四川省猪流行性腹泻病毒SNJ-P 株的分离鉴定与遗传进化分析*
2022-09-22庞茂楠蒋子睿姚学萍杨泽晓
庞茂楠,蒋子睿,王 印 ,罗 燕,姚学萍,杨泽晓
(1.四川农业大学 动物医学院,四川 成都 611130;2.动物疫病与人类健康四川省重点实验室,四川 成都 611130)
猪流行性腹泻病毒(Porcine epidemic diarrhea virus,PEDV)是一种猪肠道致病性冠状病毒,其引起的猪流行性腹泻是一种以严重呕吐、腹泻和脱水为特征的传染性疾病[1]。该病毒可通过消化道和呼吸道传播;能感染各年龄段的猪,对1 周龄内的仔猪危害最为严重,可造成100%死亡[2]。临床上,PEDV 多与猪传染性胃肠炎病毒、猪轮状病毒和猪圆环病毒等病毒混合感染[3]。因此,该病在许多国家造成了巨大的经济损失,并对全球生猪养殖业造成巨大威胁。
PEDV 属于冠状病毒科(Coronaviridae)甲型冠状病毒属(Alphacoronavirus)成员,为有囊膜单股正链RNA 病毒[4]。基因组全长约28 kb,编码4 种结构蛋白[ 纤突糖蛋白(S)、基质蛋白(M)、囊膜蛋白(E)和核蛋白(N)]、1 个辅助蛋白(ORF3)和2 个非结构蛋白 (复制酶多聚蛋白pp1a 和pp-1b)。PEDV 结构蛋白在介导病毒入侵、中和抗体产生、病毒基因组复制和转录以及病毒组装、出芽和拮抗宿主先天免疫反应中发挥着重要作用[5-7];PEDV 的非结构蛋白在病毒基因表达、RNA 合成加工以及病毒粒子组装和释放等方面发挥着重要作用[8]。
通过将PEDV S 蛋白基因进行遗传进化分析,可将PEDV 分为传统PEDV 毒株和新型PEDV 毒株[9],新型PEDV 毒株又分为S non-INDEL 型和S INDEL 型2 个亚群[10]。在生产中,S non-INDEL型导致的病死率通常更高[11]。S non-INDEL 型毒株是近年来流行的优势毒株,其基因变异较快,并引起抗原表位发生变化,是导致当前商品化疫苗免疫效果不佳的重要原因之一。本研究病料组织采集于接种过PEDV 疫苗的规模化猪场,利用该病料进行病毒分离鉴定、测定分离株的全基因序列并分析该毒株与疫苗株之间的遗传进化关系,以期为了解PEDV 流行株的变异情况及疫苗开发提供参考。
1 材料与方法
1.1 病料样本与细胞株
15 份仔猪小肠组织样本采自接种过PEDV 疫苗的四川某猪场;PEDV、TGEV (猪传染性胃肠炎病毒)和PoRV (猪轮状病毒)阳性血清以及猪肾细胞(PK-15)均由四川农业大学动检实验室保存。
1.2 主要试剂及试验动物
细胞培养液(DMEM)、胎牛血清(FBS)、胰蛋白酶和氨苄青霉素(Amp)等购自美国Gibco 公司;二甲基亚砜(DMSO)、低熔点琼脂糖、Hanks液(10×)和DL2000 Marker 购自上海生工生物工程有限公司;病毒RNA 提取试剂盒购自北京天根生化科技有限公司;反转录试剂盒和2×TaqMaster Mix 购自大连宝生物工程有限公司。
6 头1 周龄哺乳仔猪(未免疫)由四川农业大学合作猪场提供。
1.3 引物设计
