家蚕 BmChd64基因的克隆与表达定位分析
2022-09-22魏秋兰吴琼芳邓惠敏朱子丹
魏秋兰,吴琼芳,邓惠敏,朱子丹
(华南师范大学生命科学学院,华南师范大学昆虫科学与技术究所,广东省昆虫发育生物学与应用技术重点实验室,广州 510631)
昆虫是陆地上种类最多、数量最大且分布最广的生物。在长期自然演化过程中,昆虫形成了蜕皮与变态这一特殊且重要的发育特征,而这一特征主要受到蜕皮激素(20-hydroxyecdysone,20E)和保幼激素(Juvenile hormone,JH)协同调控(Riddifordetal.,2003;Jindraetal.,2013)。20E与其受体复合物EcR-USP(Ecdysone receptor,EcR;Ultraspiracle,USP)结合后,启动一系列级联反应进而调控蜕皮与变态相关基因的表达,导致蜕皮与变态(Riddifordetal.,2000;2008;Yamanakaetal.,2013)的发生。JH则与其受体MET/Tai(Methoprene,MET;Taiman,Tai)结合,启动其下游的初级反应基因Kruppelhomolog1(Kr-h1)表达,进而启动了JH级联信号通路,调控昆虫的生长与发育(Kayukawaetal.,2012;2017)。近年来许多研究逐步完善了20E与JH调控昆虫生长与变态的机制,但二者确切的互作机制有待深入阐释。Li等发现,果蝇钙调同源结构域蛋白Chd64(Calponin homology domain protein,21-kDa Calponin-like protein,Chd64)可与JH反应元件(JH-response element,JHRE)结合,还可通过与核受体Methoprene-tolerant(Met)和USP互作响应JH诱导基因的转录表达(Lietal.,2007)。另一个果蝇Chd蛋白DroVav被认为是信号转导蛋白与酪氨酸磷酸化的表皮生长因子受体(Dekeletal.,2000)。棉铃虫HelicoverpaarmigeraChd64同源蛋白HaCal响应20E诱导后以磷酸化形式进入细胞核,受JH analog methoprene(JHA)诱导后以非磷酸化形式与USP1互作进而参与调控20E或JH的信号级联(Liuetal.,2011)
Chd64是一类含有CH结构域的蛋白。CH结构域一般含100~110个氨基酸残基,它是一类肌动蛋白结合结构域,最初发现于钙结合蛋白(Calponin)的氨基末端,后来发现在多种肌动蛋白结合蛋白和信号转导蛋白中也存在CH结构域(Gimonaetal.,2002;Ferjanietal.,2010)。根据蛋白序列的不同,含有CH结构域的蛋白质主要分为3类:氨基末端含单一CH结构域的蛋白质、由两个CH结构域串联形成丝状肌动蛋白结合结构域(F-actin-binding domain,ABD)的蛋白质及含两个ABD结构的蛋白质(Korenbaum and Rivero,2002)。蛋白质中两个串联的CH结构域构成了肌动蛋白结合区,例如血影蛋白、肌动蛋白、肌养蛋白和纤维蛋白(Hartwigetal.,1994;McGoughetal.,1998)。研究认为,大部分含CH结构域的蛋白质都能与肌动蛋白相结合或者参与细胞的信号转导(Banuelosetal.,1998)。果蝇中含CH结构域的信号转导蛋白DroVav作为一个信号转导器在果蝇发育中发挥着关键作用(Dekeletal.,2000)。Liu等发现在棉铃虫中含CH结构域的类肌钙蛋白HaCal在20E和JH的通路网络中发挥重要作用,调节了相关基因的表达从而影响棉铃虫的生长发育(Fuetal.,2009;Liuetal.,2011)。在赤拟谷盗和虾中,Chd64也在20E和JH调控的通路中发挥了重要作用(Sinetal.,2014;Tarczewskaetal.,2015)。
