快慢羽鸡的翼羽长度与相关基因表达分析
2022-09-14白少川张乐超杜小龙郭艳丽高启萌王德贺李兰会
白少川,张乐超,杜小龙,郭艳丽,王 涛,高启萌,王德贺,李兰会
(河北农业大学 动物科技学院,河北 保定 071001)
鸡快羽和慢羽表型与性别相关,并由一对等位基因(K 和k+) 控制[1]。家禽生产中,以主翼羽长于覆主翼羽2 mm 以上为快羽,其他情况为慢羽,作为羽型判断标准应用于雏鸡性别鉴定[2],不仅节省人力,还能避免对雏鸡造成伤害,在我国地方鸡自别雌雄配套系生产中广泛应用[3-5]。
K 基因的大片段重复由PRLR和SPEF2基因的部分重复dPRLR-dSPEF2融合基因构成[6-7],PRLR和dPRLR反向定位在Z 染色体上,分别与dSPEF2和SPEF2的5´ 末端以“头碰头”方式连接[8]。禽PRLR 蛋白存在2 个跨膜域[9],与催乳素(PRL)结合后,可以激活JAK2-STAT5 信号通路,从而控制靶标基因的表达。SPEF2基因主要表达于精子鞭毛中,在精子尾部发育中有至关重要的作用,RNA-seq 测序发现该基因在卵巢中也有表达[10],表明在没有纤毛结构的卵巢中该基因可能仍然具有功能。
骨形态发生蛋白(BMPs)对成骨细胞增殖分化、促进骨折愈合有积极作用[11],除此之外BMP 可以抑制毛囊的生长,维持毛囊休止期,且在毛囊发育的不同时期表达量不同[12-13],其家族成员BMP2与Shh(Sonic hedge hog)参与调控羽毛的分支[14]。牦牛BMP2和BMPR-IA能够维持毛囊处于休止状态,Noggin 阻断剂抑制BMP2的表达,使毛囊由休止期过渡到生长期[15]。卵泡抑制素(FST)作为一种单链糖蛋白对转化生长因子β 超家族许多成员如肌肉抑素和BMP 等具有拮抗作用,过表达FST 小鼠的肌肉重是野生小鼠的2 ~3 倍[16],FST 可通过与BMP2 结合抑制其发挥作用,阻断下游信号转导,从而影响毛囊发育[17]。
