环磷酸鸟苷-腺苷酸合成酶-干扰素基因刺激因子信号通路与非感染性炎性疾病的相关性研究进展
2022-09-02于华祥芮海英韩子琦李若川刘路路陈玉国
于华祥,芮海英,韩子琦,李若川,刘路路,薛 丽,陈玉国
山东大学齐鲁医院急诊科胸痛中心 山东省急危重症临床医学研究中心 山东大学急危重症医学研究所 山东省卫生系统急危重症医学重点实验室 山东省高等学校心肺脑复苏研究重点实验室山东省急危重症防治工程实验室,济南 250012
陈玉国 电话:18678812777,电子邮件:chen919085@sdu.edu.cn
生物体的固有免疫系统利用模式识别受体特异性识别病原相关分子模式,以检测病原体入侵及自身异常。作为胞质DNA感受器,环磷酸鸟苷-腺苷酸合成酶(cyclic GMP-AMP synthase,cGAS)可被外源DNA、损伤细胞释放的 DNA、线粒体泄漏的DNA以及胞内产生的微核活化,催化产生第二信使环鸟苷酸-腺苷酸(cyclic GMP-AMP,cGAMP),cGAMP进一步激活干扰素基因刺激因子(stimulator of interferon gene,STING),促进Ⅰ型干扰素(interferon,IFN)和干扰素刺激基因(interferon-stimulated gene,ISG)的表达,在宿主抵御外界病原菌的入侵中发挥了重要作用,同时Ⅰ型IFN和ISG(包括CXCL10、IFIT 1、IFIT3、IRF7、ISG15、CD14)作为免疫相关细胞因子,在cGAS-STING通路过度活化时与机体多种系统疾病的炎症反应密不可分。近年来,研究发现cGAS-STING信号通路与多个系统非感染性炎性疾病有密切联系,提示其在生命体中的广泛作用。本文综述了目前国内外对cGAS-STING信号通路在非感染性炎性疾病方面的研究进展。
cGAS-STING通路
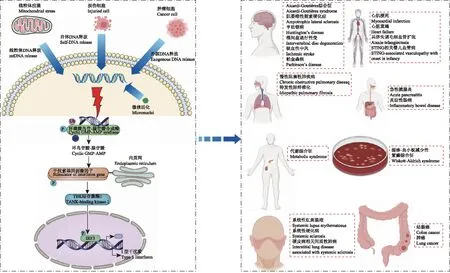
cGAS及STING的发现2013年,研究者发现当DNA暴露于哺乳动物细胞的细胞质时,在细胞质中能产生cGAMP,这种能催化产生cGAMP合成的酶,被命名为cGAS[1]。cGAS是机体内分布最广泛的细胞质 DNA 感受器。近年来发现除细胞质外,cGAS在细胞核和细胞质膜中也有分布,cGAS的不同定位对于防止cGAS对自身DNA的异常反应以及正确感知病毒感染是至关重要的。例如:位于细胞核中的cGAS的活性被核小体抑制,不能催化合成cGAMP,防止cGAS与核自身DNA发生自反应[2];cGAS的N端结构域与质膜中磷酸磷脂酰肌醇相互作用,定位在细胞膜,这一特点避免其识别基因毒性应激条件下胞质内异常自身DNA[3]。cGAS与dsDNA结合后,组装成2∶2的cGAS-dsDNA寡聚复合物,使cGAS活化[4]。活化后,cGAS的催化亚基就会发生结构重排,在GMP的2-羟基和AMP的5-磷酸之间,和在AMP的3-羟基和GMP的5磷酸之间分别形成2’- 5’和3’- 5’磷酸二酯键,合成2’3’-cGAMP。STING蛋白能够锚定于内质网或线粒体,其C端胞质结构域可通过募集相关分子而发挥重要的信号调控作用。STING是胞质内识别dsDNA并诱导产生IFN的固有免疫过程的中心分子。研究者发现cGAMP被cGAS催化合成后,作为第二信使,与STING结合,活化的STING从内质网转位至高尔基体,进而招募细胞质中的TBK1并使TBK1自磷酸化,继而诱导下游IRF3磷酸化。磷酸化的IRF3发生二聚化后入核与IFN-β启动子结合,促进Ⅰ型IFN以及其他免疫相关细胞因子的表达,发挥生物学效应[5]。
