火龙果采后病原菌的分离鉴定及丁香酚抑菌研究*
2022-09-01罗冬兰雷霁卿瞿光凡陈建业范中奇巴良杰
罗冬兰,雷霁卿,曹 森,瞿光凡,朱 婷,陈建业,范中奇,巴良杰
(1 贵阳学院,贵州550005)(2 华南农业大学)(3 福建农林大学)
火龙果原产于墨西哥、中美洲等热带地区,以肉色不同将其分为红肉、白肉、黄肉火龙果[1]。其中,红肉火龙果因其富含较高的营养物质、独特的风味深受消费者喜爱[2]。然而,火龙果成熟期为高温多雨季节,采后田间热、较高的呼吸强度极易受病原菌的侵染,从而引起果实快速腐烂变质,货架期短[3-4]。据报道,火龙果采后主要病害有炭疽病、黑斑病、果腐病、溃疡病、软腐病[5]。病原菌的侵染是导致采后火龙果品质劣变的主要因素。因此,控制果实采后病害发生对提高果实商品价值和产业发展尤其重要。
丁香酚作为一种天然的植物精油,其挥发性成分主要有菇烯、菇类化合物和酚类物质,具有抑菌活性和诱导增强果蔬抗氧化等作用[6]。周丹丹等[7]研究发现,1 μL/L 丁香酚处理能有效抑制枇杷采后炭疽病的发生,在贮藏6 d 时,其对尖孢炭疽菌的抑制率为66.7%。葛达娥等[8]研究结果表明,丁香酚通过破坏链格孢菌的细胞膜通透性,抑制菌丝生长和孢子萌发,从而有效抑制蓝莓采后黑斑病的发生。此外,米嘉琦等[9]研究发现,丁香酚处理不仅抑制芒果采后炭疽病的发生,还有效降低了芒果的腐烂率。然而,关于丁香酚对火龙果采后病原菌的抑制研究未见报道。本研究对自然发病的火龙果进行病原菌的分离鉴定,并筛选适宜浓度的丁香酚对病原菌进行抑制研究,以期为火龙果采后病害的防控提供理论基础。
1 材料与方法
1.1 材料
火龙果鲜果于2021 年9 月11 日从贵州省罗甸县火龙果种植基地采摘,选取健康、无病虫害果实用PE20 袋进行单个包装,并置于温度20 ℃、空气相对湿度85%条件下贮藏,定期观察果实发病情况。病原菌的分离培养基为PDA 培养基(马铃薯200 g,琼脂20 g,葡萄糖20 g,蒸馏水1 000 mL)。

火龙果采后病原菌的形态特征及致病性测定结果Morphological characteristics of postharvest pathogens of pitaya fruit and its pathogenicity test results
1.2 火龙果采后优势菌分离
采用组织分离法对病原菌进行分离[10]。将自然发病的果实置于75%酒精消毒30 s,无菌水冲洗3次,再用无菌吸水纸将果皮水分擦干,采用无菌手术刀切取果实病健交界处组织,放于PDA 培养基上,经过多次划线分离、单孢培养直至获得纯化菌株,并将纯化菌株于4 ℃保存备用。
1.3 病原菌传统形态学观察
将分离的病原菌接种在新的PDA 平板上培养15 d,观察记录分离菌株的菌落生长情况,利用光学显微镜观察记录菌丝和孢子的形态结构特征,并参照《真菌鉴定手册》[11]对病原菌进行初步鉴定。
1.4 病原菌致病性检测
将火龙果果实用无菌水清洗后,在75%酒精中浸泡30 s,再用无菌水清洗3 次,用无菌打孔器在果实赤道处刺直径4 mm、深3 mm 的伤口,在伤口处分别接种培养5 d 病原菌菌饼,以无菌菌饼作为对照,每株病原菌3 个果。置于温度25 ℃、空气相对湿度80%~90%条件下恒温培养4 d 后观察记录果实发病情况,根据柯赫氏法则对病原菌进行致病性检测[12-13]。