坝上长尾鸡和太行鸡作为河北省优质地方鸡品种分别于2005 年和2015 年通过国家品种遗传资源委员会鉴定,被收入《国家畜禽遗传资源品种名录》。两品种鸡存在丰富的羽色遗传资源[18],此外太行鸡ev21病毒占位区上游278 bp C>T 的T 突变型与ev21整合紧密连锁[19],但在羽毛形成过程中主翼羽和覆主翼羽长度变化以及羽型相关基因的表达情况未见报道。
鸡毛囊在16 胚龄时形态发生基本完成,之后绒羽逐渐形成,最终在胚胎期结束时羽型出现明显的区别。本研究以坝上长尾鸡和太行鸡为研究对象,测量19E 和21E 坝上长尾鸡和太行鸡主翼羽和覆主翼羽的长度,采用RT-qPCR 检测鸡胚翅羽毛囊中PRLR、SPEF2、BMP2和FST 基因的表达变化,以期探究基因表达对羽型形成的影响,为明确羽型形成的分子调控机制提供理论参考。
1 材料和方法
1.1 试验材料
河北农业大学孵化室采集19E 和21E 坝上长尾鸡鸡胚各16 枚,64 枚19E 和58 枚21E 太行鸡鸡胚毛囊、肝脏、主翼羽和覆主翼羽;北京全式金公司试剂:EasyPure® Genomic DNA Kit、RNAhold®、EasyPure® RNA Kit、Trans2K® Plus DNA Marker;保定康为世纪公司试剂:Super GelRed 和2xES Taq MasterMix(Dye); 大连宝生物公司试剂:PrimeScriptTMRT reagent Kit with gDNA Eraser;苏州金唯智公司合成引物。
1.2 方法
1.2.1 主翼羽和覆主翼羽的羽长测量 采集坝上长尾鸡和太行鸡右翅第4 和第5 根主翼羽和覆主翼羽,游标卡尺量测羽长(mm),将第4 和第5 根羽长的平均值作为羽长数据。
1.2.2 DNA 和RNA 提取及cDNA 的合成 按试剂盒使用说明提取坝上长尾鸡和太行鸡毛囊总RNA 和肝脏DNA,电泳检测DNA 和RNA 的完整性;将RNA 反转录合成cDNA,使用鸡内参引物β-action检验反转录成功与否。
