铜绿微囊藻对萼花臂尾轮虫与大型溞竞争关系的影响
2022-09-13李海露何雨晗
李海露 姚 慧, 何雨晗 耿 红 徐 军 何 亮 张 欢
(1. 南昌大学资源与环境学院,鄱阳湖环境与资源利用教育部重点实验室,南昌 330001;2. 中国科学院水生生物研究所,武汉 430072;3. 中南民族大学生命科学学院,武汉 430074)
在自然生态系统中,物种之间普遍存在竞争关系。竞争在一定程度上决定群落中物种组成和种群数量[1],因此是调节种群密度与群落结构的重要因子。浮游动物是湖泊生态系统中初级消费者的重要组成部分,轮虫和枝角类作为浮游动物中优势类群,在食物网中是连接初级生产者浮游植物和次级消费者的重要枢纽。轮虫和枝角类生活史特征较为相似[2],食性具有较多重叠[3],占据相似的生态位,因此,它们之间势必会引发激烈地种间竞争,引发资源限制效应[3]。
轮虫和枝角类的竞争关系已有较多研究。例如,Gilbert[4,5]发现溞通过机械性干扰和掠夺性竞争相结合的方式抑制轮虫种群增长。Macisaac等[6]研究表明枝角类通过不同的竞争方式对轮虫的抑制程度不同。在无其他环境胁迫时,枝角类在轮虫与枝角类的竞争中常占优势。轮虫和枝角类的生殖及种群增长能力对环境因子变化(如捕食压力、有毒有害物质[8])的感知灵敏,但此种感知能力存在种间差异,种间竞争优势也具有不确定性[9],因此,环境胁迫,如有毒/有害藻类暴发(或暴发程度),对轮虫与枝角类的竞争关系的影响仍知之甚少。
蓝藻水华是一个日益严重的世界性问题,蓝藻水华发生期间,藻类生物量急剧上升,可导致浮游动物群落结构改变[10,11],严重影响整个水生生态系统的结构和功能[12]。在清水稳态下,大型枝角类等大型浮游动物摄食率较高,且为滤食性生物,与小型浮游动物如轮虫相比,其摄食颗粒大,摄取效率高,因此大型枝角类类群在湖泊“清水期”占据优势地位[13],Dodson等[7]提出“大小效率假说”,认为在自然环境中由于竞争,小型浮游动物相对大型浮游动物(如枝角类)处于劣势。但是,在水体中蓝藻暴发后,其产生的有毒微囊藻会对大型溞的生长繁殖产生抑制作用[14],而小型浮游动物(如轮虫)受此影响不大,故常呈现出较高丰度[15]。那么是否因为高浓度的蓝藻抑制了枝角类种群的发展,影响大型溞与萼花臂尾轮虫竞争关系,进而导致水华后轮虫占据主要优势? 目前关注枝角类与轮虫群落结构的研究中,大多研究的是食物[16—19]、温度[19,20]、个体大小[4,21,22]及相对起始密度[23,24]等因素对枝角类与轮虫之间的竞争的影响及微囊藻毒素对单一物种影响[25—27],而微囊藻对浮游动物种间竞争的研究却较少。在蓝藻水华发生时,浮游植物群落中95%是由微囊藻组成[28],铜绿微囊藻(Microcystis aeruginosa)是最为常见的水华蓝藻[29]。因此,本实验以铜绿微囊藻为例,重点研究不同铜绿微囊藻浓度对萼花臂尾轮虫和大型溞种群竞争关系的影响。