PEDV 检测引物参考《猪流行性腹泻诊断技术》(NY/T 544—2015)[12];TGEV 和PoRV 检 测引物参考郭容利等[13]和卓秀萍[14]的研究(表1);根据PEDV CV777 株(登录号:AF353511)、CH/S株(登录号:JN547228)及DR13 株(登录号:JQ 023161)的全基因序列,运用Primer 5.0 软件设计22 对引物扩增分离株全基因组序列。以上引物均由上海生工生物工程有限公司合成。

表1 鉴别引物序列Tab.1 The identified primers sequences
1.4 组织样本PCR 鉴定
提取15 份组织样本的总RNA 并反转录成cDNA。根据《猪流行性腹泻诊断技术》(NY/T 544—2015)[12]及 郭容利等[13]和卓秀萍[14]设计的TGEV 与PoRV 检测方法进行RT-PCR 扩增,PCR产物经1%琼脂糖凝胶进行电泳,对15 份样品中PEDV 的感染情况进行分析。
1.5 病毒分离、纯化及病毒滴度测定
1.5.1 病毒分离
取PEDV 阳性小肠组织1 g 制成原代病毒液,分装后于-80 ℃保存备用。将1 mL 原代病毒液接种至状态良好的PK-15 细胞,于37 ℃孵育1 h 后吸弃病毒液,添加胰酶终质量浓度为4 μg/mL 的维持液5 mL,待细胞病变达80%~90%时,将细胞置于液氮中冻融3 次后离心取上清作为传代接种液。按上述方法进行病毒传代,并观察细胞病变情况。
1.5.2 病毒纯化
将1 mL 病毒传代接种液用DMEM 进行5 个系列的10 倍梯度稀释(10-1~10-5),后接种单层PK-15 细胞,另有一孔加1 mL 维持液作空白对照。于培养箱孵育1 h 后吸弃传代接种液,每孔添加2 mL 含1%低熔点琼脂糖的DMEM (2×)覆盖液,铺平后继续培养。当出现细胞病变时,每孔添加2 mL 含0.05%结晶紫的覆盖液,继续培养。眼观无色斑点为蚀斑,镜下观察蚀斑具体位置,用灭菌10 μL 枪头挑取单个蚀斑进行扩大培养。
为进行分离株的RT-PCR 检测,分别提取第3、7、10 代及纯化病毒的细胞上清液总RNA,反转录成cDNA 后进行PCR 鉴定。PCR 反应体系为25 μL,包括:PremixTaq12 μL,RNase Free ddH2O 10 μL,上、下游引物各0.5 μL,模板2 μL。反应条件为:95 ℃预变性3 min;94 ℃变性40 s,56 ℃退火40 s,72 ℃延伸45 s,共35 个循环;72 ℃延伸10 min。反应结束后,取7.5 μL PCR产物进行电泳。
1.5.3 病毒滴度测定
将纯化的病毒用DMEM 进行10 个系列的10 倍梯度稀释(10-1~10-10),后接种于长成单层细胞的96 孔板中,每个稀释度设8 孔,每孔接种100 μL;最后2 列接种维持液作阴性对照。将细胞继续培养,3 d 后观察并记录每孔细胞病变情况。按照Reed-Muench 法计算病毒滴度(TCID50)。
1.6 仔猪感染试验及临床症状和病理学观察