1.2.3 坝上长尾鸡和太行鸡的羽型鉴定 羽型鉴定参考张秀玲[20]的双重PCR 灰度值法。PCR 反应体系:DNA 模板1 μL,上下游引物各0.5 μL,2×ES Taq MasterMix 5 μL,ddH2O 补齐10 μL。PCR 反应程序:95 ℃ 5 min,95 ℃ 30 s,60 ℃ 30 s,延伸72 ℃ 40 s,24个循环,72 ℃ 5 min。
1.2.4 RT-qPCR 引物设计 根据NCBI GenBank数据库中鸡PRLR(NM_204854.1)、SPEF2(XM_015277513.1)、BMP2(NM_204358.1)、FST(NM_205200.1)、β-action(L08165.1)序列,利用Primer Premier 5.0 设计引物,引物要求跨外显子,并通过NCBI Primer Blast 检验引物特异性。试验所用引物见表1。
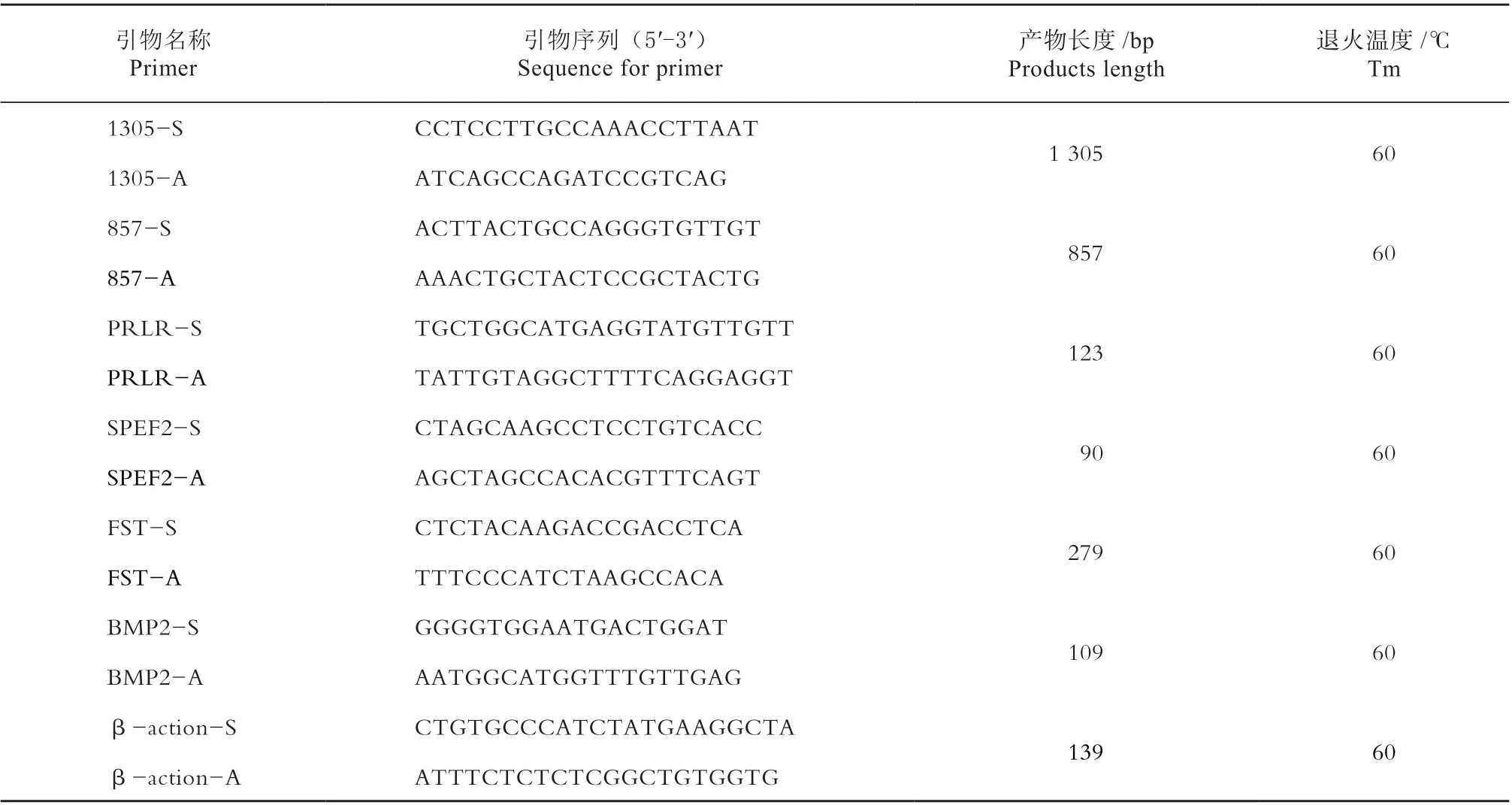
表1 试验用引物Table 1 Primers used in the experiment
1.6 数据分析
采用2-△△Ct法计算各基因的相对表达量。采用SPSS 24.0 单因素方差分析两胚龄的快慢羽太行鸡和坝上长尾鸡的主翼羽和覆主翼羽长度差异和羽型相关基因表达变化,组间差异进行Duncan 多重比较,P<0.05 被认为差异显著。
2 结果与分析
2.1 坝上长尾鸡和太行鸡的羽型鉴定
电泳检测结果仅有857 bp 扩增的个体为快羽鸡,除857 bp 外还存在1 305 bp 扩增的个体为慢羽鸡,该片段由慢羽鸡特有的融合基因扩增产生(见图1)。
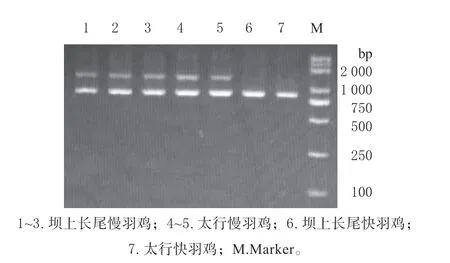
图1 羽型检测凝胶电泳图Fig.1 Gel electrophoresis images of feather type detection
