米仔兰一年生幼茎的发育解剖学研究
2022-09-09吕佳峻朱栗琼招礼军杨凤兰陈惠昕龙孟元
吕佳峻 朱栗琼 招礼军 杨凤兰 陈惠昕 龙孟元
(广西大学林学院 广西南宁 530004)
米仔兰(Aglaia odorata)是楝科(Meliaceae)米仔兰属(Aglaia)的小乔木、灌木,产于广东、广西,花黄色且芳香似兰,不仅是优良的观赏树种,还是改善血液循环、治疗跌打损伤等疾病的传统中药材[1-3]。目前学者们对于米仔兰的研究有扦插繁殖、叶绿体基因组序列、化合物的除草和抗癌作用等[2,4-6]、关于米仔兰茎的发育解剖学研究罕见报道。对植物茎进行解剖研究可为选育、栽培、分类和鉴定等提供依据[7-8],还可揭示植物的生长发育规律[9],其中导管分子形态特征对树体生长起着重要指示作用[10]。另外,学者们研究植物茎的解剖结构特征与植物抗旱、抗寒和抗虫害等抗逆性的相关性[11-14]。在茎的发育解剖学研究中,相关学者指出,茎发育过程一般包括原分生组织、初生分生组织、初生生长和次生生长4个阶段[15-16],少部分植物有三生生长[17]。茎的初生结构有表皮、皮层和维管束,不同生长时期各类细胞的比例不同,随着生长发育植物茎的各项功能逐渐完善[18-19]。自然界中双子叶植物约有20万个物种,分布广泛,具有重要的经济价值[20]。近年来关于茎的发育解剖学研究多见于单子叶植物[21-23],而双子叶植物较少。本研究拟从米仔兰一年生茎着手,按照发育顺序,研究第1~6 节茎的解剖结构和导管分子特征,探究米仔兰幼茎的发育规律及其与结构和功能相适应的方式,为双子叶植物幼茎的发育解剖学研究提供理论依据。
1 材料与方法
1.1 材料
实验材料于2021 年8 月采自广西大学校园内生长健壮的5 株米仔兰,植株为全光照下、株间相隔至少10 m。
1.2 方法
1.2.1 样品采集与处理每株采集东西南北4 个方向,每个方向采集3 条一年生枝条,总共60 条。将顶芽下第1 节定为茎的第1 节,依次往下为茎的第2、3、4、5、6 节。于节处截断,相同位置的归为一类,分别置于6 个装有70% FAA 液(70%酒精∶冰醋酸∶38%甲醛=90∶5∶5)的容器中固定48 h,待用。
1.2.2 制片
1.2.2.1 茎的石蜡切片各节随机取30 段样品的中段(长约1 cm),采用常规石蜡切片技术与叔丁醇脱水法相结合[24-25]。软化:使用冰醋酸∶过氧化氢=1 ∶1(V/V)混合溶液浸泡样品并抽真空0.5 h,处理3 h。脱水、透明:采用50%、70%、85%、95%叔丁醇依次各处理1.5 h,纯叔丁醇处理0.5 h,重复2 次。浸蜡:用50%石蜡(叔丁醇∶石蜡=1∶1,V/V)、75%石蜡(叔丁醇∶石蜡=3∶1,V/V)将脱水后的材料依次处理6 h;在纯石蜡溶液中处理12 h,期间每隔4 h 更换1 次石蜡溶液,共更换3 次。经过包埋、切片、展片、晾片后,进行以下操作:TO 脱蜡→梯度酒精复水→番红染色→梯度酒精脱水→TO 透明;用中性树胶进行封片,每段切片至少10 片;在光学显微镜下观察并拍照记录,在图像测量软件Image J 中进行数据测量,每一指标随机选取30 个重复。观察和测定指标如下:
表皮率=表皮横切面积/茎段横切面积×100%
皮层率=皮层横切面积/茎段横切面积×100%
初生韧皮部率=韧皮部横切面积/茎段横切面积×100%
初生木质部率=初生木质部横切面积/茎段横切面积×100%
髓率=髓横切面积/茎段横切面积×100%
1.2.2.2 导管离析制片按上述方法各节取6 段样品,除去表皮,浸没在冰醋酸∶30%过氧化氢=1:1(V/V)的混合离析液中,放入70℃烘箱中离析24 h;使用蒸馏水冲洗干净、番红染色,去浮色后,置于装有蒸馏水的玻璃瓶中摇散;用滴管吸取样液制成临时玻片,在光学显微镜下观察。每节制作10 个临时装片,每个取3 个视野,每个视野选取1 个典型的导管分子,总共30 个,并拍照记录。在图像测量软件Image J 中测量每个导管分子的长度、宽度、尾端长度和端壁倾斜度,其中尾端长度和端壁倾斜度测量导管两端2 个数据,取平均值。