将6 头1 周龄健康哺乳仔猪(未免疫)分为试验组和对照组,每组3 头。饲养环境相同,分别隔离饲养。试验组将10 mL 纯化PEDV 病毒液拌料饲喂仔猪;对照组将10 mL 细胞培养液以同样的方式饲喂仔猪。每隔4 h 观察仔猪临床症状。将感染后死亡的仔猪剖检,观察各组织器官病理变化;取病变明显的肺脏、肝脏及小肠组织制作病理切片,利用实验室建立的方法测定各组织器官病毒载量。
1.7 分离株的全基因组测序及系统发育进化树构建
提取PEDV SNJ-P 株的总RNA 并反转录为cDNA。使用1.3 节中设计的全基因组扩增引物扩增分离株全基因序列,并回收纯化。将纯化的DNA 片段连接到pMD19-T (simple)载体,并转入DH5α 感受态细胞进行克隆扩增。经PCR 鉴定为阳性的菌液送至上海生工生物工程有限公司测序。使用MEGA 6.0 软件将获得的22 条序列进行拼接,获得分离株全基因序列。基于NCBI 上38 株国内外PEDV 毒株的S 蛋白基因序列信息,运用MegAlign 软件构建系统发育进化树,并进行遗传进化分析。以传统疫苗株CV777 为参考,分析SNJ-P 株的各基因序列及S 蛋白氨基酸序列遗传变异情况。
2 结果与分析
2.1 病料PCR 鉴定结果
由图1 可知:PEDV、TGEV 和PoRV 的阴性和阳性对照均成立,阳性对照条带大小分别为774、554 和229 bp,与预期相符。15 份样本中仅有1 份样本检测PEDV 呈阳性,且该份样本TGEV 和PoRV 检测均呈阴性。PCR 鉴定出的PEDV 阳性样本为后续毒株的分离提供了材料。

图1 病料RT-PCR 检测结果Fig.1 The results of RT-PCR detection of disease material
2.2 病毒分离、纯化及病毒滴度测定结果
由图2 可知:病毒感染后,盲传至第4 代开始出现明显的细胞病变效应(cytopathic effect,CPE);盲传至第7 代,细胞产生稳定的CPE;盲传至第10 代,细胞产生CPE 的时间提前,在接毒后约12 h 即观察到CPE 产生,24~36 h CPE 可达80%以上。细胞病变主要表现为细胞内颗粒增多、细胞圆缩脱落、大面积溶解和细胞间隙增大等。

图2 PK-15 细胞接种病毒液后各代病变情况Fig.2 The pathological changes of different generations PK-15 cells after inoculation with virus fluid
由图3 可知:接毒后12 h 细胞开始产生病变;3 d 后覆盖含有结晶紫的覆盖液,2~3 d 可以观察到蚀斑,空白对照组无蚀斑产生。
由图4 可知:第3、7、10 代以及纯化病毒液PEDV 检测均呈阳性,且条带亮度随传代数的增加而增大。这表明随传代数的增加,病毒增殖能力不断增强,不断适应培养环境。按照Reed-Muench 法计算病毒滴度(TCID50)为1×107.5/100 μL。

图4 各代病毒液RT-PCR 检测结果Fig.4 The results of RT-PCR detection in each generation of virus fluid