不同品种鸡羽型见表2, 19E 和21E 坝上长尾鸡快慢羽各有8 枚,太行鸡19E 与21E 的快羽和慢羽型分别有22 和42、13 和45 枚。

表2 快慢羽鸡数量Table 2 Number of early- feathering and late-feathering chickens
2.2 快慢羽鸡主翼羽和覆主翼羽长度变化
快慢羽鸡主翼羽和覆主翼羽长度见表3。19E坝上长尾和太行慢羽鸡主翼羽分别长于覆主翼羽1.35 mm(P<0.05)和0.42 mm(P>0.05),太行慢羽鸡覆主翼羽显著长于坝上长尾鸡(P<0.05);另外,19E 坝上长尾快羽鸡的2 种羽长均显著短于太行快羽鸡(P<0.05),表明19E 前的坝上长尾鸡翼羽生长较慢或发育较晚,慢羽表型不明显。

表3 19 和21E 快慢羽鸡翼羽长度Table 3 Length of flight feathers for early and late feathering chickens on 19E and 21E
19E 坝上长尾慢羽鸡的主翼羽略长于快羽鸡(P>0.05),21E 时显著短于快羽鸡(P<0.05);19 和21E 的覆主翼羽均显著长于后者(P<0.05)。19 和21E 太行慢羽鸡的主翼羽均显著短于快羽鸡(P<0.05);19E 覆主翼羽略短于快羽鸡(P>0.05),21E 显著短于后者(P<0.05)。由此可知,坝上长尾鸡慢羽表型的形成在于主翼羽生长速度的降低和覆主翼羽生长速度的提高,太行慢羽鸡在19E呈现慢羽表型后,其翼羽21E 基本保持原来的生长态势。
2.3 不同胚龄两品种快慢羽鸡羽型相关基因的表达变化
PRLR在19E 坝上长尾慢羽鸡表达量是快羽鸡的1.61 倍(P<0.05),2 种羽型在21E 的表达均显著降低(P<0.05),但羽型间无显著差异(P>0.05)。19E 太行慢羽鸡的PRLR表达约为19E 快羽鸡和21E 快慢羽鸡的4 倍以上(P<0.05),后三者表达无显著差异(P>0.05),太行快羽鸡的PRLR表达未随鸡胚发育出现显著变化(P>0.05)。