基于Chd64蛋白在果蝇和棉铃虫生长发育中激素信号传导途径的重要作用,本论文克隆了鳞翅目模式昆虫同时也是重要经济昆虫家蚕中与果蝇DmChd64同源的BmChd64基因,并对该蛋白氨基酸序列进行生物信息学预测,同时利用实时荧光定量PCR(Quantitative Real-time PCR,qRT-PCR)检测了该基因在家蚕中的时空表达,并利用共聚焦显微镜进行了该蛋白的亚细胞定位分析,以期丰富家蚕等全变态昆虫变态发育分子调控机理的研究,为家蚕发育的人为调节与害虫的生物防治提供新的思路。
1 材料与方法
1.1 实验材料和主要试剂
本研究选用的家蚕品系为大造,蚕卵由广东省农业科学院蚕业与农产品加工研究所提供,家蚕幼虫定时以桑叶饲喂,于温度26℃,相对湿度65%~75%,光周期L ∶D=12 ∶12条件下培养至成虫。大肠杆菌DH5α、BL21、原核表达载体pET-28a及真核表达载体pIE1-EGFP-N1均由本实验室保存。Trizol Isolate抽提试剂盒、pMD-18T载体、DNA限制性内切酶、T4连接酶、Taq酶、低分子量标准蛋白marker、DNA marker、M-MLV反转录试剂盒和实时荧光定量PCR试剂盒均购自TAKARA公司;质粒提取和DNA胶回收试剂盒为TianGen产品;实验所用其他试剂均为国产分析纯试剂;实验涉及的引物合成和相关DNA测序委托华大基因公司完成。
本研究所用BmN细胞株来源于家蚕卵巢细胞,为本实验室培养与保存,细胞培养条件为27℃恒温,所用培养基为含有10%胎牛血清(Fetal Bovine Serum,FBS)的Grace培养基(GIBCO,USA)。细胞每隔3 d以1 ∶3的比例稀释培养基作继代培养。后均匀铺板于24孔细胞培养板,待细胞密度长至80%~90%时进行转染和后续实验。
1.2 家蚕总RNA的提取与cDNA合成
选取处于5龄游走期的家蚕,脂肪体、中肠、表皮和翅原基组织;取5龄至蛹期的家蚕,隔天取翅原基组织,并于液氮中速冻后研磨。按Trizol Isolate抽提试剂盒的说明提取家蚕组织的RNA,用紫外分光光度计和1%琼脂糖凝胶电泳检测RNA的纯度和浓度及质量,然后按照TAKARA公司的M-MLV RTase cDNA Synthesis Kit(反转录试剂盒)说明书,反转录合成第一链cDNA。
1.3 家蚕 BmChd64基因的克隆及序列分析
以果蝇中的DmChd64(Acession No:AAF47840)为参照基因,在NCBI上通过Blast分析对比出家蚕中对应的同源基因,即BmChd64(Accesion No:NP_001040372;相似性:81%;E-value:4e-109)。根据序列设计PCR引物(见表1),以家蚕cDNA为模版,进行PCR扩增,反应条件为:95℃预变性3 min;94℃变性30 s,55℃退火30 s,72℃延伸30 s,30个循环,最后72℃延伸10 min。反应结束后,用1%琼脂糖凝胶电泳检测PCR产物。参照DNA胶回收试剂盒的方法进行胶回收。将胶回收纯化后的PCR产物与pMD18-T载体连接,转化到DH5α菌体中,筛选阳性克隆进行测序。使用DNAstar软件中的Clustal Alignment Programe对测序后的序列进行比对鉴定。基因序列比较与进化树构建使用DNAStar软件的Clustal W方法。

表1 本研究所使用到的引物Table 1 Primers used in this study
1.4 家蚕BmChd64的原核表达和纯化