2.3 仔猪感染临床症状和病理变化
临床表现:试验组仔猪在感染后6 h 开始出现精神沉郁;12 h 后陆续出现腹泻症状;30~36 h后出现食欲减退、水样腹泻、脱水及黏膜苍白等症状;感染后40~48 h,3 只仔猪均死亡。对照组仔猪表现正常。
病理学观察:病死仔猪消化道病变最严重,胃和小肠内有未消化的乳糜,并发酵产生大量气体;小肠壁薄而透明,内充满黄绿色水样内容物,气味腥臭;肠系膜充血,肿胀严重;其他组织器官病变不明显,脾脏轻微血肿,底部有淤血;肺脏轻微肿胀,右肺叶出现淤血点;心脏局部水肿;肾脏与肝脏无明显病变。
取病变明显的肺脏、肝脏和小肠组织制作病理切片进行观察发现:肺泡壁增厚,细胞成分增多,肺泡出现萎陷融合现象(图5a);局部肝细胞肿胀,肝窦变窄或消失,肝索纹理不清,含铁血黄素沉积(图5b);空肠绒毛高度普遍降低,并出现绒毛萎缩、断裂和脱落等现象(图5c);十二指肠绒毛高度普遍降低,严重萎缩(图5d)。
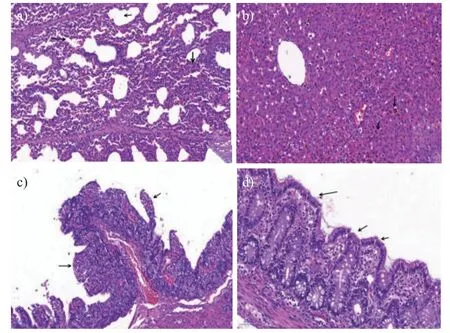
图5 病死仔猪肺脏、肝脏、空肠及十二指肠组织病理学变化(H.E,400×)Fig.5 Pathological changes of lung,liver,jejunum and duodenum in dead piglets
利用本实验室建立的qPCR 方法检测病死仔猪各组织器官的病毒载量,结果表明:肠道组织病毒载量最高,为3.82×106copies/mL;其次是肺脏,为1.47×105copies/mL;心脏、肝脏、脾脏和肾脏等组织器官病毒拷贝数均较低,分别为3.03×103、1.31×104、3.00×103和1.09×103copies/mL。该毒株具有较强的肠道嗜性,符合PEDV 感染特点。以上结果表明成功分离获得1 株PEDV 毒株,并将其命名为PEDV SNJ-P。
2.4 分离株的全基因组测序及遗传进化分析
PEDV SNJ-P 株全基因共28 003 bp,序列已上传NCBI,登录号为MK702008.1。将分离株与不同地区毒株及疫苗株的S 蛋白基因进行遗传进化分析,结果(图6)表明:分离株属于S non-INDEL 型,与同型的中国分离株PEDV-WS (登录号:KM609213.1)、CH/JLDH/2016 (登录号:MF 346935.1)和CH/HNLH/2015 (登录号:KT199103.1)亲缘关系较近,与同型的疫苗株AJ1102-F12 (登录号:MK584552.1)和LW/L (登录号:MK392335.1)亲缘关系相对较远,与传统疫苗株CV777 (登录号:LT905450.1)和DR13 (登录号:JQ023162.1)亲缘关系较远。
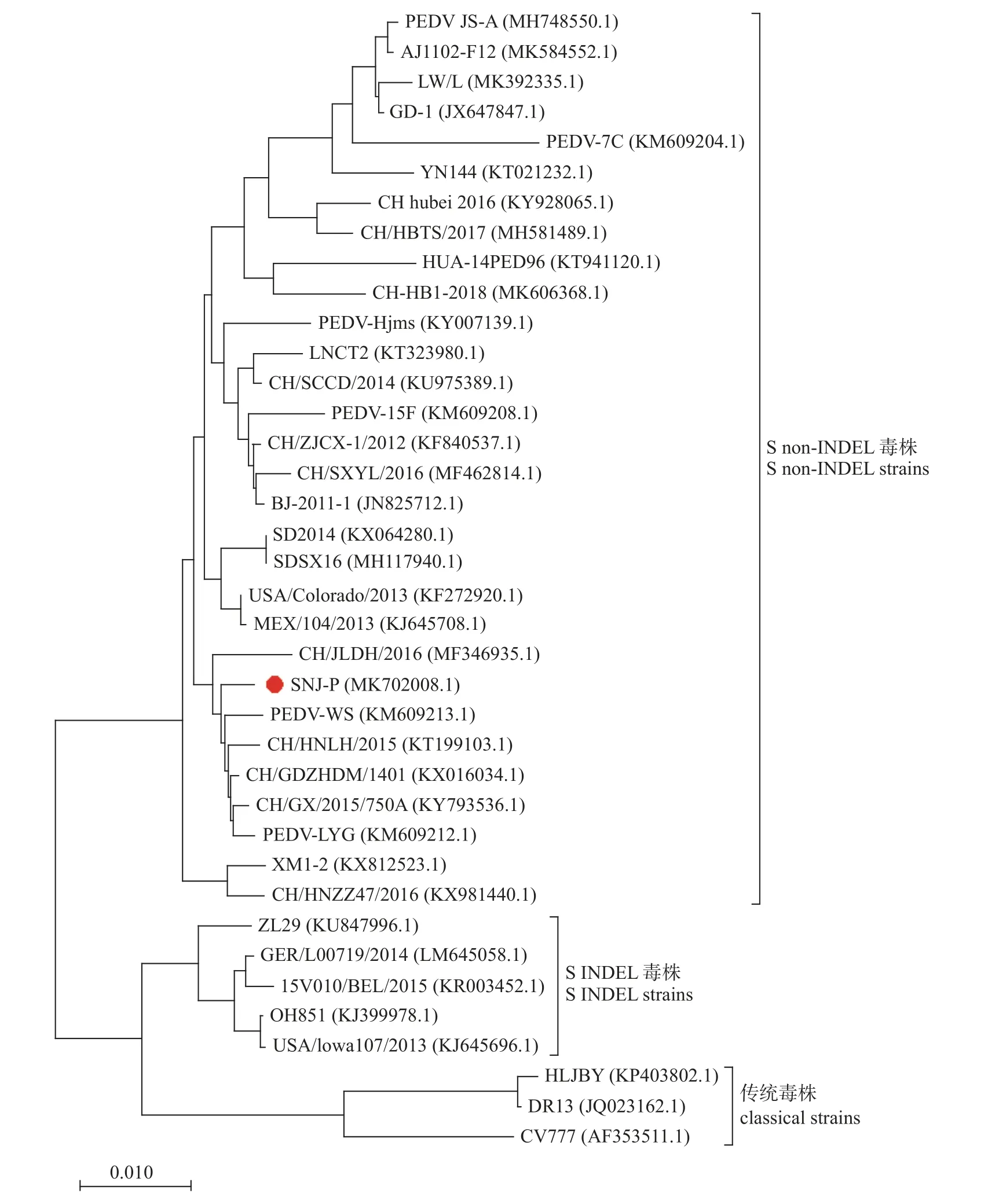
图6 基于SNJ-P 株与参考株S 蛋白基因构建的系统发育进化树Fig.6 Phylogenetic tree based on S protein gene construction of SNJ-P strain and reference strain
2.5 各序列片段遗传与变异情况