SPEF2在19E 坝上长尾慢羽鸡的表达为快羽鸡的2.6 倍(P<0.05),21E 快慢羽鸡均显著升高(P<0.05),且达到相同水平(P>0.05);SPEF2在19 和21E 太行慢羽鸡表达均显著高于同胚龄的快羽鸡(P<0.05),但2 种羽型鸡的表达未随胚龄发生显著变化(P>0.05)。
BMP2在19E 坝上长尾慢羽鸡表达量显著高于21E(P<0.05),19E 和20E 慢羽鸡表达量分别为快羽鸡的4.6 倍和2.8 倍(P<0.05),而快羽鸡的表达未随胚龄出现显著差异(P>0.05)。太行鸡BMP2的表达量未随胚龄和羽型发生显著变化(P>0.05)。
FST在19E 坝上长尾慢羽鸡的表达显著低于此时快羽鸡和21E 的2 种羽型鸡(P<0.05),后三者无显著差异(P>0.05)。19E 太行快羽鸡和21E快慢羽鸡FST的表达量无显著差异(P>0.05),而19E 太行慢羽鸡表达量分别为前三者的5 倍(P<0.05)。
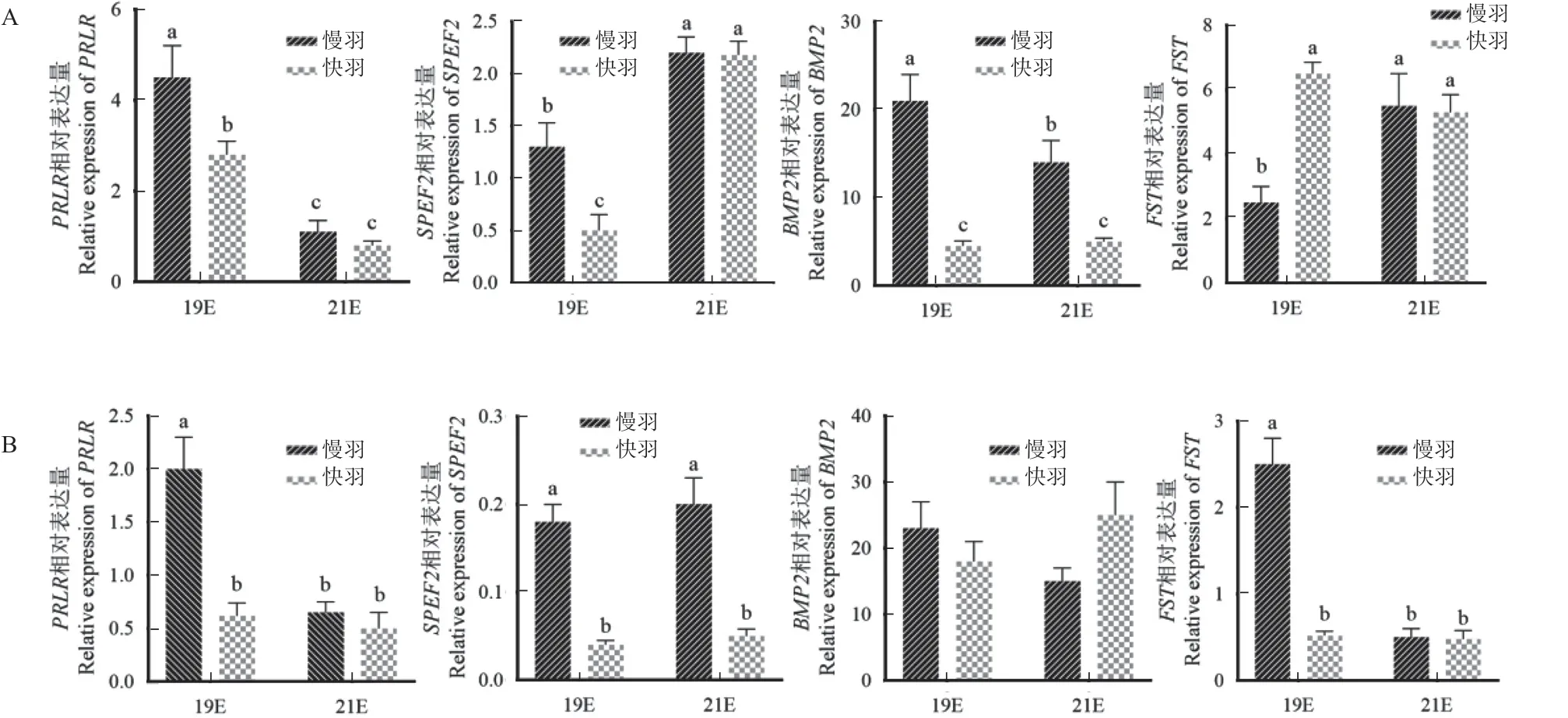
图2 坝上长尾鸡(A)和太行鸡(B)毛囊中基因的表达Fig.2 Gene expression in hair follicles of Bashang long-tailed chickens (A) and Taihang chickens (B)
2.4 不同品种快慢羽鸡相关基因的表达变化
PRLR和SPEF2在坝上长尾鸡和太行鸡上均表现出慢羽鸡表达显著高于快羽鸡(P<0.05),尤其在太行慢羽鸡表达量分别为快羽鸡的2.3 倍和4.2 倍;BMP2 在坝上长尾慢羽鸡的表达为快羽鸡的3.4 倍(P<0.05),而太行快慢羽鸡间无显著差异(P>0.05);FST 在坝上长尾慢羽鸡的表达显著低于快羽鸡(P<0.05),而在太行慢羽鸡的表达为快羽鸡的3.1 倍(P<0.05)(见图3)。