1 材料与方法
1.1 实验材料准备
有毒铜绿微囊藻(Microcystin-producingM.aeruginosaFACHB-905)和小球藻(Chlorella pyrenoidosaFACHB-1067),均由中国科学院水生生物研究所藻种库提供,采用BG-11[30]培养液在光照培养箱中培养。培养条件为(24±1)℃,光照强度为2500 lx,周期为14h∶10h(昼∶夜)。实验取对数生长期的小球藻和铜绿微囊藻进行离心(7000 r/min,10min),离心后弃除上清液进行浓缩富集,添加少量EPA[31]培养液后放置于4℃ 冰箱中备用。实验前,采用视野计数法对铜绿微囊藻和小球藻进行计数并换算成浓度,实验时用EPA培养基稀释到实验所需浓度。
实验所用的萼花臂尾轮虫(Brachionus calyciflorus)采集自湖北省武汉下涉湖(北纬: 30°8′N,114°10E)的休眠卵孵化而得,大型溞(Daphnia magna)采集自云南省昆明市新运粮湖,各挑选一株进行单克隆繁殖,获得萼花臂尾轮虫种群与大型溞种群。两种浮游动物均采用EPA[31]培养基培养。用小球藻进行投喂。每两天更换一次培养基,取带卵的母体进行保种和扩培,使萼花臂尾轮虫种群和大型溞种群保持在指数增长阶段。实验前两种浮游动物均在(24±1)℃ 的光照培养箱中稳定培养至少3个月。
1.2 实验设计
参考蓝藻水华暴发时的浮游植物生物量(5×104cells/mL)[32],设计4组铜绿微囊藻浓度梯度(分别为0、5×104、1×105和5×105cells/mL,记为对照组、低浓度组、中浓度组和高浓度组)。每个微囊藻浓度梯度下设置3个动物处理组: 萼花臂尾轮虫单独培养,大型溞单独培养,萼花臂尾轮虫和大型溞混合培养,每组4个重复。在实验中,每个处理组除了添加对应浓度的铜绿微囊藻之外,还添加1×105cells/mL 小球藻作为食物。
在实验前,向对应的处理组中加入200 mL EPA培养液,并加入对应的萼花臂尾轮虫和大型溞。对照组与处理组的萼花臂尾轮虫和大型溞初始比例均为1∶1,且初始密度均为50 ind./L。
在观察时,将培养萼花臂尾轮虫的烧杯搅拌混匀,用移液枪吸取10 mL培养液,用浮游动物计数框计数,大型溞全部计数。实验中每48h重新添加对应的藻类,并更换一次培养基。在更换时,用孔径为50 μm的筛网滤出实验个体,转移至新的烧杯中。对照组实验至种群稳定增长为止,处理组实验至萼花臂尾轮虫或大型溞种群死亡结束。
1.3 实验参数与统计分析
种群增长率计算参考公式:r=(lnNt-lnN0)/t。N0和Nt分别指实验开始和第t天时实验动物种群密度,将种群密度取对数并与实验周期进行回归分析。各处理组种群增长率进行单因素方差分析(One-way ANOVA),用Tukey’s HSD法多重比较对不同浓度铜绿微囊藻下单独培养和混合培养的萼花臂尾轮虫及大型溞种群增长是否存在显著差异进行分析,实验参数统计分析在R软件中进行。
