山地草甸植被生物多样性及群落稳定性对坡向的响应
2022-08-31乔欢欢何国兴杨冬阳关文昊刘志刚杨军银柳小妮
乔欢欢, 李 强, 何国兴, 纪 童, 杨冬阳, 文 铜, 关文昊, 刘志刚, 杨军银, 柳小妮
(甘肃农业大学草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
草地植被特征是草地生产稳定性、生态稳定性调控研究的基础。群落稳定性是一个相对的特征,它与研究区域的气候特征、地理位置、植物群落结构、土壤养分特征等因子密切相关[1],且植物群落结构与功能共同影响群落稳定性[2]。深入了解草地植被的群落组成、结构和功能,才能使草地的生产、生态功能得以全面发挥[3]。明确植物群落生物多样性与群落稳定性的变化规律是研究草地群落演替阶段的重要依据,对草地生态保护和合理利用具有重要的科学意义。
环境筛选、物种间的相互作用和种群构成的随机性决定了自然群落的生物多样性和群落稳定性[4-6]。坡向决定着植被受到的光热条件,通过改变坡向对太阳辐射的接收量和水文过程,会影响植被的发育[7],进而对生物多样性和群落稳定性产生重要影响[8-9]。国内学者相关研究发现,坡向的变化对植物的生长和分布具有显著影响[10-11]。杨倩等[12]发现不同坡向下土壤因子对植被分布的贡献不同。张英团等[13]发现坡向对山地环境的丰富度和优势度影响明显。刘方炎等[14]发现位于干热河谷北坡的灌草丛和西北坡的群落稳定性明显优于其他坡向的植物群落。
祁连山是我国西部青藏高原和北部内陆高原荒漠地区草原自然生态的重要分水岭,具有优良的草地成土生态环境和独特的草原生物环境多样性[15]。分布在祁连山的山地草甸是以中生植物为主组成的群落,其发育条件为山地温带气候,海拔高,气温低,蒸发蒸腾量小,对维持植被群落结构和保障生态系统功能具有重要作用[16]。学者在祁连山的其它地区展开许多草地多样性和生物量方面的研究[17-19],而东祁连山不同生境,特别是不同坡向的山地草甸植被生物多样性和群落稳定性的报道相对较少。祁连山生态环境脆弱,一旦植被遭到破坏,其自然恢复不仅是长期的,而且是极其困难的[20],并且祁连山草地围栏区块化管理未考虑微地形对草地生态系统服务能力的影响[21]。因此本研究调查了祁连山东段金强河流域不同坡向山地草甸的植被特征,分析山地草甸的植被群落成特征、生物多样性和群落稳定性的差异性,可以更好的认识群落组成、变化和发展,为祁连山草地的合理利用和恢复提供一定借鉴依据。
1 材料与方法
1.1 研究区概况
试验地位于祁连山东部的甘肃天祝藏族自治县抓喜秀龙乡金强河流域(37°40′N,102°32′E)处的山地草甸,平均海拔2 960 m。该区全年昼夜温差大,年均温在-0.1℃~0.6℃间,≥0℃年积温1 380℃·d-1,年均降水量416 mm,多集中于7—9月,年蒸发量为1 592 mm。全年无绝对无霜期,植株的生长周期120~140 d。土壤类型主要为山地暗栗钙土,分布的优势植物有线叶嵩草(Kobresiacapillifolia)、珠芽蓼(Polygonumviviparum)、多裂萎陵菜(Potentillamulatifida)、垂穗披碱草(Elymusnutans)等。
1.2 样地设置和调查
2020年8—9月,提前对山地草甸地形与植被的进行考察,选取了坡位基本相同和全年禁牧的山地草甸为研究对象,消除坡位、放牧强度等对研究结果的干扰。在海拔2 800 m处选择了位于阴坡(坡度20°)、半阴坡(坡度23°)、半阳坡(坡度23°)、阳坡(坡度23°)的4个样地,选取的样地大小均为30 m×30 m,各样地每隔20 m设置一个样点并钉上木桩作为标记,然后在每个样点的上、中、下分别设置一个1 m×1 m的样方,样方之间的距离为10 m,调查植物群落的物种组成及其盖度、高度(自然高度下各植物种分别测 10 株的平均值)。分种采集样方内地上生物量(齐地面刈割,放入样品袋中),称鲜重后,带回实验室于105℃烘箱中杀青2 h后,60℃下恒重称重。采集完样方内的地上生物量后,去除土层上的枯落物后,采用环刀法[22](100 cm3)取样,测定土壤含水量(Soil water content,SWC)。根据祁连山山地草甸群落中不同植物物种的生活型(包括形态、繁殖和扩散特征等)的差异[23],我们将所有物种划分为以下四个功能群:禾本功能群、豆科功能群、莎草功能群和杂草功能群。坡向的量化方法参照王孝安[24]。

