尖叶匐灯藓对不同栽培基质的生长和生理响应
2022-09-01史秉洋王秀荣陈洪梅杨铄渊
史秉洋,王秀荣,陈洪梅,杨铄渊
(贵州大学 林学院,贵阳 550025)
尖叶匐灯藓[Plagiomniumacutum(Lindb.) T. Kop],属真藓目提灯藓科匐灯藓属,因其植株形态美观[1]、覆盖能力强且具有药用价值[2],拥有广阔的应用市场和前景。目前关于尖叶匐灯藓的繁殖栽培研究较少,巨大的市场需求与匮乏的繁殖栽培技术造成对野生尖叶匐灯藓过度采摘,严重破环自然资源与生态环境。因此,快速有效的繁殖栽培技术是解决尖叶匐灯藓市场需求的当务之急。
苔藓栽培大多采用传统的土壤基质,繁殖速率较低,不能快速满足市场应用的需求,因此寻找一种适于苔藓植物生长的成本低、绿色环保的栽培基质是苔藓植物产业化栽培的关键。研究表明森林倒木上苔藓植物生长较好[3],倒木基质会影响苔藓植物N和P的含量[4],苔藓植物多样性及其组合空间结构的复杂程度随着倒木的腐解进程逐渐增加[5],其配子体的生长和生活性状也受倒木腐解类型的影响[6-7]。因此,倒木对苔藓生长的影响为寻找新型苔藓栽培基质提供了新的思路,但倒木是否可以作为栽培基质且更有效促进苔藓植物的生长,以及其影响苔藓植物生长的机理是什么,都有待进一步考证。鉴于此,本研究以尖叶匐灯藓作为栽培材料,以森林中常见的Ⅱ、Ⅲ、Ⅳ腐解等级马尾松倒木和常见的栽培土壤作为栽培基质,探讨其对尖叶匐灯藓生长和生理指标的影响以及影响的关键因子,以期为苔藓繁殖栽培提供新的途径和思路。
1 材料和方法
1.1 试验地概况
试验地位于贵州省贵阳市花溪区贵州大学西校区(106°65′~107°17′E,26°45′~27°22′N),海拔高度为 1 137.76 m,属于亚热带湿润温和型气候,兼有高原性和季风性气候特点。年平均气温为 15.3 ℃,年平均相对湿度为77%,年平均总降水量为 1 129.5 mm,年平均日照时数为1 148.3 h。试验期间于每天8:00-9:00、12:00-13:00和17:00-18:00用温湿度计记录环境温度、湿度变化(图1)。苔藓喜阴湿环境,且有文献报道尖叶匐灯藓的适宜光照范围为遮光率70% ~ 90%[8],故栽培区域做3层遮阳网处理,保证遮光率在70%以上。根据实际情况进行喷雾保湿,见干浇水,不干不浇,并定期清除杂草。
1.2 试验材料
参试尖叶匐灯藓采摘于贵州省贵阳市观山湖公园。土壤基质选择苔藓栽培常用的颗粒土、通用营养土以及草炭土;马尾松倒木采集于贵州省贵阳市贵州大学西校区马尾松林,参照闫恩荣等[9]、马豪霞等[10]和刘凌等[5]对森林粗木质残体腐解等级的划分及判定标准,确定Ⅱ、Ⅲ、Ⅳ级倒木的腐解等级(表1),采用树皮、边材和心材混合取样。

表1 倒木等级分类表Table 1 Fallen logs classification table
1.3 试验设计