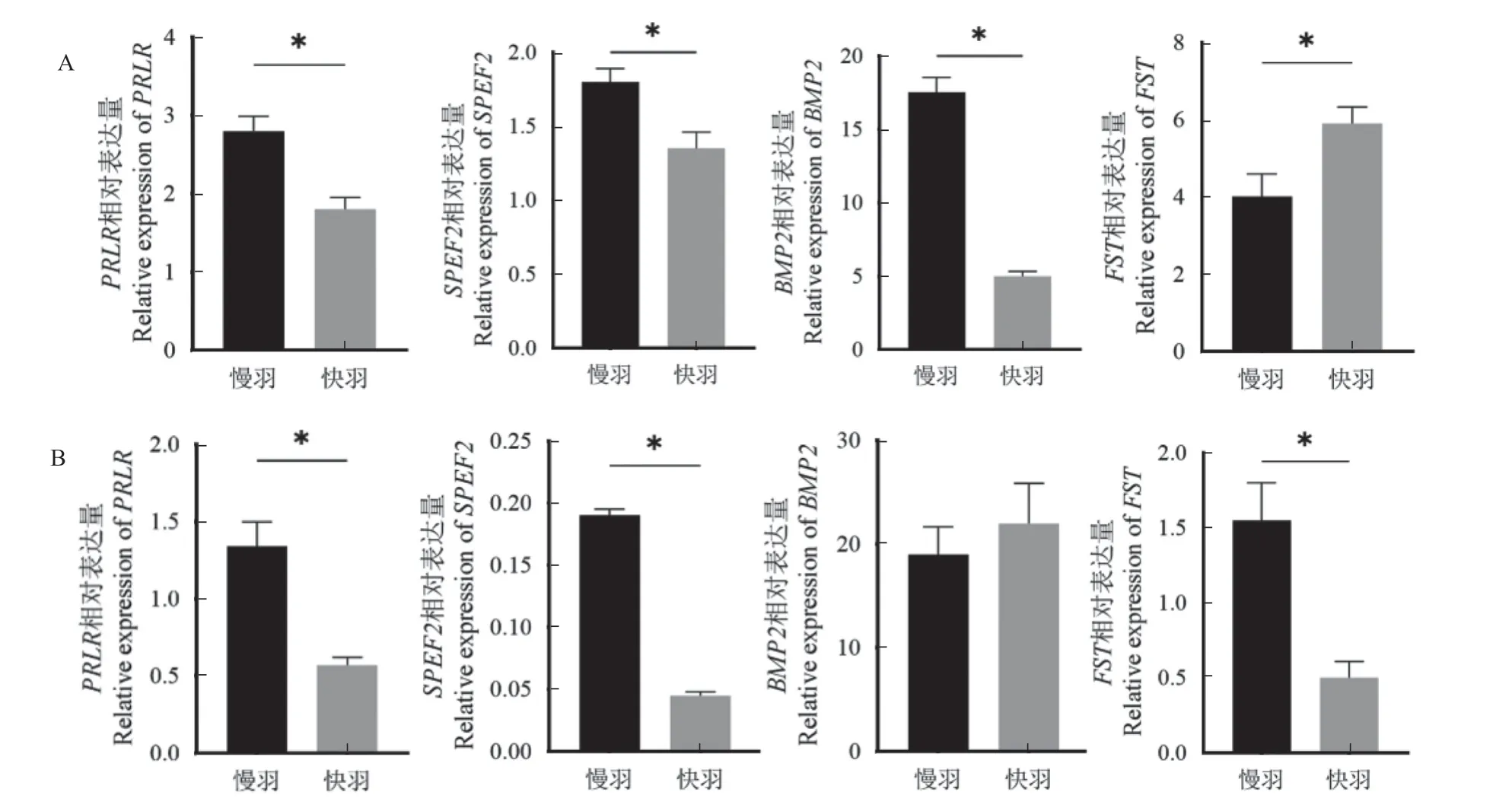
图3 坝上长尾鸡(A)和太行鸡(B)不同羽型毛囊中基因的表达Fig.3 Differences of gene expression in follicles of Bashang Long-tail chicken (A) and Taihang chicken (B)with different feather type
3 讨论与结论
3.1 快慢羽鸡主翼羽和覆主翼羽长度变化
配套系生产的商品鸡根据羽型自别雏鸡雌雄,主翼羽长于覆主翼羽2 mm 以上为快羽,其它为慢羽。娄义州等发现不同日龄武农I 系乌骨鸡主翼羽和覆主翼羽变异程度存在差异[21],赵彩娟等发现2周龄快羽鸡覆主翼羽长度显著大于慢羽鸡[22]。本试验对19E 和21E 快慢羽坝上长尾鸡和太行鸡的主翼羽和覆主翼羽长度进行测量,发现19E 前坝上长尾鸡的羽毛发育较晚或生长较慢;19-21E 坝上长尾慢羽鸡2 种翼羽生长速度快于太行慢羽鸡,但其主翼羽相对于快羽鸡生长受到抑制。坝上长尾鸡的羽型分化时间较太行鸡晚,主翼羽和覆主翼羽生长速度差异是快慢羽表型形成的原因之一。
3.2 快慢羽鸡基因表达变化
本试验发现PRLR基因在慢羽的坝上长尾和太行鸡胚翅羽毛囊中的表达均显著高于快羽鸡,这一结果与Luo 等发现文昌慢羽鸡与快羽鸡的PRLR的表达一致[23]。Craven 等发现敲除PRLR的小鼠毛发周期休止期缩短,并提前进入毛囊的生长期[24],提示PRLR在毛囊激活中起到抑制作用。白春艳等也提出PRLR对毛囊激活有抑制作用,是慢羽性状的主要候选基因[25],PRLR基因参与调控抑制慢羽鸡的羽毛生长发育。本试验对不同胚龄鸡胚翅羽毛囊PRLR的表达分析发现,19E 坝上长尾和太行慢羽鸡的表达高于快羽鸡,且高于21E 快慢羽鸡间差异。另一方面,19E 快羽的坝上长尾鸡的表达高于21E 的慢羽鸡,而19E 太行快羽鸡与21E 的太行快慢羽鸡表达无差异,这与19E 的慢羽表型坝上长尾鸡未形成而太行鸡已显现,19E 坝上长尾鸡快羽鸡主翼羽和覆主翼羽长度均短于同胚龄坝上长尾慢羽鸡和太行快羽鸡一致,表明19E 时坝上长尾快羽鸡高浓度的PRLR抑制快羽鸡毛囊的生长发育。