2 结果
2.1 低浓度铜绿微囊藻对萼花臂尾轮虫和大型溞竞争的影响
结果显示,低浓度铜绿微囊藻可增加萼花臂尾轮虫和大型溞的种群密度(图1a)。在混合培养体系中,轮虫在对照组(无微囊藻添加组)存活时间为3d,在低浓度实验组(含有低浓度铜绿微囊藻组)存活时间为5d(图2a),并且大型溞在低浓度实验组的体系中种群密度高于对照组(图2a)。该结果说明在小球藻食物浓度不足时,低浓度铜绿微囊藻能够作为食物为萼花臂尾轮虫和大型溞提供能量供给。萼花臂尾轮虫和大型溞摄食低浓度铜绿微囊藻没有改变两者种间竞争关系,大型溞仍占优势,萼花臂尾轮虫种群最后衰亡。
2.2 中浓度铜绿微囊藻对萼花臂尾轮虫和大型溞竞争的影响
单独培养和混合培养的萼花臂尾轮虫种群密度在中浓度铜绿微囊藻中都低于对照组(图1b),实验开始7d之内,对照组和中浓度实验组大型溞种群密度增长趋势不变,后期单独培养体系和混合培养体系中的大型溞种群密度均呈现增长趋势,且实验组种群密度高于对照组(图1b和2b),而在添加中浓度铜绿微囊藻的实验组中轮虫不能存活(图1b)。同时,在二者竞争体系中对照组萼花臂尾轮虫仅存活9d(图2b),由此说明中浓度的铜绿微囊藻不利于萼花臂尾轮虫增长繁殖,会导致其种群增长缓慢(图1b),反之中浓度铜绿微囊藻可作为食物促进大型溞种群密度增加(图1b),使大型溞建立有毒铜绿微囊藻的耐受性,该结果也说明大型溞对铜绿微囊藻的耐受性高于萼花臂尾轮虫。
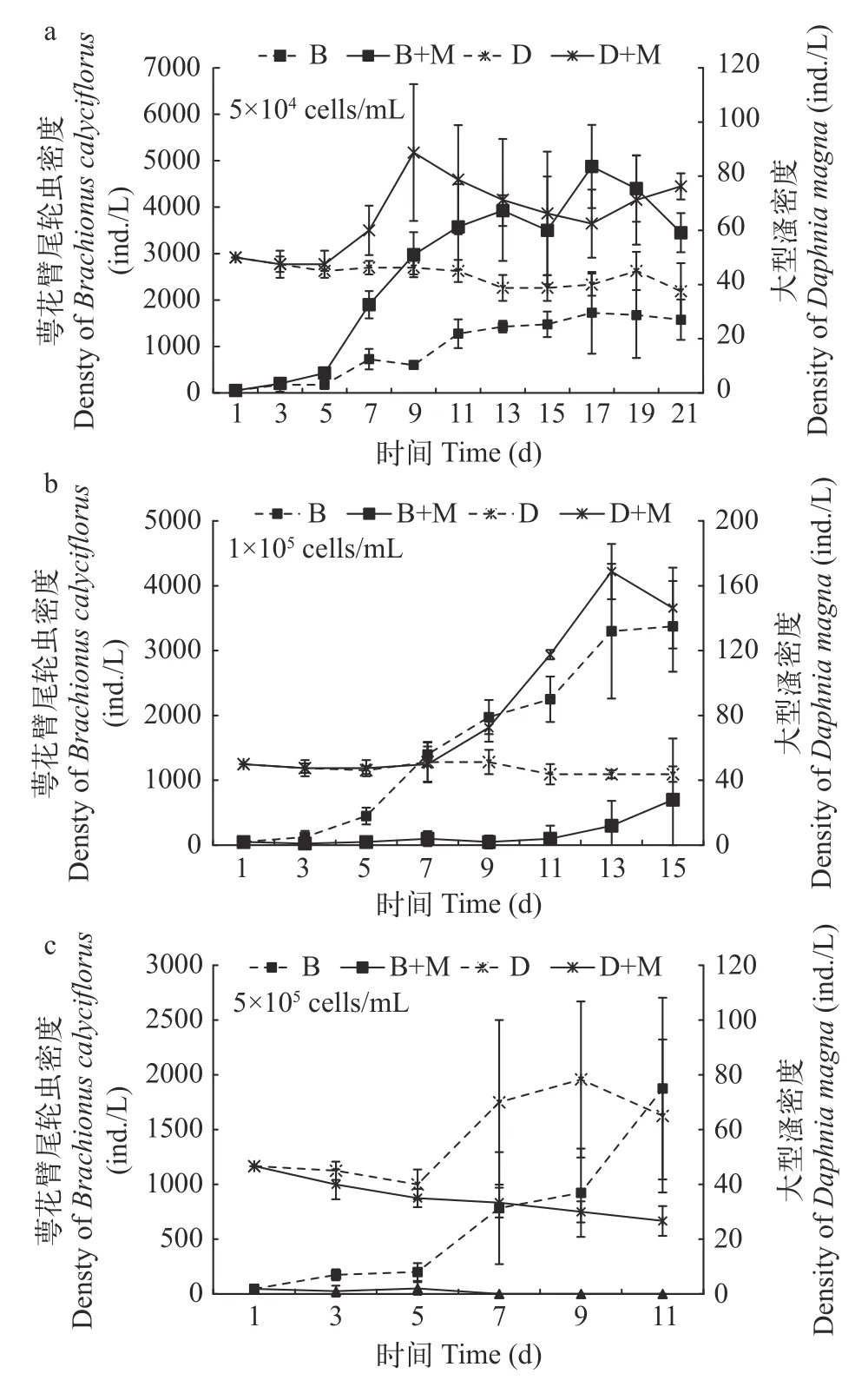
图1 不同浓度微囊藻添加对萼花臂尾轮虫和大型溞单独培养的种群数量变化Fig. 1 Different densities of Microcystis aeruginosa on the population variations of Brachionus calyciflorus and Daphnia magna in the mono-cultures
2.3 高浓度铜绿微囊藻对萼花臂尾轮虫和大型溞竞争的影响
当铜绿微囊藻浓度较高时,萼花臂尾轮虫种群在较短时间内衰退,同时大型溞种群增长受到抑制,其密度呈现下降趋势(图1c)。在种间竞争实验中,萼花臂尾轮虫仅存活5d,大型溞密度显著减少(图2c)。由此显示铜绿微囊藻处于高浓度时会使得萼花臂尾轮虫和大型溞的生长繁殖受到抑制,枝角类对有毒铜绿微囊藻建立的耐受性无法抵制摄食高浓度铜绿微囊藻毒性累积所引起的毒性效应,使得大型溞幼体不能存活,最后导致大型溞衰亡。