用BamHⅠ和XhoⅠ对目的DNA片段和pET-28a载体进行双酶切,反应体系为质粒10 μL,BamH I酶1 μL,XhoⅠ酶1 μL,10 × K Buffer 3 μL,ddH2O 5 μL。反应条件为37℃ 3 h。胶回收后将二者连接,构建重组质粒pET-28a-BmChd64,将重组质粒转化到BL21菌体中,加入最终浓度为1 M IPTG进行诱导表达,SDS-PAGE检测表达情况。为了确定重组蛋白是否以可溶的形式表达,收集经IPTG诱导的菌体后,将菌体进行超声破碎,分别收集沉淀和上清液,在12%凝胶上进行SDS-PAGE检测。蛋白纯化按照Ni-NTA His·Bind Resin(Novagen)试剂盒的方法进行。取1 mL Ni2+柱于管中,使用2倍体积蒸馏水洗2次,2倍体积1×Charge (50 mM NiSO4)缓冲液洗3次,2倍体积1×Binding buffer(0.5 M NaCl,20 mM Tris-HCl,5 mM Imidazole,pH7.9)缓冲液洗2次,将收集的上清液用0.45 μM的滤膜过滤后加入管中,分别以10倍体积的1×Binding buffer、6倍体积的1×Washing buffer (0.5 M NaCl,20 mM Tris-HCl,60 mM Imidazole,pH7.9)和6倍体积的1×Elute buffer (0.5 M NaCl,20 mM Tris-HCl,1 M Imidazole,pH7.9)洗柱子,离心收集上清,该上清含纯化蛋白。用SDS-PAGE检查蛋白纯化效果。
1.5 BmChd64在家蚕不同发育时期各组织中的表达分析
以家蚕各发育时期翅原基cDNA和5龄游走期翅原基、表皮、脂肪体和中肠组织cDNA为模版,以家蚕Rp49(Gene ID:778453)作为内参基因,参照qRT-PCR试剂盒的方法(TAKARA,大连),采用20 μL体系,10 μL SYBR Premix Ex TaqTM,上、下游引物各0.8 μL,6.8 μL ddH2O,2 μL DNA模板。使用7300 Real-time PCR System进行PCR反应,PCR反应程序如下:95℃变性30 s,40个循环,循环条件为95℃ 3 s,60℃ 30 s。每个样品重复进行3次,检测BmChd64的表达。为增加数据的可比性,需提前检测目标基因和内标基因的扩增效率是否一致。然后采用2法来确定每一个目标基因相对内标基因Rp49的相对表达量。用Graphpad Prism 5统计分析软件,采用ANOVA(多个处理间的两两比较)或独立样本T测验(两个样品间比较)来进行处理间的差异比较分析。
1.6 BmChd64的亚细胞定位
将PCR扩增得到的BmChd64的ORF片段插入到pMD18-T中,用BamHⅠ和XhoⅠ进行双酶切后,将目的DNA片段插入到真核表达载体pIE1-EGFP-N1上,并通过双酶切验证重组质粒pIE1-BmChd64-EGFP。
转染前一天,加入完全培养基培养BmN细胞过夜,至细胞密度80%~95%时,进行转染。转染时,去除细胞培养皿中的完全培养基,用无血清(FBS)的Grace’s Insect培养基清洗细胞两次,再加入适量的无FBS的Grace’s Insect培养基到培养皿中,然后加入200 μL转染混合物,轻轻混匀,培养8 h后,更换含10% FBS的Grace’s Insect培养基,继续培养。24 h后,通过激光共聚焦显微镜观察带有GFP标签的BmChd64,进行亚细胞定位。
2 结果与分析
2.1 BmChd64基因克隆与序列分析
以家蚕cDNA为模版,对BmChd64基因进行PCR扩增,产物经1%凝胶电泳分离后在500~700 bp中间处显示出一条特异性条带(图1)。对该特异性PCR产物进行胶回收后与pMD18-T载体连接,通过PCR检测阳性克隆。提取阳性克隆细胞的质粒进行DNA测序,测序结果显示该PCR产物与GenBank的相应序列相同。
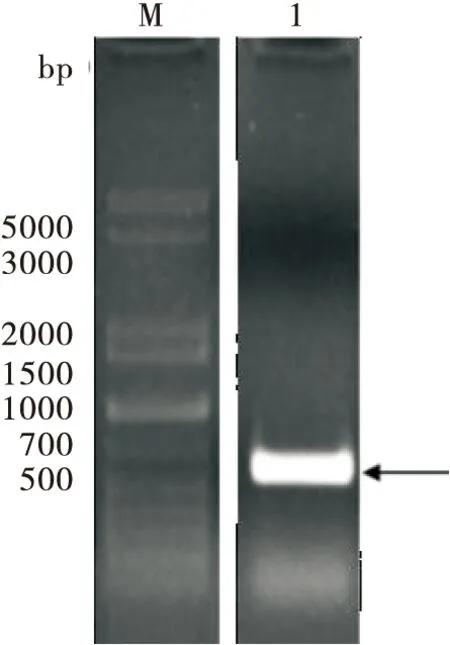
图1 BmChd64基因的PCR扩增Fig.1 PCR amplification of BmChd64 gene注:M,5000 DNA分子量标准;1,BmChd64基因的PCR扩增。Note:M,DL5000 DNA marker;1,PCR amplification of BmChd64 gene.