1.5 病原菌rDNA-ITS 序列分析
1.5.1 病原菌DNA 提取
取一定量纯化后培养5 d 的病原菌菌丝,参照真菌基因组DNA 快速抽提试剂盒(生工生物工程)提取病原菌DNA。
1.5.2 病原菌ITS 区PCR 扩增及凝胶电泳
选用真菌通用引物ITS1(5′-TCCGTAGGTGAA CCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGAT ATGC-3′)进行PCR 扩增。PCR 反应体系:TaqMast erMix 110 μL、ITS1 和ITS4 1 μL、模板DNA 1 μL、ddH2O2补加至20 μL。扩增程序:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,30 个循环;72 ℃终延伸10 min[14]。PCR 产物经1%琼脂糖凝胶电泳检测后,寄送生工生物工程(上海)股份有限公司进行测序,测序结果在NCBI 网站的GenBank 数据库中进行BLAST 同源性比对分析。
1.6 丁香酚对火龙果病原菌体外抑菌效果
采用菌丝生长速率法[15]测定丁香酚对火龙果采后病原菌菌丝生长的影响。称取一定质量丁香酚液配制成10 mg/mL 母液,分别吸取一定量的母液加入到10 mL PDA 培养基中,使得丁香酚质量浓度为0.05、0.10、0.15、0.20、0.25 mg/mL,充分摇匀后倒入无菌培养皿中,备用。用无菌打孔器从纯化后培养5~7 d 的病原菌菌落边缘打取直径4 mm 菌饼放于PDA 培养基中央,以不加丁香酚培养基作为对照(CK),每个处理3 次重复。28 ℃培养7 d 或对照组长满整个培养基后测量菌落直径,并参照王丹等[16]方法计算抑菌率。
1.7 数据处理
采用Excel 2016 软件进行数据统计,采用SPSS Statistics 23 软件的Duncan’s 法对数据进行差异显著性分析,采用Origin 2018 软件作图。
2 结果与分析
2.1 火龙果自然发病症状
贮藏3 d 后,果实鳞片开始萎蔫;贮藏5 d 后,局部鳞片和果柄处长出少量菌丝;贮藏8 d 后,果柄处长出大量白色、灰色和灰白色菌丝,肉眼可见,并覆盖整个果柄面,菌丝逐渐向果身扩展;同时,果柄周围果皮褐腐溃烂,果皮出现许多小黑点(图版1-a),失去食用价值。
2.2 病原菌的形态特征
将自然发病的火龙果果实采用组织块分离法分离得到4 株病原菌,记作H-1、H-2、H-3 和H-4。菌株H-1 在PDA 培养基上培养(图版1-b-A1~A3),菌丝生长较快,初期为灰白色棉絮状,后期变为灰黑色或暗灰色,菌落基质初期为暗黄色,后期变为棕褐色;在显微镜下观察发现,菌落培养7 d后产生成熟的分生孢子,孢子呈椭圆形、卵圆形或短棒形,孢子两端钝圆,无色,无膈膜,串珠状排列。根据其形态特征并参照真菌分类手册,初步鉴定为新暗色柱节孢菌(Neoscytalidium dimidiatum)。