分别将Ⅱ、Ⅲ、Ⅳ腐解等级的马尾松倒木粉碎混匀,分别制成WⅡ、WⅢ、WⅣ3种倒木基质,再加上常用的颗粒土(SG)、通用营养土(SN)以及草炭土(SC),试验共设置6种栽培基质。取相同体积的6种栽培基质均匀平铺于垫有无纺布的种植盆中,移栽前用水浇透。借鉴孙俊峰的混合播种法(面积法+重量法)[11],用粉碎机把苔藓植物配子体粉碎后称取1.40 g,然后以4 cm×4 cm面积均匀平铺于基质上,每个处理重复3次。
1.4 测定指标及方法
1.4.1 栽培基质指标栽培前对各基质理化性质和养分指标进行测定。基质的理化性质指标借鉴李耀龙等[12]、赵婧等[13]的测定方法,主要包括容重、持水能力、总孔隙度、通气孔隙、持水孔隙、大小孔隙比。栽培基质养分性质指标包括全氮、全磷和全钾。采用H2SO4-HClO4消煮法制备待测液,CleverChem全自动间断化学分析仪测定全氮含量,钼黄比色法测定全磷含量[14],采用火焰光度计法测定全钾含量[14]。
1.4.2 尖叶匐灯藓生长指标(1)生长长度测定:栽培30 d后,每隔10 d在每个处理中随机挑选10株苔藓测量其生长长度(精度0.1 cm)。(2)生长盖度测定:栽培30 d后,每隔10 d拍照,之后用AutoCAD2016描图测量苔藓面积,并用下面公式计算苔藓盖度。
C=(A/B)×100%
式中:C表示苔藓盖度,A表示苔藓面积,B表示种植区面积。
1.4.3 尖叶匐灯藓生理指标参考包维楷等[15]和王学奎等[16]的方法测定尖叶匐灯藓叶绿素含量,稍有改动。在不同栽培基质处理下生长120 d后,将尖叶匐灯藓清洗干净并将水分沥干,称取0.2 g置于10 mL离心管中,加入8 mL 95%的乙醇,黑暗条件下浸提24 h后,过滤并定容至25 mL。然后,以95%的乙醇作为空白对照,用分光光度计测定665和649 nm波长下的吸光度。最后根据Arnon法计算叶绿素a含量、叶绿素 b含量、叶绿素a/b和叶绿素总含量。可溶性糖含量采用蒽酮比色法测定[16],可溶性蛋白质含量采用考马斯亮蓝G-250(Bradford法)法测定[16]。
1.5 尖叶匐灯藓生长发育状况综合评价
用模糊隶属函数方法对尖叶匐灯藓生长发育状况进行综合评价。计算公式如下:
F1(Xi)=(Xi-Xmin)/(Xmax-Xmin)
F2(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)
式中:X表示第i个指标测定值,Xmin和Xmax分别表示第i个指标的测定最小值和最大值。对尖叶匐灯藓生长生理指标进行Pearson相关性分析,若该指标与尖叶匐灯藓的生长指标呈负相关关系,则采用反隶属函数公式F2计算其隶属函数值。将不同栽培基质处理下的尖叶匐灯藓生长发育指标的隶属函数值进行累加后求平均值,即得到植株形态综合评价指数,指数值越大说明该基质栽培处理下的植株生长越好[17-18]。
1.6 数据处理