图2 不同浓度微囊藻添加对萼花臂尾轮虫和大型溞混合培养的种群数量变化Fig. 2 Different densities of Microcystis aeruginosa on the population variations of Brachionus calyciflorus and Daphnia magna in the mixing cultures in the control and vary level of M.aeruginosa treatment
实验总共进行11d,根据种群密度变化计算各处理组种群增长率并进行单因素方差分析,用Tukey’s HSD 法对同一物种不同处理进行多重比较。结果表明在单独培养时,萼花臂尾轮虫和大型溞种群摄食低浓度铜绿微囊藻(5×104cells/mL)和中浓度铜绿微囊藻(1×105cells/mL)都有利于增长率增加,而高浓度铜绿微囊藻(5×105cells/mL)不利于两者种群增长,萼花臂尾轮虫种群不能存活,大型溞种群增长率下降。在竞争体系中,萼花臂尾轮虫都会受大型溞抑制而衰亡,加入低浓度和中浓度铜绿微囊藻后大型溞种群相比较对照组增长率上升,而加入高浓度铜绿微囊藻在一定程度上导致大型溞种群增长率下降。这表明一定浓度范围内的铜绿微囊藻可作为食物增强大型溞种群与萼花臂尾轮虫种群的生理机能,促进增长率增加,而这种促进作用达到一定限度后铜绿微囊藻中的毒素就会影响二者的生长繁殖,间接提高种群的死亡率。同时,表1也说明,低浓度铜绿微囊藻仅对大型溞种群增长有显著性影响(P<0.01),而对萼花臂尾轮虫种群增长没有影响(P>0.05);中浓度、高浓度铜绿微囊藻对萼花臂尾轮虫和大型溞种群增长率均具有显著性影响(P<0.01)。
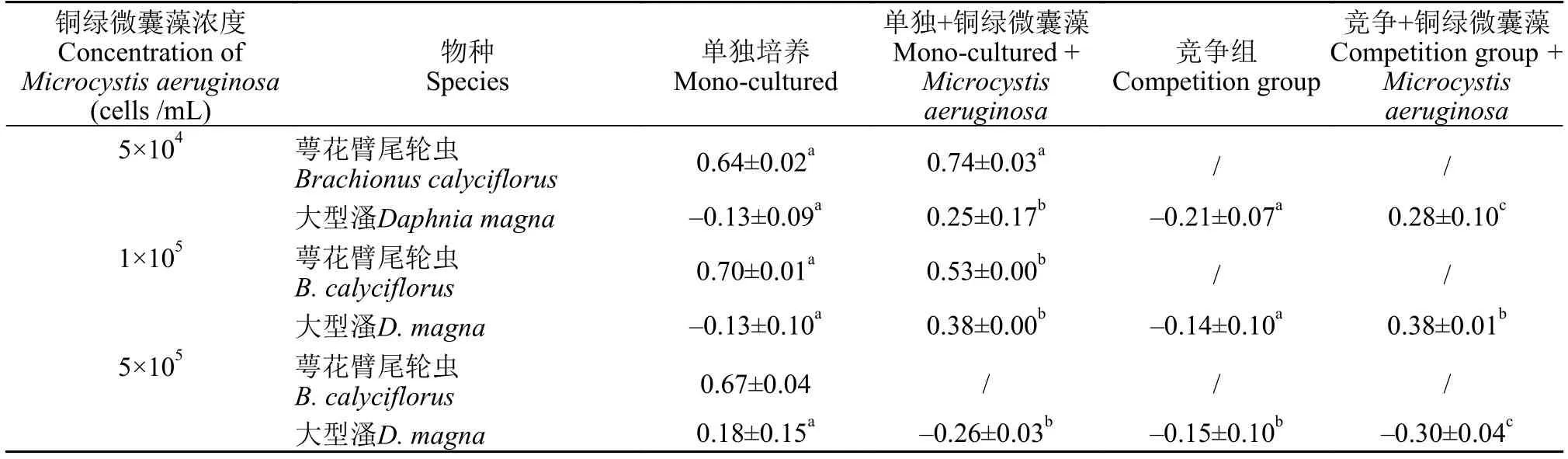
表1 实验中各处理组萼花臂尾轮虫和大型溞的种群增长率对比(平均数±标准差)Tab. 1 Comparision of population growth rate of Brachionus calyciflorus and Daphnia magna in different treatments during the experiment (mean±SD)
3 讨论
Dadson等[7]提出“大小效率假说”表明: 在同等食物浓度条件下,大型溞作为体型较大的个体,掠食能力比萼花臂尾轮虫等体型较小个体更强,萼花臂尾轮虫会被大型溞竞争性地淘汰。本研究对照组(无铜绿微囊藻组)的实验结果支持这一假说。有学者在蚤状溞与螺形龟甲轮虫同时存在的水体中加入有毒微囊藻,其毒性作用会对蚤状溞的竞争产生抑制,而有利于螺形龟甲轮虫生长繁殖[33]。在自然水体中,枝角类体型较大,滤食率比轮虫大20—1300倍[34],更容易滤食藻类。因此,相对于萼花臂尾轮虫,大型溞的生长繁殖受到有毒微囊藻的抑制作用可能更大。而本研究实验结果表明,在不同铜绿微囊藻浓度下,大型溞在二者种间竞争过程中是优势类群,萼花臂尾轮虫是劣势类群,即使二者共存于低浓度铜绿微囊藻(蓝藻暴发初始值)时,萼花臂尾轮虫也会在竞争过程中死亡。