cGAS-STING信号通路的激活与炎症细菌、病毒和DNA片段等外源性DNA,外源性反转录病毒cDNA;部分外源性RNA病毒,微核,细胞碎片,内源性DNA等多种物质可以被cGAS感知,激活cGAS-STING信号通路[6]。另外,Gehrke等[7]证明在活性氧刺激或紫外线辐射时会导致鸟嘌呤氧化为8-羟基鸟嘌呤(8-hydroxy Guanine,8-OHG),造成自身DNA氧化损伤,也会引起cGAS-STING信号通路的激活。在感染的情况下,侵入细胞质中的DNA或RNA病毒、细菌等病原体被cGAS识别,激活cGAS-STING通路介导的下游炎症,炎症因子介导天然免疫应答以抵抗病原微生物感染[6]。在非感染的情况下,各种致病因素或病理刺激导致自身胞质DNA的异常释放也会激活该通路,从而导致自身无菌性炎症,参与了机体多种组织器官的非感染性炎性疾病[6]。
cGAS-STING信号通路的负性调节机制翻译后修饰的调节作用:(1)翻译后修饰对cGAS的调控:阿司匹林能在Lys384、Lys394或Lys414位点乙酰化cGAS,抑制cGAS活性,减轻Aicardi-Goutières综合征中DNA诱导的自身免疫炎症反应[8]。(2)翻译后修饰对STING的调控:蛋白磷酸酶PPM1A可使人STING蛋白S358位点去磷酸化,拮抗STING聚集,避免STING过度激活[9]。蛋白相互作用也可以调控cGAS-STING通路。cGAS同Beclin- 1相互作用,通过抑制cGAMP的产生和增强自噬降解胞质DNA,避免cGAS的过度激活[10]。可见,cGAS及STING分子的负性调节机制对于避免cGAS-STING通路的过度激活及其介导的非感染性炎性疾病的治疗至关重要。
cGAS-STING信号通路与非感染性炎性疾病
cGAS-STING通路介导的Ⅰ型IFN炎症信号反应,参与了各个系统非感染性炎性疾病的发生发展(图1)。
神经系统Aicardi-Goutières综合征是一种遗传性神经系统疾病。其由参与核酸代谢的基因突变所引起,例如trex1、RNaseH2和SAMHD1。Trex1基因敲除小鼠多种器官组织中cGAMP和炎性因子水平增加,生存期缩短[11],而cGAS或STING基因敲除可抑制此现象[12]。提示cGAS-STING通路是Trex1功能缺陷相关的自身免疫性疾病的关键机制,为其治疗提供了新的见解。
肌萎缩性侧索硬化症(amyotrophic lateral sclerosis,ALS)的发生与Ⅰ型IFN途径上调相关的神经炎症细胞因子谱和TDP- 43的细胞质积累相关。研究发现,当TDP- 43侵入线粒体并导致线粒体DNA的释放,可激活cGAS-STING通路驱动下游炎症,导致ALS患者脊髓样本中cGAS信号代谢物cGAMP水平升高[13]。
cGAS-STING通路激活介导的神经炎症在衰老与年龄相关神经系统疾病的进程中发挥了重要作用。例如,亨廷顿病小鼠的线粒体DNA释放、cGAS激活和神经炎症增加,而褪黑素作为一种由神经元线粒体合成的内源性自由基清除剂,可以逆转这一现象[14]。cGAS-STING的上调与椎间盘退行性变的发生有关,在H2O2处理的髓核细胞中抑制cGAS/STING/NLRP3通路,可发挥抗凋亡、抗炎症和提高细胞活力的作用[15]。
P:磷酸化;Ub:泛素化;Ac:乙酰化;IRF3:干扰素调节因子3
同时,cGAS-STING通路引起的神经炎症和组织损伤在缺血性中风中起重要作用,为缺血性中风的治疗开辟了新的途径。例如,小胶质细胞中cGAS的缺失可特异性降低脑组织炎症,减轻小鼠脑缺血再灌注模型的病理损伤[16]。
研究亦表明,帕金森病也与内源性DNA释放导致的IFN增加有关。在Parkin或Pink1缺失小鼠中,线粒体应激状态下导致DNA释放,激活cGAS-STING信号通路,而STING的敲除可以减轻帕金森病的炎症反应和神经变性[17]。
循环系统心肌梗死(myocardial infarction,MI)会出现坏死细胞中DNA的释放。研究发现在小鼠MI模型中,大量坏死心肌细胞中的DNA片段被心脏巨噬细胞吞噬而出现在胞质中,从而激活cGAS-STING通路,促进以IFN为主的炎症反应;与野生型小鼠相比,缺失该通路关键分子的小鼠(包括敲除cGAS、IRF3和IFNAR1)在MI模型中明显具有较高的早期存活率[18]。与此相一致的是Cao等[19]发现在小鼠MI模型中,cGAS基因缺失小鼠心脏组织梗死区中M1型巨噬细胞显著减少,M2型巨噬细胞明显增多,促进心肌组织修复,改善心室重构和提高存活率。