对克隆得到的BmChd64的cDNA序列进行分析。结果显示,该基因开放阅读框(ORF)的碱基数为567 bp,编码188个氨基酸。BmChd64蛋白的分子量大小为20.9 kDa,理论等电点为8.41。该基因所编码的蛋白在第27~130个氨基酸处存在CH结构域(图2)。

图2 BmChd64核苷酸及其编码的氨基酸序列Fig.2 Nucleotide and deduced amino acid sequences of BmChd64注:阴影序列显示CH域。下划线,5′非翻译区和3′非翻译区;红色字体,起始密码子与终止密码子;***,翻译终止位点;黄色底纹,CH结构域。Note:Shadow sequence showed a CH domain.Underline,5′ untranslated region and 3′ untranslated region;Red font,Start codon and stop codon;***,Translation termination site;Yellow shading,CH domain.
2.2 BmChd64的同源性比对与进化分析
为了解Chd64蛋白在昆虫中的功能是否保守,将家蚕BmChd64的氨基酸序列与埃及伊蚊Aedesaegypti、冈比亚按蚊Anophelesgambiae、棉铃虫Helicoverpaarmigera、体虱Pediculushumanuscorporis、赤拟谷盗Triboliumcastaneum、黑腹果蝇D.melanogaster、果蝇D.mojavensis及豌豆蚜Acyrthosiphonpisum进行同源序列比对和相似性分析。结果显示,BmChd64与其它昆虫的同源氨基酸序列具有极高的相似性,其中,与赤拟谷盗的相似性最高为84%,与DmChd64的相似性为80.9%。不同昆虫间同源氨基酸序列的高相似性(图3和表2),暗示了该基因在不同昆虫中的功能较保守,推测其在昆虫中可能发挥相似的功能。基于同源性构建了系统进化树,结果显示,BmChd64与鞘翅目赤拟谷盗的亲缘关系最近,但与同为鳞翅目的棉铃虫的HaCal亲缘关系较远(图4)。

图3 不同昆虫的Chd64蛋白的氨基酸序列比对Fig.3 Amino acid sequence alignment of Chd64 protein from different insects注:Chd64蛋白来源及GenBank登录号。1,家蚕 Bombyx mori transgenlin (Chd64) protein,NP_001040372.1;2,赤拟谷盗 Tribolium castaneum,XP_975100.1;3,埃及伊蚊 Aedes aegypti,XP_001652323.1;4,果蝇 Drosophila mojavensis,XP_015015904.1;5,黑腹果蝇 Drosophila melanogaster Cdh64,NP_001261398.1;6,冈比亚按蚊 Anopheles gambiae,XP_321834.4;7,豌豆蚜 Acyrthosiphon pisum calpoin-like,NP_001191867.1;8,体虱 Pediculus humanus corporis,XP_002426427.1;9,棉铃虫 Helicoverpa armigera calpoin,XP_021189834.1。