采用Excel 2010和SPSS 19.0软件对数据进行单因素方差分析,用Duncan’s法(P<0.05)进行多重比较,各指标之间相关性采用Pearson相关系数进行分析,用Adobe Photoshop 2020、Excel 2010和Origin 2021进行绘图。
该项目充分体现了新时期治水思路,同时也是洪泽县水利现代化中“骨干河湖整治达标率”这一水利现代化指标实现的重要基础,具有较好的经济、社会、生态和环境效益。
2 结果与分析
2.1 不同栽培基质理化性质比较
不同栽培基质间的理化性质存在显著差异(表2)。其中,SG基质的容重显著高于其他基质,而总孔隙度和通气孔隙最低;WⅣ基质的总孔隙度和通气孔隙最高,大小孔隙比最大;WⅢ基质的持水孔隙最大,大小孔隙比最小;WⅡ基质的持水能力值最大(661.26%),而SG基质的持水能力值最低;SC基质的全氮含量最高(6.57 g·kg-1)并显著高于其他基质,SG基质的全氮含量最低,仅为0.02 g·kg-1;SG基质的全磷含量最高,并显著高于其他基质;SN基质的全钾含量最高,而WⅡ基质的含量最低。可见,倒木基质(WⅡ、WⅢ和WⅣ)的总孔隙度、持水能力以及全氮含量显著高于SG和SN基质,而SG和SN基质的容重、全磷含量和全钾含量显著高于WⅡ、WⅢ和WⅣ;而草炭土基质(SC)除总孔隙度和持水孔隙与倒木基质有显著差异,其他指标与SG和SN基质较为接近。

表2 不同栽培基质的基本理化性质Table 2 Physical and chemical properties of different culture substrates
2.2 栽培基质对尖叶匐灯藓生长的影响
2.2.1 尖叶匐灯藓生长长度尖叶匐灯藓的生长长度在不同栽培基质间有显著差异(图2,A;图3)。其中,在栽培30 d时,尖叶匐灯藓的生长长度在WⅣ基质上最长(0.72 cm),而在SG基质上最短,仅为0.36 cm;在栽培70 d后,尖叶匐灯藓生长速率在SG基质上减慢,这可能是因为在70~80 d内空气湿度经历先急剧下降后又迅速上升(图1)过程,而SG基质的持水能力又最差,从而影响尖叶匐灯藓的生长速率,与其他基质处理开始出现明显差距。在栽培90~120 d期间,所有基质处理尖叶匐灯藓生长长度在达到最大值后,生长速率均减慢,可能因为期间温度回升,空气湿度相较前期有所下降,从而减缓了尖叶匐灯藓的生长速率;而生长长度出现负增长的原因可能是尖叶匐灯藓为应对高温环境出现叶片卷缩而导致的测量误差(图3)。在整个栽培期间,WⅢ基质上尖叶匐灯藓生长长度在100 d达到最大值(2.81 cm),也是所有基质中尖叶匐灯藓所能达到的最大生长长度。
同时,为更确切判断不同栽培基质中尖叶匐灯藓的生长状况的差异,取120 d内各个时间段不同栽培基质中尖叶匐灯藓的生长长度平均值(图2,B)。其中,WⅢ基质中尖叶匐灯藓的平均生长长度最大,但与WⅡ和WⅣ基质处理无显著差异;而SG基质中尖叶匐灯藓的平均生长长度最小,其生长状况最差,并与其他基质差异明显。在整个栽培期间,尖叶匐灯藓的平均生长长度由大到小依次为WⅢ>WⅣ>WⅡ>SN>SC>SG。可见,倒木基质(WⅡ、WⅢ和WⅣ)栽培基质中的尖叶匐灯藓生长长度优于其他基质。