越来越多的证据表明,在人类和实验动物心脏衰竭中,心脏炎症引起左心室重塑和功能障碍。TAC模型小鼠左心室组织中cGAS/STING通路被激活,表现出明显的病理心脏重构和左心室功能障碍,抑制cGAS可以提高TAC后的早期存活率,保留左心室的收缩功能,并减弱包括心肌肥厚、纤维化和凋亡在内的病理重构[20]。
ATM激酶基因突变会导致共济失调毛细血管扩张(ataxia-telangiectasia,A-T),伴有多种炎症表现。研究发现在A-T患者及ATM基因缺失小鼠模型中IFN增加,但在ATM-/-STING-/-双敲小鼠中,这种炎症表型未见升高,证明ATM基因缺失造成的IFN升高与cGAS-STING通路有关[21]。
STING的异常突变也会引起一种致命的儿童自发炎症疾病,即STING相关婴儿期发病的血管病(STING-associatedvasculopathy with onset in infancy,SAVI)。SAVI是一种以全身炎症、严重皮肤血管病变、肺间质病变和复发性细菌感染为特征的常发性疾病。这种STING的异常突变导致IFN信号激活,进而导致过度炎症反应,威胁患儿生命[22]。
呼吸系统慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)是一种严重的慢性炎症性疾病,其特征是气道炎症反应增强。研究显示COPD患者外周血DNA损伤水平升高,且游离DNA水平可用于识别预后不良风险增加的COPD患者[23]。吸烟是COPD的主要原因之一,它导致氧化-抗氧化失衡、DNA损伤、细胞死亡和随后的肺部炎症。dsDNA释放后cGAS-STING通路激活触发的Ⅰ型IFN依赖的肺部炎症,在香烟诱导的肺损伤中发挥关键作用[24],可能是COPD的治疗靶点。
特发性肺纤维化(idiopathic pulmonary fibrosis,IPF)是一种原因不明的以呼吸困难和肺功能进行性恶化为特征的慢性肺纤维化性疾病。其中,矽肺是由于吸入二氧化硅或石英而引起的肺间质性疾病,特征是严重肺纤维化和慢性炎症。STING信号通路在矽肺患者的肺部被激活,表现为STING过表达、磷酸化和二聚体的形成、TBK1和IRF3的磷酸化以及CXCL10的产生,而cGAS或STING的缺失可改善硅引起的肺部炎症[25]。
另外,一项研究表明,气道上皮细胞cGAS缺失显著降低了卵白蛋白诱导的过敏性气道炎症,提示cGAS-STING信号通路参与了哮喘的发病过程[26]。
在新型冠状病毒肺炎患者中,细胞因子水平升高,其中CXCL10、CCL7在重症患者中显著增高,CXCL10是一个典型的ISG,提示1型IFN的激活。且新型冠状病毒肺炎患者血清游离DNA水平升高[27]。笔者推测新型冠状病毒感染后,由于应激或组织损伤导致的DNA释放可能激活cGAS-STING通路,导致1型IFN和炎症细胞因子的过量产生,从而加剧COVID- 19的严重程度。
消化系统在急性胰腺炎小鼠模型中,死亡腺泡细胞的DNA通过STING激活促进炎症反应,当cGAS或STING敲除后,模型小鼠中的胰腺水肿减轻、炎症和胰腺损伤的标志物下降[28]。
炎症性肠病(inflammatory bowel disease,IBD)是一类非特异性炎性疾病,持续过度激活的免疫应答是IBD的主要标志。Liu等[29]报道肠道黏膜免疫系统的树突状细胞通过cGAS-STING通路监视、识别和处理异常信号,树突状细胞对DNA的识别异常将造成免疫稳态的失衡,导致炎症性肠病的易感性增加。