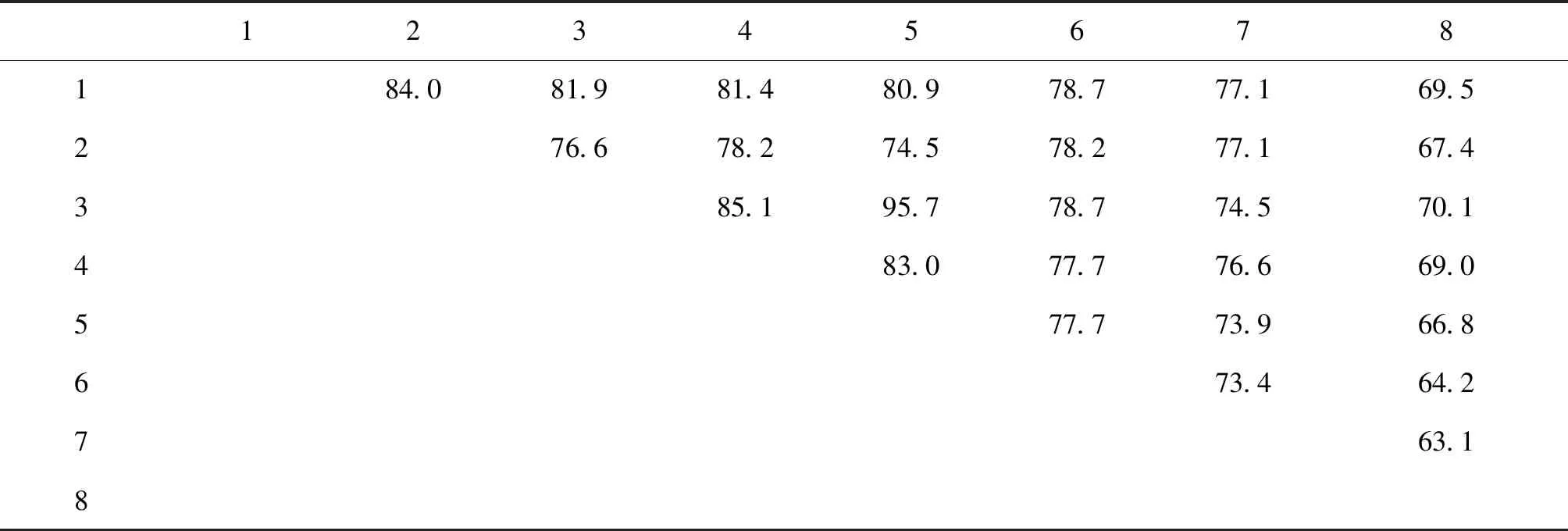
表2 不同昆虫Chd64蛋白的氨基酸序列相似性(相似度(%))Table 2 Amino acid sequence identity of Chd64 protein from different insects(Identity)

图4 BmChd64与其它昆虫同源氨基酸序列的系统进化树Fig.4 Phylogenetic tree of Chd64 protein from different insects
2.3 BmChd64重组蛋白的表达与纯化
将重组质粒pET-28a-BmChd64转化到大肠杆菌,在37℃,200 rpm的培养条件下,加入IPTG诱导蛋白表达,以经IPTG诱导的pET-28a菌体蛋白作为对照。SDS-PAGE检测结果显示,在约26 kDa处出现一条特异性条带(图5-A,泳道2),因pET-28a含有2个His标签,每个标签蛋白为3 kDa,预测BmChd64蛋白的分子量为 20.9 kDa,出现的目的条带与预测的pET-28a-BmChd64蛋白大小相近,这表明BmChd64重组蛋白在原核系统中成功表达。将菌体经过超声波破碎后,在上清和沉淀中都检测到重组蛋白(图5-A,泳道3和4),表明重组蛋白以可溶的形式表达。蛋白纯化后,在小于29 kDa处出现纯化的特异性蛋白条带,与预测的pET-28a-BmChd64蛋白约26 kDa大小一致(图5-B,泳道1和2)。

图5 BmChd64重组蛋白表达的SDS-PAGE分析Fig.5 SDS-PAGE analysis of expression of BmChd64 protein注:A,BmChd64重组蛋白诱导表达;M,蛋白分子量标准;1,经IPTG诱导的pET-28a菌体蛋白;2,经IPTG诱导的菌体重组总蛋白;3,诱导的菌体重组蛋白上清;4,诱导的菌体重组蛋白沉淀。B,BmChd64重组蛋白纯化;M,蛋白分子量标准;1、2,纯化后BmChd64重组蛋白。Note:A,Induced expression of BmChd64 recombinant protein;M,Standard protein maker;1,pET-28a bacterial protein induced by IPTG;2,Recombinant total protein of pET-28a-BmChd64 induced by IPTG;3,Supernatant recombinant protein of pET-28a-BmChd64 induced by IPTG;4,Precipitate recombinant protein of pET-28a-BmChd64 induced by IPTG.B,Purification of BmChd64 recombinant protein;M,Standard protein maker;1 and 2,BmChd64 recombinant protein after purification.