SNJ-P 株与传统疫苗CV777 的全基因序列比对结果表明:SNJ-P 株全基因序列碱基共发生980 处突变、16 处插入和11 处缺失,两者的碱基变化数如表2 所示。

表2 SNJ-P 株与CV777 株基因组碱基变化数对比Tab.2 Comparison of nucleotide changes between SNJ-P strain and CV777 strain
2.6 S 蛋白氨基酸位点变化情况
由图7 可知:SNJ-P 株S 蛋白基因共有4 161个碱基,与CV777 株相比其碱基存在较多变化,主要集中在S1 区 (1~789 aa),共发生271 处突变、6 处缺失和15 处插入。164~166 nt 插入TTG等3 个碱基,177~185 nt 插入AGGTGTTAA 等9 个碱基,415~417 nt 插入GAT 等3 个碱基,分别导致55~56 aa、59~62 aa 及139 aa 位插入氨基酸IG (异亮氨酸和甘氨酸)、QGVN (谷氨酰胺、甘氨酸、缬氨酸和天冬酰胺) 和D (天冬氨酸)。480~486 nt 缺失TGGAAA 等6 个碱基,导致160~162 aa 位缺失氨基酸DGK (天冬氨酸、甘氨酸和赖氨酸)。79~87 nt 由CAGTCTACT 突变为TCAGCTAAC,导致27~29 aa 位氨基酸由QST (谷氨酰胺、丝氨酸和苏氨酸)变为SAN (丝氨酸、丙氨酸和天冬酰胺) ;203~215 nt 由GCACAGGCATTGA 突变为CTGGCCAACATCC,导致68~72 aa 位氨基酸由GTGIE (甘氨酸、苏氨酸、甘氨酸、异亮氨酸和谷氨酸) 变为AGQHP (丙氨酸、甘氨酸、谷氨酰胺、组氨酸和脯氨酸) ;601~608 nt 位由CGCAGAAGT 突变为AGTGGAGGT,导致201~203 aa 位氨基酸由SGG (丝氨酸、甘氨酸和甘氨酸)变为RRS (精氨酸、精氨酸和丝氨酸)。其余突变位点均是由单碱基突变导致单个或2 个间断氨基酸发生改变。S 蛋白的插入与缺失均发生在S1 区,同时S1 区有63 处氨基酸位点发生突变,S2 区有19 处氨基酸位点发生突变。

图7 SNJ-P 与CV777 的S 蛋白氨基酸序列比对Fig.7 Comparison of amino acid sequence of S protein between SNJ-P and CV777
3 讨论
猪流行性腹泻病毒是引起仔猪腹泻、脱水并导致死亡的重要病原之一,在中国猪群中传播时间长、波及范围广,对生猪养殖业造成了巨大的经济损失。PEDV 毒株的分离仍然是病毒学研究及疫苗开发的重要内容。该病毒毒株在细胞增殖过程中存在细胞适应能力弱、易出现病毒滴度降解等问题[15]。因此,探索能使病毒稳定增殖的细胞系及培养条件,对于流行毒株的分离尤为重要。研究表明:PEDV 可在多数猪源细胞系中生长,包括猪膀胱细胞[16]、猪肾细胞系[17]、猪睾丸细胞系[18]、猪肺泡巨噬细胞系[19]以及猪小肠上皮细胞系[20]等,但目前猪源细胞系分离PEDV 流行株的培养条件仍需进一步探索。