹雯星等,2020)、黑刺粉虱Aleurocanspinfetus(唐天成等,2018)等害虫存在种内干扰现象。本研究结果证明大草蛉在捕食桃蚜时也存在种内干扰,且随着虫龄的增加种内干扰作用增强。种内干扰不仅与天敌密度相关,还与猎物密度有关(邱海燕,2020),因此在田间释放大草蛉控制桃蚜时,必须综合考虑害虫种群密度和大草蛉的种内干扰作用,恰当掌握天敌释放数量,达到控制害虫危害及维持天敌种群持续生存的目标。
本研究结果表明,应用大草蛉开展桃蚜生物防治有较好前景。但本研究是在实验室条件下进行的,田间天敌昆虫对害虫的防治效果还受到环境温湿度(贾静静等,2019)、寄主植物(王甦等,2016)、天敌性别(金剑雪等,2011)、农药施用(李锐等,2014)等多种因素的影响,其田间释放技术还需要进一步深入研究。