2.4 BmChd64在家蚕各发育时期各组织中的表达分析
利用qRT-PCR分别分析了家蚕5龄游走期脂肪体、中肠、表皮和翅原基中BmChd64的表达量和5龄至蛹期的翅原基中BmChd64基因的mRNA的表达情况。结果显示,BmChd64在家蚕5龄游走期的各组织都有一定量的表达,BmChd64在脂肪体中的表达量最低,在翅原基中的表达量最高,其次是表皮和中肠(图6),BmChd64在翅原基的高表达暗示其与变态发育中翅原基的发育相关。在翅原基中,BmChd64在5龄末期和游走期都有较高水平的表达,在蛹初期表达下降,到蛹期 4 d表达又升高,至蛹末期表达量最高(图7)。BmChd64在翅原基的表达模式与家蚕体内20E的滴度变化高度一致(杨鑫华,2010;Wangetal.,2018),暗示其在翅原基的表达可能与20E有关。

图6 qRT-PCR检测BmChd64在脂肪体、中肠、表皮和翅原基中的表达Fig.6 qRT-PCR analysis of expression profiles of BmChd64 in the fatbody,midgut,epidermis and wing disc注:EP,表皮;MG,中肠;FB,脂肪体;WD,翅原基。不同字母表示两者之间显著差异,P<0.05。Note:EP,Epidermis;MG,Midgut;FB,Fat body;WD,Wing disc.Different letters indicated significant differences,P<0.05.

图7 qRT-PCR检测BmChd64在翅原基中的表达Fig.7 qRT-PCR analysis of expression profile of BmChd64 in the wing disc注:L,幼虫期;D,天;W,游走期;PP,预蛹期;P,蛹期。数字表示在相应时期的天数。不同字母表示两者之间显著差异,P<0.05。Note:L,Larval stage;D,Day;W,Wandering;PP,Prepupal stage;P,Pupal stage.The numbers indicated the days in the corresponding stages.Different letters indicated significant differences,P<0.05.
2.6 BmChd64蛋白的亚细胞定位
蛋白质的功能与其亚细胞定位密切相关,通过了解蛋白质的亚细胞定位信息,可为探究蛋白质的生物学功能提供线索。为检测BmChd64在细胞内的定位,本论文构建了绿色荧光蛋白(GFP)标记的BmChd64蛋白表达载体,通过双酶切验证pIE1-BmChd64-EGFP载体构建成功。用构建好的pIE1-BmChd64-EGFP转染BmN细胞株,并通过共聚焦显微镜检测荧光信号。结果显示,转染了EGFP载体的细胞中,荧光信号在细胞核和细胞质中均匀分布,且细胞质比细胞核的绿色荧光更强。在转染了BmChd64-pEGFP载体的细胞中,荧光信号虽在细胞核和细胞质中都有分布,但围绕细胞核的细胞质区域的荧光明显更暗,出现一个暗圈,而细胞核中荧光信号更强(图8-D,F),暗示了细胞质中的BmChd64蛋白也可能入核,其主要是在细胞核中起作用。

图8 BmChd64在BmN细胞中的定位Fig.8 Localization of BmChd64 in BmN cells注:A,BmN转染EGFP质粒后绿色荧光图像;B,BmN转染EGFP质粒后白光图像;C,BmN转染EGFP质粒后荧光和白光Merge图像;D,BmN转染EGFP-BmChd64质粒后绿色荧光图像;E,BmN转染EGFP-BmChd64质粒后白光图像;F,BmN转染EGFP-BmChd64质粒后荧光和白光Merge图像;N,细胞核;C,细胞质;绿色荧光,目的蛋白存在的位置。Note:A,Green fluorescence image after transfection of EGFP vector in BmN cells;B,White light image after transfection of EGFP vector in BmN cells;C,Fluorescence and white light Merge image after transfection of EGFP vector in BmN cells;D,Green fluorescence image after transfection of EGFP-BmChd64 plasmid in BmN cells;E,White light image after transfection of EGFP-BmChd64 plasmid in BmN cells;F,Fluorescence and white light Merge image after transfection of EGFP-BmChd64 plasmid in BmN cells;N,Nuclei;C,Cytoplasm.The fluorescence presented the location of target gene.