2.2.2 尖叶匐灯藓生长盖度尖叶匐灯藓生长盖度在不同栽培基质间也差异显著(图3;图4,A)。其中,栽培30 d时,尖叶匐灯藓的生长盖度在WⅣ基质中最大(2.59%),而在SG基质中最小(0.52%)。在整个栽培期间,SG基质中尖叶匐灯藓的生长盖度始终处于最小,WⅢ基质中尖叶匐灯藓生长盖度在培养100 d时达到最大(15.43%),且是所有基质中所达到的最大生长盖度。在培养100~120 d期间,所有基质中尖叶匐灯藓生长盖度在达到最大后,其生长速率减慢,甚至出现负增长,可能因为期间温度回升,空气湿度明显下降,影响了尖叶匐灯藓的生长蔓延。
同样,由不同栽培基质中尖叶匐灯藓的生长盖度平均值(图4,B)可以看出,WⅡ基质中尖叶匐灯藓的平均生长盖度最大,而与SN、WⅢ和WⅣ基质无显著差异;SG基质中尖叶匐灯藓的平均生长盖度最小,其生长状况最差,并明显低于其他基质;在120 d栽培期内,尖叶匐灯藓的平均生长盖度由大到小依次为WⅡ>WⅢ>WⅣ>SN>SC>SG。可见,倒木基质(WⅡ、WⅢ和WⅣ)中栽培的尖叶匐灯藓生长盖度优于其他基质。
2.3 栽培基质对尖叶匐灯藓部分生理指标的影响
尖叶匐灯藓的部分生理指标在不同栽培基质间存在显著性差异(表3)。叶绿素可以一定程度上反映植物的光合能力强弱,从而反映植株体内有机物的积累能力。其中,各基质中尖叶匐灯藓的叶绿素a、叶绿素b以及总叶绿素含量均以SC基质最高,且与其他基质间存在显著的差异,而又以SN基质最低,并与其余基质大多差异显著。同时,各基质中尖叶匐灯藓的叶绿素a/b值以SG基质最大,且与除SC和WⅡ以外的基质均有显著差异。说明SC基质中尖叶匐灯藓的生长代谢能力最强,有利于体内代谢产物的积累。可溶性糖可以提供植株生长所需的绝大部分能量,可溶性蛋白质含量在一定程度上代表植株体内酶的活性水平,较高的可溶性糖和可溶性蛋白含量有助于提高植物的抗逆性。表3显示,SG基质中尖叶匐灯藓的可溶性糖总含量显著高于其余基质,其可溶性蛋白质总含量也最高,它与SC基质无显著差异,与其他基质处理均存在显著差异。可见,SG和SC基质最有利于增加尖叶匐灯藓生长和抗性。
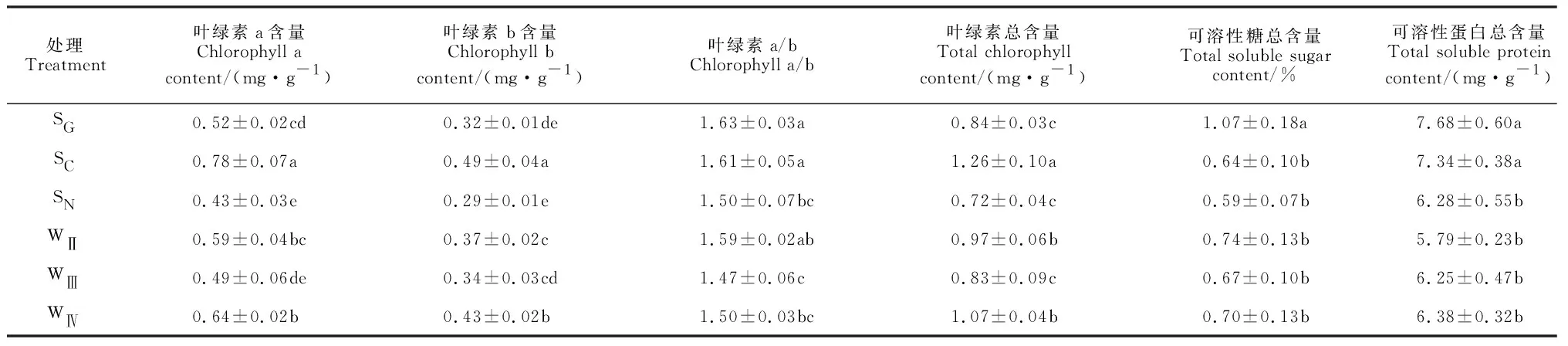
表3 不同栽培基质处理下尖叶匐灯藓部分生理指标的变化Table 3 Some physiological indexes of P. acutum under different substrate treatments
2.4 栽培基质理化性质与尖叶匐灯藓生长生理指标的相关性