菌株H-2在PDA培养基上培养(图版1-b-B1~B3),菌丝初期为白色,后逐渐变为淡棕色,后期呈红棕色,并分泌红褐色色素;菌落基质呈现明显同心轮纹,外层暗黄色,内层红棕色;菌落培养7~15 d 产生分生孢子,分生孢子呈球形、近球形,无膈膜。根据其形态特征并参照真菌分类手册,初步鉴定为黑附球菌(Epicoccum nigrum)。
菌株H-3 在PDA 培养基上培养(图版1-b-C1~C3),初期为白色绒毛状,后期变为浅黄色;菌落基质初期为米黄色,后期呈黄棕色;培养5 d 后产生分生孢子,分生孢子弯曲显著或微弯,呈镰刀形,有膈膜,横膈膜1~3 个,无色。根据其形态特征并参照真菌分类手册,初步鉴定为镰刀菌属(Fusariumsp.)。
菌株H-4 在PDA 培养基上培养(图版1-b-D1~D3),菌落初期为白色绒毛状,菌丝呈网状形展开生长,后期变为暗黄色;菌落基质初期为米黄色,后期呈暗黄棕色或暗灰色;在显微镜下观察发现,菌丝粗壮,内有油球,菌落培养7 d 后,产生分生孢子,孢子呈球形、近球形,无膈膜。根据其形态特征并参照真菌分类手册,初步鉴定为球黑孢菌(Nigrospora sphaerica)。
2.3 病原菌的致病性测定结果
选取无病虫害、无机械损伤的果实,将果实置于75%酒精中浸泡30 s,无菌水冲洗2~3 次后放于超净工作台中自然晾干,备用。采用刺伤接种法将菌株H-1、H-2、H-3 和H-4 分别接种至火龙果果实上,对照组接无菌菌饼,并将果实置于温度25 ℃、空气相对湿度85%条件下贮藏4 d。刺伤接种菌株H-1(图版1-c-A)果实,刺伤部位长出灰白色菌丝,果实开始褐腐溃烂。刺伤接种菌株H-2(图版1-c-B)、H-3(图版1-c-C)、H-4(图版1-c-D)引起火龙果致病,其刺伤部位长出大量菌丝,肉眼可见,对照组均无发病。取接种发病果实组织再次进行分离,分离得到的菌株与原菌株培养性状一致。
2.4 病原菌的分子生物学鉴定
将火龙果病原菌PCR 产物经1%琼脂糖凝胶电泳检测得到4 株病原菌序列长度均为500~700 bp;并将其产物送至生工生物工程(上海)股份有限公司进行测序,测序结果提交NCBI 网站的GenBank数据库中进行BLAST 同源性比对分析,并下载同源较高的序列,采用邻近法构建系统发育树(图1)。由系统进化树分析结果得知,菌株H-1 与Neoscytalidium dimidiatum具有较高的同源性,其相似度高达100%,结合其形态特征鉴定为新暗色柱节孢菌;菌株H-2 与Epicoccum nigrum处于同一分支,相似度达91%,结合其形态特征鉴定为黑附球菌;菌株H-3 与Fusarium oxysporum有较高同源性,相似度达99%,结合其形态特征鉴定为尖孢镰刀菌;菌株H-4 与Nigrospora sphaerica有较高的同源性,相似度达100%,结合其形态特征鉴定为球黑孢菌。

图1 火龙果4 株病原菌的rDNA-ITS 序列PCR 电泳图及系统发育树
2.5 丁香酚对火龙果病原菌菌丝抑制及室内毒力测定
2.5.1 丁香酚对病原菌菌丝抑制效果