1.2.3 数据处理与分析使用Excel 2016 对米仔兰幼茎横切结构以及导管分子特征参数等数据进行整理和作图,采用SPSS 22.0 进行数据分析,采用LSD 作多重比较。
2 结果与分析
2.1 米仔兰一年生幼茎横切结构随发育的变化
米仔兰一年生幼茎的横切结构如图1-I 所示,其横切面不规则,由外到内依次是:表皮、皮层、初生韧皮部、束中形成层、初生木质部和髓,共6 部分。由表1 可知,随着茎的发育,其直径不断变粗,各横切结构厚度均呈现上升趋势。表皮由1 层排列紧密的细胞组成,在第1~2 节茎中多数为不规则的近圆形(图1-C、1-G),发育至第3~4 节茎为近正方形(图1-K、1-O),第5~6 节茎则是长方形状的扁平细胞(图1-S、1-W),在第1~6 节茎表皮细胞最外部均形成了角质膜。皮层从第1~6 节增厚了16.46 μm(表1),靠近表皮处的5~8 层细胞为厚角组织,细胞较小、排列紧密,且含有叶绿体,可进行光合作用;靠近韧皮部6~10 层细胞为薄壁组织,细胞较大、排列疏松,因此间隙发达,便于储藏空气。在发育过程中这两部分组织的细胞层数增加不显著,但细胞体积不断增大,并且靠近韧皮部的薄壁细胞形状由第1~2 节茎的近圆形,不断被挤压成不规则的扁平状(图1-B、1-F、1-J、1-N、1-R、1-V)。
米仔兰一年生幼茎为外韧维管束,初生韧皮部在外,初生木质部在内,具有束中形成层。初生韧皮部第6 节厚度是第1 节的1.54 倍,初生韧皮部率增加了2%;初生木质部厚度第6 节是第1节的2.25 倍,初生木质部率提高了6.20%(表1)。解剖发现,各维管束形状不同、间隔不明显,髓射线较窄。随着发育,维管束的形状逐渐不易区分(图1-B、1-F、1-J、1-N、1-R、1-V)。木质部中有木射线和导管,茎横切面在有棱或凹槽处的初生木质部明显增厚,但发育至第5 节茎后,增厚的特征开始消失(图1-A、1-E、1-I、1-M、1-Q、1-U)。
髓位于茎中央,由较大的薄壁细胞组成,其直径逐渐增大,第6 节是第1 节的1.46 倍;髓率先增后减,但最终增加了1.80%(图1-A、1-E、1-I、1-M、1-Q、1-U,表1)。

表1 米仔兰一年生幼茎横切结构多重比较
2.2 米仔兰一年生幼茎导管分子随发育的变化
按次生壁加厚方式分类,米仔兰一年生幼茎导管分子类型有:环纹导管、梯纹导管、网纹导管和孔纹导管(图2),其中环纹导管极少。各类导管的比例随着发育,从第1~6 节茎,呈现由以梯纹导管为主向以网纹导管、孔纹导管为主的变化(图3)。导管分子具尾情况有:两端具尾、一端具尾一端无尾以及两端无尾,其中两端无尾在第3 节茎开始出现(如图2、4)。导管分子尾端长度先增后减,从第 1~5 节茎逐渐上升,在第1~2 节茎时增加显著,但第2~5 节茎的上升趋势均不显著,至第5~6 节茎发育时期则开始下降,且第6 节茎导管分子的尾端长度与第1 节差异不显著(表2)。
由表2 可知,在发育过程中,导管分子的长度缩短,宽度增加,二者变化趋势相反。第1~5节茎期间导管长度缩短不明显,彼此差异不显著;发育至第6 节茎时长度显著下降,与先前时期导管分子的长度呈极显著差异。导管宽度变化情况与长度类似,在第1~3 节茎宽度加大不明显,差异不显著;从第4 节茎开始呈显著加大,发育至第5~6 节茎的导管宽度与第1~4 节茎均呈极显著差异。
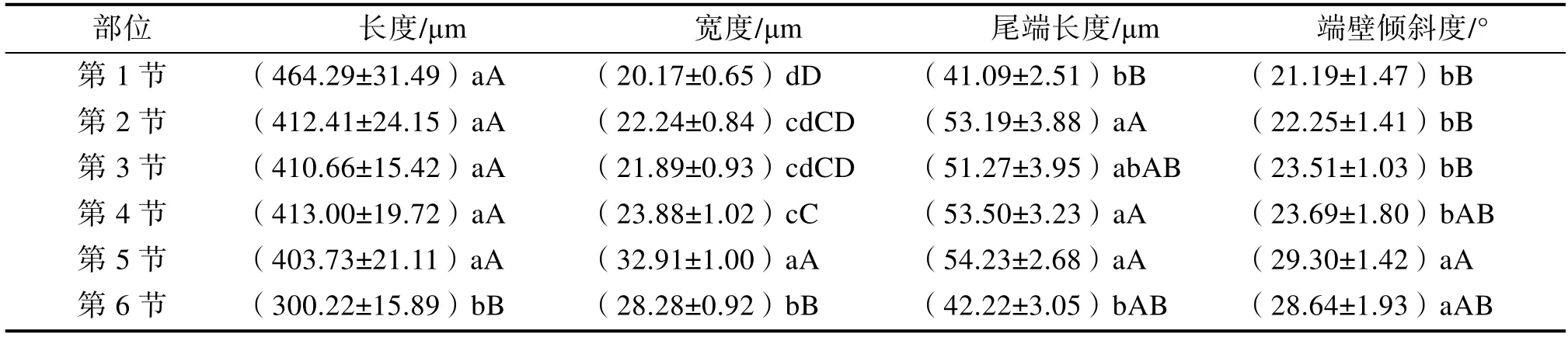
表2 米仔兰一年生幼茎导管分子特征参数多重比较