表4 栽培基质理化性质与尖叶匐灯藓生长生理指标的Pearson相关系数Table 4 Pearson correlation coefficients between physical and chemical property indexes of culture substrates and growth physiological indexes of P. acutum
2.5 不同栽培基质下尖叶匐灯藓生长发育状况综合评价
判断植物生长状况的优劣应该是对植物的生长形态特征及内在营养成分的综合性状分析,单一的指标不能反映植物的本质属性,因此利用模糊隶属函数综合评价对尖叶匐灯藓的生长发育状况进行综合分析。表5显示,尖叶匐灯藓的生长形态指标与叶绿素a/b值、可溶性糖总含量和可溶性蛋白质总含量呈负相关关系,而与叶绿素a、叶绿素b、叶绿素总含量呈正相关关系。在利用隶属函数对尖叶匐灯藓的生长发育状况进行评价时,采用反隶属函数公式计算叶绿素a/b值、可溶性糖总含量和可溶性蛋白质含量的隶属函数值,用隶属函数公式计算叶绿素a、叶绿素b、叶绿素总含量的隶属函数值。结果(表6)表明,在所有栽培基质中,WⅣ基质的综合评价指数最高(0.755 2),而SG基质综合评价指数最低(0.079 7),并与其余基质差异明显;从尖叶匐灯藓生长发育状况综合评价指数来看,各栽培基质的表现由优到差依次为WⅣ>SC>WⅢ>WⅡ>SN>SG。这与生长指标的表现不尽相同。

表5 尖叶匐灯藓各项生长生理指标间的Pearson相关系数Table 5 Pearson correlation coefficients between growth physiological indexes of P. acutum
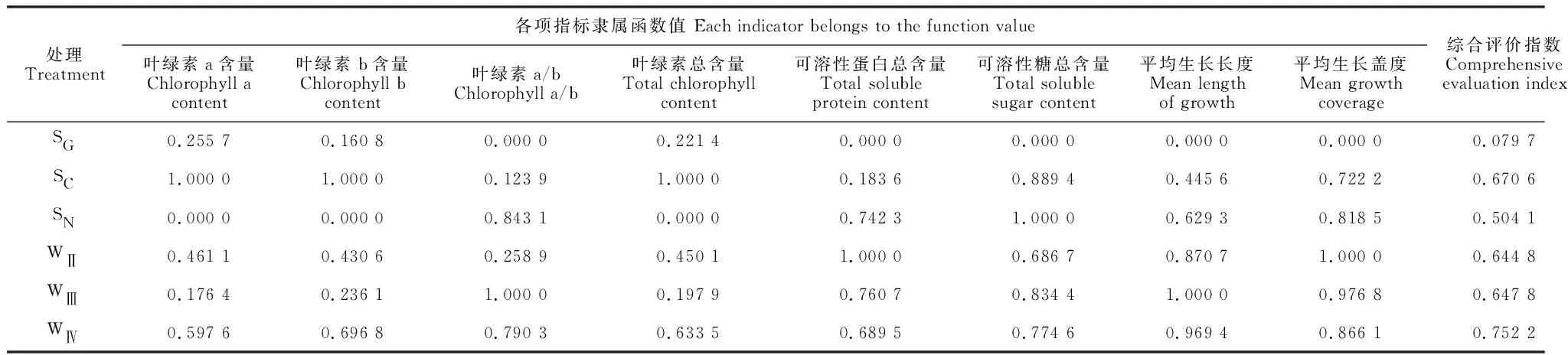
表6 不同栽培基质处理的尖叶匐灯藓生长发育状况综合评价Table 6 Comprehensive evaluation of growth and development of P. acutum under different cultivation substrates
3 讨 论
植株形态指标能反映其长势强弱,可用来判断不同栽培基质对植物生长的影响,从而判断该栽培基质的优劣[19],而基质的材料、结构、性质是影响植株生长状况的关键因素。同时,植物能够茁壮生长也离不开体内各种物质参与的代谢和调节。叶片光合色素含量可以反映作物光合作用能力的强弱,与作物的生长潜能有关[20],适当强度的光合作用将有利于植株体内有机物的积累[21]。可溶性糖和可溶性蛋白质可维持植物体细胞的渗透平衡,提高植物生物抗逆性[22]。本研究发现SC(草炭土)基质中尖叶匐灯藓的平均生长长度和平均生长盖度指标并不突出,但是SC处理尖叶匐灯藓生长状况的综合评价指数排名第二,与生长表现较好的倒木基质评价指数接近,主要原因在于SC基质中尖叶匐灯藓的叶绿素含量显著高于其他基质;而通过基质的理化性质对比,SC基质除全氮含量在所有基质中含量最高外,其他理化性质并不突出,试验证明全氮含量与尖叶匐灯藓的叶绿素含量呈极显著正相关;且有研究证明植物中叶绿素含量与氮含量有很强的相关性[23-24],因此可能因为SC基质中较高的氮含量而提高了尖叶匐灯藓中的叶绿素含量,从而综合评价排名靠前。