3 结论与讨论
本研究成功克隆了家蚕的BmChd64基因,该基因cDNA的开放阅读框为567 bp,编码188个氨基酸,预测蛋白分子量大小为20.9 kDa,理论等电点为8.41,存在单个CH结构域,与其它昆虫中已报道的Chd64蛋白含CH结构域一致(Lietal.,2007;Liuetal.,2011;Kozowskaetal.,2014;Tarczewskaetal.,2015)。同源性比对表明该基因编码的氨基酸序列与赤拟谷盗TcChd64和果蝇DmChd64同源性较高,推测BmChd64与TcChd64和DmChd64的功能相似。DmChd64的生物学功能已有研究证实,其通过与EcR/USP和Met等蛋白结合,共同调控JH应答基因的表达,在介导果蝇JH和20E激素信号交流过程中发挥了关键作用(Lietal.,2007)。赤拟谷盗TcChd64蛋白的功能尚未有报道,Kozowska等对TcChd64蛋白序列进行分析发现,其与DmChd64的相似性达74%,推测其与DmChd64的功能相似(Kozowskaetal.,2014)。因此,本研究推测,BmChd64可能与DmChd64一样,参与了家蚕JH和20E的激素调控过程。
同时,本研究利用qRT-PCR检测了BmChd64在家蚕不同组织中的表达。结果显示,BmChd64在5龄游走期的多个组织都有一定量的表达,其中BmChd64在翅原基中表达量最高,其次是表皮和中肠,表达量最低的是脂肪体,该基因在翅原基中表达量最高,表明该基因在翅原基的发育过程中有着重要的作用。在翅原基中,BmChd64在 5龄中后期的表达水平较低,而到了5龄末期和游走期表达水平较高,在蛹初期表达下降,到蛹期4 d表达又升高,至蛹末期表达量最高,这一变化趋势与20E在家蚕体内的滴度变化(杨鑫华,2010;Wangetal.,2018)趋势相吻合。在棉铃虫中,20E诱导HaCal的快速磷酸化,HaCal可通过20E或JH信号迅速转移到细胞核中与USP1相互作用,同时HaCalRNAi可以抑制20E对USP1、PKC和HR3的诱导(Liuetal.,2011)。因此推测,BmChd64可能在20E的作用下大量表达,进而导致翅原基完成幼虫-蛹期的变态发育并促进5龄中后期的蛹翅向成虫翅转变。
研究蛋白质的亚细胞定位有助于更好地了解目的蛋白的作用机理和调控机制。本研究用构建好的BmChd64-EGFP转染家蚕BmN细胞株后通过共聚焦显微镜检测荧光信号,荧光信号在细胞核和细胞质中都有分布,但主要是分布在细胞核中。Liu等发现在棉铃虫中,含有CH结构域的蛋白HaCal磷酸化能迅速进入细胞核内与核内蜕皮激素受体复合物EcR-USP结合,从而在20E和JH的通路网络中发挥重要作用(Liuetal.,2011)。因此推测家蚕BmChd64蛋白可能也与HaCal一样会被磷酸化,而磷酸化影响了蛋白的亚细胞定位。Kozowska等利用Kratky图表明果蝇DmChd64和赤拟谷盗TcChd64蛋白很可能在两个末端都有内在的无序区域(intrinsically disordered regions,IDRs),这些区域可作为与各种因子进行多重相互作用的平台,并为其调节功能奠定基础(Kozowskaetal.,2014)。BmChd64与DmChd64、TcChd64同源性最高,推测家蚕BmChd64蛋白可能也可与其它因子互作,而与其它因子的互作也会影响BmChd64的亚细胞定位。综上所述,家蚕BmChd64可能在20E的作用下参与调控家蚕翅原基的变态发育。