不同质量浓度丁香酚对火龙果采后病原菌菌丝生长抑制效果见表1。由表1 可知,随着丁香酚质量浓度的增加,其对4 株病原菌的抑制率呈递增趋势,且不同浓度间抑制效果差异显著。当丁香酚浓度为0.20 mg/mL 时,对4 株病原菌的抑制率大于70%,其中,完全抑制F.oxysporum生长。当丁香酚浓度为0.25 mg/mL 时,对N.dimidiatum、E.nigrum、F.oxysporum和N.sphaerica的抑菌率分别为87.66%、97.49%、100.00%和95.07%。由此可见,丁香酚对火龙果采后病害的防治具有潜在的应用价值。
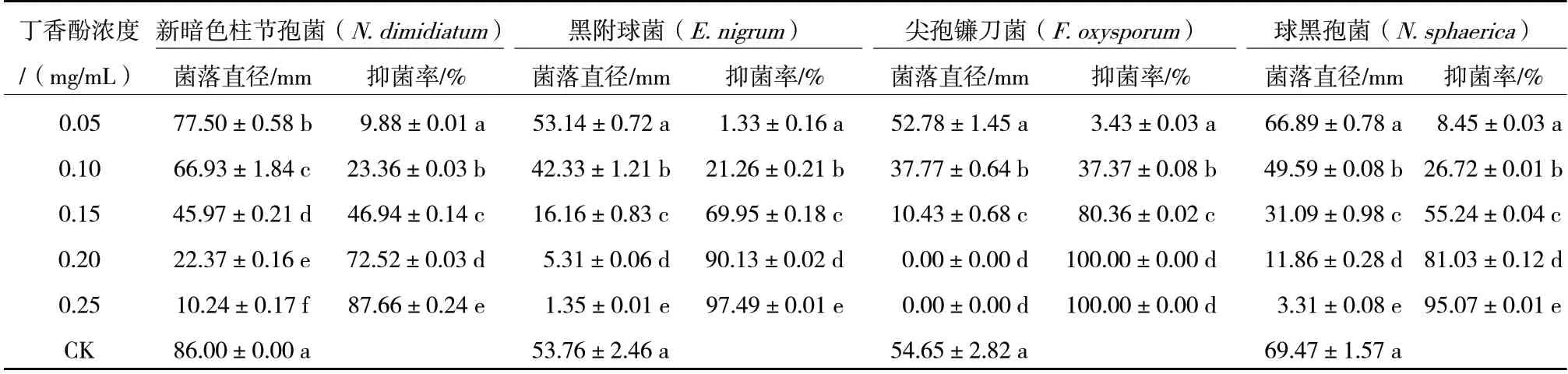
表1 丁香酚对火龙果采后病原菌菌丝生长的抑制效果
2.5.2 丁香酚对病原菌菌丝室内毒力测试
通过菌丝生长抑制试验,求出丁香酚对4 株病原菌菌丝毒力回归方程及EC50,结果见表2。由表2可知,F.oxysporum对丁香酚的敏感性最高,EC50为0.114 7 mg/mL,其他依次为E.nigrum、N.sphaerica、N.dimidiatum,EC50分别为0.128 3、0.156 0、0.174 8 mg/L。

表2 丁香酚对火龙果采后病原菌菌丝生长的毒力回归方程
3 讨论与结论
近年来,火龙果采后病害的发生造成严重的经济损失。据前人研究报道,崔志婧等[17]研究发现,上海市进口火龙果软腐病主要由尖孢镰刀菌(Fusarium oxysporum)和单隔镰刀菌(Fusarium dimerum)引起;郭力维等[18]认为,引起云南火龙果果腐病的病原菌为霜霉目白秀科桃吉尔霉(Gilbertella persicaria);Guo 等[19]首次发现平头炭疽菌(Colletotrichum truncatum)可引起火龙果炭疽病发生;王会会等[20]发现引起火龙果溃疡病的致病菌为新暗色柱节孢菌。此外,胡翠平等[21]还发现,引起广西白心火龙果采后腐烂的致病菌为新月弯孢菌(Curvularia lunata)。综上研究报道可知,引起火龙果腐烂的病原菌种类较多,且致病力可能与火龙果品种、病原菌种类及区域条件有关。本研究从自然发病的火龙果果实分离病原菌,并经过致病性测定、形态学及分子生物学鉴定,确定引起贵州省罗甸县火龙果采后腐烂的病原菌主要是新暗色柱节孢菌(Neoscytalidium dimidiatum)、黑附球菌(Epicoccum nigrum)、尖孢镰刀菌(Fusarium oxysporum)和球黑孢菌(Nigrospora sphaerica)。
丁香酚不仅具有广谱杀菌性,而且在果蔬采后保鲜上具有潜在的应用价值;能够有效提高枇杷[7]、草莓[22]的贮藏品质,降低青茄[23]冷害发生,抑制苹果[6]采后病原菌生长。本研究发现,丁香酚对火龙果采后病原菌具有较好的控制效果。其中,当丁香酚浓度为0.20 mg/mL 时,其对4 株病原菌的抑制率均大于70%。通过毒力测试发现,丁香酚对F.oxysporum抑制效果最显著,EC50为0.114 7 mg/mL,其次为E.nigrum、N.sphaerica、N.dimidiatum,EC50分别为0.128 3、0.156 0、0.174 8 mg/L。因此,丁香酚可应用于火龙果采后病害防控。然而,关于丁香酚对火龙果采后病原菌的抑菌机理及贮藏保鲜效果还需进一步的研究。