通过Pearson相关性分析,发现尖叶匐灯藓的可溶性蛋白质含量和可溶性糖含量与生长长度和盖度呈负相关关系,在所有基质中SG(颗粒土)上尖叶匐灯藓的可溶性糖含量和可溶性蛋白质含量最高,叶绿素总含量较低,其生长状况最差。这个原因可能是相比其他基质SG基质的全氮含量较少,总孔隙度小,持水能力差,从而影响其正常光合和生长,而尖叶匐灯藓为适应环境的影响,通过提高可溶性糖含量和可溶性蛋白质含量来增加自身生物抗性,维持正常生命活动。冯伟文研究发现泥炭土和菜园土最适宜尖叶匍灯藓的生长,木碎片上尖叶匍灯藓生长效果最差,覆盖面积最小[1],而本在研究中倒木基质栽培效果较好,可能是本研究中倒木与木碎片的种类、颗粒大小存在差异而导致结果不同,且本研究仅基于栽培基质的理化性质和苔藓生长状况推断倒木适用于苔藓栽培,该结论是否具有普遍性还需要有更多的研究来验证。
苔藓植物群落的生长发育及其对养分的吸收与利用受生长基质质量和光照等环境因子的影响[25-26]。郝占庆发现不同苔藓盖度下土壤中氮、钾含量差别不大,但没有苔藓覆盖的土壤中磷含量最高,并与其他土壤差异显著,且随苔藓盖度的增加土壤中磷含量有减少的趋势[27];而本研究发现尖叶匐灯藓的生长形态指标与基质中的全氮含量呈极显著正相关,而与全磷和全钾含量呈显著负相关,这与郝占庆的研究结果不一致,可能因为苔藓种类的不同而存在差异,且本实验并没有对尖叶匐灯藓内部的氮、磷、钾含量进行测定,关于苔藓对基质中的养分吸收有待进一步研究。
另外,不同栽培基质对尖叶匐灯藓生长和生理代谢的影响效应具有差异性,而尖叶匐灯藓生长还受到外界环境等多种因素的影响[8]。土壤或基质中细菌的各个生理类群参与了种植环境中的物质循环,其种类和数量直接反映种植生态环境系统结构的稳定性和差异性[28]。有研究报道指出苔藓与微生物之间存在互利共生的作用[29],倒木中含有丰富的微生物[30-31],而在本研究中倒木基质更适宜尖叶匐灯藓的生长,是否是因为倒木中的某种微生物促进了尖叶匐灯藓的生长尚有待进一步考证。
综上所述,本研究中6种栽培基质的理化性质存在显著差异,倒木基质(WⅡ、WⅢ和WⅣ)的总孔隙度、持水能力以及全氮含量显著高于颗粒土(SG)和通用营养土(SN)基质,而SG和SN基质的容重、全磷含量和全钾含量显著高于倒木基质;草炭土基质(SC)除总孔隙度和持水孔隙与倒木基质有显著差异外,其他指标与SG和SN基质较为接近。通过模糊隶属函数评价可知,WⅣ基质(Ⅳ级马尾松倒木)综合评价指数最高,其上尖叶匐灯藓综合生长发育状况最好,可以作为栽培尖叶匐灯藓最优基质。本研究证明适当增大基质中的孔隙度、持水能力和全氮含量将有利于尖叶匐灯藓的生长发育。