1988 年,HOFFMAN通过在Vero 细胞培养基中加入胰酶成功分离获得1 株PEDV 毒株;随着学者对PEDV 更加深入的研究,Vero 细胞成为分离PEDV 最常用的细胞种类,相关的试验方法也趋于完善,但Vero 细胞系分离PEDV 毒株存在增殖不稳定、滴度易降低等问题[20]。PK-15 细胞具有生长速度快、环境耐受力强等特点,且可以用于体外分离PEDV 毒株[21],但相关记录较少,试验方法并不详尽。因此,本研究尝试建立以PK-15 细胞为基础的毒株分离条件,利用PK-15 细胞进行病毒分离,并在其维持培养基中添加终质量浓度为4 μg/mL 的胰酶,盲传至第4 代即产生明显的病变,传至第7 代后产生稳定CPE。
PEDV 能感染各年龄段的猪,尤以1 周龄内的仔猪病死率最高。究其原因,一方面与初生仔猪肠绒毛更新修复能力较弱有关;另一方面与初生仔猪免疫系统不健全、抵抗病原入侵的能力较弱有关[22]。本研究利用SNJ-P 株进行动物感染试验,结果显示:仔猪出现呕吐、腹泻和脱水等典型临床症状。病毒载量检测结果显示:小肠病毒载量最高,表明此分离毒株具有较高的肠道组织嗜性。以上研究结果与周书亭等[23]和卓秀萍等[24]的研究结果基本一致。
将分离株与不同地区毒株及疫苗株的S 蛋白基因进行遗传进化分析,结果表明:分离株属于S non-INDEL 型,与同型的中国分离株PEDV-WS、CH/JLDH/2016 和CH/HNLH/2015 亲缘关系较近,与同型的疫苗株AJ1102-F12 和LW/L 亲缘关系相对较远,与传统疫苗株CV777 和DR13 亲缘关系较远。通过对SNJ-P 株与CV777 的S 蛋白氨基酸序列进行比对发现:与CV777 相比,SNJ-P株S 蛋白S1 亚基存在多处氨基酸突变、插入和缺失等变化,导致SNJ-P 株相应的抗原表位发生改变。以上结果从遗传进化层面说明传统毒株制备的疫苗免疫效果不佳,可能与当前流行毒株基因组变异较快有关,从而导致相应抗原表位发生改变。
目前,针对猪流行性腹泻尚无特效药,感染后只能通过隔离猪群或紧急免疫降低猪群的死亡率。疫苗免疫仍然是预防该病的主要措施,现有的疫苗主要以灭活苗和弱毒苗为主[25]。在中国,早在1999 年就开发出针对PEDV 经典毒株的疫苗[26];近年来,针对S non-INDEI 型毒株的商品化疫苗不断上市,如基于AJ1102 株的PEDV 和TGEV 二价灭活疫苗等[27],为PEDV 的防控提供了一定的帮助。由于病毒变异较快,现有的疫苗保护力有限,一些规模化猪场通过制备自家组织灭活苗,免疫母猪产生抗体,仔猪通过吮吸初乳获得抗体产生免疫保护,在一定程度上提高了仔猪存活率。但自家组织灭活苗同样具有一定的局限性,如:仅针对本场常发毒株,不适合大规模推广应用;仅在疫情暴发后才能制备使用;疫苗制备过程不规范,易使病毒灭活不完全,导致猪场长期带毒,加大病毒传播风险等。
4 结论
本研究利用PK-15 细胞从疑似病料中成功分离获得PEDV SNJ-P 株,该毒株与传统疫苗株和S non-INDEI 型疫苗株亲缘关系均较远,说明当前PEDV 毒株变异速度快,商品化疫苗无法提供有效保护。