该结果说明野外蓝藻水华发生时,轮虫占优势的原因可能不仅仅是因为铜绿微囊藻浓度变化,还可能与其他环境因子相关。例如,黄林等[9]通过在温度、食物、相对起始密度等方面对轮虫与枝角类的竞争结局作出系统整理与分析,认为生态系统中各个因素都可能会对轮虫与枝角类的竞争造成影响。杨威[35]实验发现微囊藻的出现能够改变蚤状溞和长额象鼻溞之间的竞争结果,但此结果会受不同生态因子的影响。
在实验中,添加低浓度铜绿微囊藻后,与对照组相比单独培养的种群都增加;而混合培养的大型溞的增长率显著增加,萼花臂尾轮虫种群在增长后衰退更快,最后在第7天衰亡,而对照组仅存活3d。这一结果表明蓝藻水华暴发初期萼花臂尾轮虫在二者竞争中可以利用自身优势成为优势类群,但随着群落的发展,最终大型溞仍处于优势地位。尽管低浓度铜绿微囊藻在一定程度上都会促进大型溞和萼花臂尾轮虫的增长,但大型溞个体比轮虫大,根据“大小效率假说”,大型溞始终处于优势地位。明瑞梁等[36]在盔型溞与脆弱象鼻溞的竞争体系中发现,个体较大的盔型溞在不同处理中均占优势,即大型浮游动物比小型浮游动物更具优势。当食物资源有限时,大型枝角类利用其体型优势在食物竞争上抑制小型枝角类和轮虫的生长繁殖,在水体中占据优势地位[37,38]。因此,浮游动物之间的竞争结局可能不是单一因素决定,需要考虑多种因素耦合作用。
高浓度铜绿微囊藻组实验进行11d,单独培养时大型溞种群增长率下降,但未衰亡,而萼花臂尾轮虫在高浓度铜绿微囊藻组却不能存活,一方面可能是微囊藻作为内毒素随着个体对摄食食物进行消化吸收,产生毒素的过程需要累积,达到一定量后才会表达出对毒性的响应[39];另一方面,尽管轮虫对铜绿微囊藻有一定耐受性,但高浓度的微囊藻对轮虫的增长与繁殖产生高强度的抑制作用[40]。李效宇等[41]用有毒和无毒的铜绿微囊藻同时喂食大型溞种群,发现无论微囊藻是否会对大型溞产生直接毒性效应,大型溞种群生长繁殖都会受到微囊藻抑制,且认为微囊藻影响种群的消化吸收是导致种群生殖率降低的主要因素。因此,实验中尽管大型溞依旧占据主要生态位,是培养体系中的优势种群,但其种群增长率有下降趋势。一般来说轮虫作为体型较小的浮游动物,因其生长周期比较短、繁殖速率快,在种群密度上轮虫相比较枝角类而言更占优势[42],然而二者竞争结局基本不受铜绿微囊藻浓度的影响,即添加不同浓度有毒铜绿微囊藻,在混合培养体系中大型溞都占优势。
综上,实验证明即使大型溞对萼花臂尾轮虫的竞争优势受铜绿微囊藻浓度的影响不大,但是当铜绿微囊藻浓度超过一定阈值时,铜绿微囊藻会对大型溞及萼花臂尾轮虫的生长繁殖产生影响,甚至导致萼花臂尾轮虫种群衰亡。本文利用控制实验法研究铜绿微囊藻浓度对轮虫和枝角类种间竞争的影响,在自然水体中种间竞争的相互作用受到多种因素的协同作用,需要多因素实验来揭示自然水体中两类群的丰度分布格局。