内分泌系统代谢综合征是一种常见的代谢紊乱,主要表现为糖耐量异常、胰岛素抵抗、肥胖、高血压、脂质代谢紊乱、组织慢性炎症。在脂肪组织中,cGAS-STING通路依赖的慢性炎症在肥胖导致的代谢功能障碍中发挥重要作用,STING敲除可减轻肥胖引起的脂肪组织炎症、胰岛素抵抗和糖耐量受损[30]。
血液系统湿疹、血小板减少伴免疫缺陷综合征(Wiskott-Aldrich syndrome,WAS)是一种罕见的X-连锁的原发性免疫缺陷病,以血小板减少、出现湿疹、反复感染及自身免疫和恶性肿瘤发生率增加为特点。该疾病由血细胞和免疫细胞的WAS基因突变,导致WAS蛋白表达水平降低引起。cGAS/STING通路的激活有助于WAS基因突变细胞中Ⅰ型IFN的过量产生[31]。cGAS/STING通路有望成为缺乏合适供者的患者的重要治疗方案。
风湿免疫系统系统性红斑狼疮(systemic lupus erythematosus,SLE)是一种慢性炎症性自身免疫性疾病,在SLE患者血清中IFN和多种ISG的表达升高,并与SLE疾病活动相关[32]。最近研究发现SLE患者血清中IFN和ISG的表达升高与cGAS-STING通路有关[33],且通过敲除cGAS或STING,症状可以得以改善[34]。
系统性硬化病是一种多系统自身免疫性疾病,以慢性炎症和进行性皮肤和内脏纤维化为特征。硬化病相关间质性肺病患者血浆中线粒体DNA升高,cGAS-STING激活以及Ⅰ型IFN和IL- 6表达升高[35]。
肿瘤近来研究发现,氧化应激、代谢改变和基因不稳定性导致肿瘤细胞核内DNA和线粒体DNA发生损伤并释放至细胞质中促进cGAS-STING通路激活,进而促进或抑制肿瘤的发展。不同类型的肿瘤,cGAS-STING通路在其中发挥的功能不同。抑制肿瘤:cGAS-STING通路激活的肿瘤内免疫细胞募集对抗肿瘤免疫至关重要。cGAS-STING通路激活后促进免疫依赖抗原呈递细胞将肿瘤抗原提呈给CD8+T细胞,从而增强T细胞的抗肿瘤作用。例如,在小鼠结肠腺癌模型中,使用STING激活剂活化树突状细胞,促进提呈抗原给CD8+T细胞发挥抗肿瘤作用[36]。促进肿瘤:cGAS-STING信号通路持续活化介导的慢性炎症会诱发炎症所驱动的肿瘤发生。例如:在二甲基苯丙蒽诱导的皮肤癌模型中,STING信号通路激活可诱导炎性细胞因子和皮肤炎症的产生,诱发炎症驱动的上皮癌发生[37]。
cGAS-STING通路作为疾病防治新靶点
激活cGAS-STING通路分子被认为是治疗病毒感染疾病和提高抗肿瘤免疫的一种有吸引力的策略。环二核苷酸作为一种既能激活鼠又能激活人STING蛋白的激活剂,在小鼠B16黑色素瘤、4T1乳腺癌、CT26结肠癌、胰腺癌、皮肤癌以及B细胞淋巴瘤等多种类型肿瘤中,表现出明显的抗肿瘤作用和较好的应用前景[38]。但是由于cGAS-STING通路与肿瘤关系的复杂性,在上述炎症驱动的肿瘤中使用STING激动剂有可能会过度激活 STING信号通路而促进肿瘤的发展。靶向该通路的免疫治疗距离临床实践还需要更深入的研究。
除了激活该通路治疗病毒感染疾病与抗肿瘤之外,随着与cGAS-STING有关的非感染性炎性疾病进入人们的视野,研究者同样希望出现该通路的分子抑制剂。有研究发现苏拉明能够取代与cGAS 结合的DNA,阻止cGAS活化[39]。抗疟疾药物如氯喹和羟氯喹能够抑制DNA和cGAS之间的结合,阻碍通路的激活[40]。氨来占诺阻断TBK1-IRF3-IFN炎症信号通路,减轻慢性炎症反应,增加胰岛素敏感性,减少肝脏脂肪变性,改善小鼠急性心肌梗死后心功能[41],或许将来成为临床上治疗代谢性疾病及急性心肌梗死的新靶点。
总结与展望
感知胞质DNA的cGAS-STING信号通路作为先天免疫的重要组成部分,除了在抵抗病原体感染中发挥重要作用,在多个系统的多种非感染性炎性疾病中的重要作用逐渐被人们所认识,提示此通路更广阔的研究前景,给多种疾病寻求治疗靶点提供了可能。但此信号通路参与的多种生理病理过程仍有待进一步的探究。随着对这一通路的持续探索,未来可能会有与人类健康和疾病相关的新的重大发现。