随着发育,导管分子的端壁倾斜度不断增大,但第1~4 节茎倾斜度增加趋势不显著,发育至第5 节时倾斜度显著上升。
3 讨论与结论
3.1 米仔兰一年生幼茎横切面结构组成及发育规律
米仔兰一年生幼茎横切面形状不规则,结构与一般双子叶植物相同[26]。在米仔兰第1~6 节茎的发育中表皮最外部均形成了角质膜,保护作用增强[27]。
本研究发现,米仔兰幼茎皮层厚度以及薄壁细胞体积逐渐增大,说明随着发育,位于皮层的薄壁细胞应对水分降低的能力不断提高[28],虽然皮层率呈现下降的趋势,但占比仍然在30%以上,与张敏等[29]对朝鲜越桔的研究结果类似。从第1~6 节茎,皮层薄壁细胞的形状逐渐不规则,部分被挤压成扁平状,与陈存瑞[30]对牡丹的研究结果一致。因此,米仔兰皮层发育规律与其它木本香花植物相同。
米仔兰幼茎维管束为外韧型,但髓射线较窄、束状外形不突显,与一般双子叶植物不同。初生韧皮部是植物有机物质的运送渠道[31],初生木质部是植物水分运输的关键[32]。随着发育,其初生韧皮部和初生木质部占比不断提高,输导能力逐渐增强。第1~5 节茎,初生韧皮部率大于初生木质部率,第6 节茎则相反,说明在第5~6 节茎发育时期,束中形成层的分化重心由初生韧皮部转向初生木质部,幼茎木质化速度进一步提高。
髓中的薄壁细胞体积逐渐增大,是米仔兰幼茎水分储存能力提高的表现,同时是茎直径加大的原因之一[33-34]。
3.2 米仔兰导管分子发育规律及其系统学意义
导管分子特征有:次生壁加厚方式、宽度、长度、具尾类型、尾端长度以及端壁倾斜度等[35]。本研究发现,米仔兰一年生幼茎导管分子的各项特征在个体发育过程中的演化规律不同。
3.2.1 导管类型导管分子次生壁加厚方式有5种,按照进化顺序依次是环纹导管、螺纹导管、梯纹导管、网纹导管和孔纹导管[36],次生壁木质化加厚程度增加,管腔加大,输导效率增强。环纹和螺纹导管出现在茎发育早期,梯纹、网纹和孔纹导管出现在茎发育较成熟时期[37-38]。米仔兰一年生幼茎发育后期,网纹和孔纹是次生壁加厚的主要方式。导管宽度越大,输导效率越高,但宽度加大也更容易栓塞化[39-40],降低导管分子的长度可以抵御栓塞的形成[41],同时可增强茎的机械支持力[42]。本研究中,导管长度随发育缩短,宽度则增加。
3.2.2 导管具尾及端壁倾斜状况导管具尾类型有:两端具尾、一端具尾一端无尾、两端无尾[43]。本研究中,从发育初期到发育后期,类型由两端具尾、一端具尾一端无尾2 种类型增至3 种类型,且前2 种具尾类型的比例提高。导管无尾较具尾进化,但有学者认为,尾端的形成以及伸长有利于提高水分运输效率和速率,即原始特征更利于不良环境[44];也有研究指出,同种植物导管分子的尾端长度不受环境因素的影响[45]。米仔兰导管分子的尾端长度先增后减,发育初期和后期的尾端长度最小。
导管分子端壁倾斜度的变化是导管分子的演化特征之一[46]。本研究发现,从第1~6 节茎,导管分子端壁倾斜度逐渐增大,说明米仔兰茎导管分子的端壁倾斜度特征较原始[47]。有学者认为,端壁倾斜度增大,上下两个导管分子接触的面积加大,输导效率越高[44];另有学者持不同意见,认为端壁倾斜度的增加会导致运输阻力上升,输导效率降低[38]。因此,笔者认为,不同的植物类型结果不同,要视具体情况而定,可通过测定导水率得到结论。
综上所述,米仔兰一年生幼茎的发育结构呈现一定规律,与大部分双子叶植物幼茎相似,同时结构特征与其功能性适应,保障了植株正常生长的需要。本研究从发育解剖学的角度研究米仔兰幼茎,比较了米仔兰与一般双子叶植物幼茎发育的异同,但关于楝科植物解剖学的研究鲜有,缺乏与同科植物的比较。因此,需要对其它楝科植物的幼茎进行发育解剖研究,更进一步地对其进行比较和分析,以得出更加准确的结论。