SPEF2由于存在ATP/GTP 结合位点和富含脯氨酸的结构域,因此被认为可能与羽毛生长通路中的信号转导有关。本试验发现除21E 坝上长尾快慢羽鸡外,SPEF2在坝上长尾和太行慢羽鸡中的表达均高于快羽鸡,Zhao 等研究表明,在1 日龄绿壳快羽和慢羽蛋鸡中SPEF2的表达一致[26],说明在鸡胚羽毛发育过程中SPEF2与PRLR协同作用发挥抑制慢羽羽毛生长的作用。本试验还发现21E 坝上长尾2 种羽型鸡的SPEF2表达均高于19E,且快慢羽间无差异,这可能与坝上长尾鸡慢羽表型分化较晚或翅羽生长速度的差异有关。
19E 和21E 坝上长尾慢羽鸡BMP2表达量均高于快羽鸡,提示BMP2可能存在抑制毛囊发育的功能,与宋亮丽发现牦牛BMP2发挥维持毛囊休止状态功能相似[15]。但在太行快慢羽鸡中各胚龄之间表达量无差异,可能太行鸡羽型分化完成早,BMP2抑制毛囊发育功能已完成。Liu 等发现FST可通过与BMP2结合抑制其发挥作用,阻断下游信号转导,从而影响毛囊发育,FST对BMP2有拮抗作用[17],试验发现19E 坝上长尾慢羽鸡FST表达量低于快羽鸡,说明慢羽鸡中FST对BMP2的拮抗作用较低,BMP2能发挥抑制毛囊发育的功能。21E 坝上长尾慢羽鸡FST显著高于19E,且与同胚龄快羽鸡之间无差异,此时羽型已经完成分化,表明高水平FST发挥对BMP2的拮抗作用,羽毛发育生长加快。同理,19E 太行慢羽鸡中FST的高表达,也是太行慢羽鸡此时羽型分化完成所致。
本试验通过测量翅羽长度和基因表达定量发现,太行鸡在19 胚龄已表现慢羽表型,而坝上长尾鸡的慢羽表型在21 胚龄才呈现;推测PRLR和SPEF2在慢羽鸡翅羽毛囊中的高表达,及BMP2和FST在太行鸡和坝上长尾鸡翅羽毛囊中的差异表达,参与鸡慢羽表型的形成。