图1 样地空间分布示意图Fig.1 Schematic diagram of spatial distribution of sampling sets注:S,阳坡;W,半阳坡;E,半阴坡;N,阴坡Note:S,Sunny slope;W,Semi-sunny slope;E,Semi-shady slope;N,Shady slope
1.3 指标及其计算
1.3.1植被含水率和土壤含水量 植被含水率的计算公式如下[25]:

(1)
土壤含水量测定方法[22]为:
土壤含水量的测定:将采用环刀法采集的原状土带回实验室,称重(鲜土+环刀重)为M1,放置在烘箱,105℃烘至恒重,称重(干土+环刀)为M2,将干土去掉环刀称重为M3。土壤含水量计算公式如下:

(2)
1.3.2草地生物多样性 草地生物多样性指数计算公式如下[26]:
Margalef物种丰富度:d=(S-1)/lnN
(3)
Pielou物种均匀度:E=H′/lnS
(4)
(5)
Shannon-Weiner多样性:H=PilnPi
(6)
式中,为样方内的植物种i的相对重要值,重要值=(相对盖度+相对高度+相对地上生物量)/3,S为样方内的物种数量,N为样方内的全部植株数量。
1.3.3植物群落Godron稳定性 采用郑元润[27]和罗久富等[28]对Godron稳定性[29]测定方法改进后的数学方法,用一元三次方程进行拟合,提高拟合精确度[30]。作山地草甸4个坡向植物群落Godron稳定性拟合曲线。先将样方中不同种植物的盖度按降序排列,并转换为相对盖度,按相对盖度降序逐步累加起来,然后将样方内植物种类的总和取倒数,按植物种类的顺序逐步累积,再将植物种类倒数百分率同累积相对盖度一一对应,绘制散点图并用平滑曲线连接。然后再与直线y=100-x相交,交点即为植物群落稳定性参考点。一般地,稳定性比值越接近 20/80,群落就越稳定。
1.4 数据处理
采用Excel 2021和SPSS 21.0软件进行数据整理分析。利用单因素分析(Oneway ANOVA) 和Duncan极差法进行差异显著性检验。应用Origin2022绘制植被多样性箱线图和相关性分析图,其中相关性分析用皮尔逊相关分析法(Pearson’s correlation)确定显著性。图表中数据为均值±标准误。
2 结果与分析
2.1 植被特征
由表1可知,4个样地植被盖度、草层高度和地上生物量均呈现出阴坡>半阴坡>半阳坡>阳坡的分布格局(表1),且不同坡向的植被盖度差异显著(P<0.05)。阴坡的草层高度显著高于阳坡(P<0.05),半阴坡、半阳坡和阳坡的草层高度差异不明显。在阳坡的地上生物量显著低于阴坡、半阴坡和半阳坡(P<0.05),阴坡、半阴坡和半阳坡差异不显著。

表1 不同坡向山地草甸植被特征Table 1 Vegetation characteristics of mountain meadow at different aspect
2.2 不同植物功能群的地上生物量
莎草科地上生物量在4个样地中呈现出阴坡>半阴坡>半阳坡>阳坡的变化趋势(表2)。阴坡的莎草科地上生物量显著高于半阴坡、半阳坡和阳坡(P<0.05),阳坡的莎草科地上生物量和豆科地上生物量显著低于阴坡、半阴坡和半阳坡(P<0.05),禾本科地上生物量呈现出半阴坡>半阳坡>阴坡>阳坡的变化趋势,阴坡和阳坡禾本科地上生物量显著低于半阴坡和半阳坡(P<0.05)。豆科地上生物量呈现出半阳坡>阴坡>半阴坡>阳坡的变化趋势。杂类草地上生物量呈现出阳坡>半阳坡>半阴坡>阴坡的变化趋势,阴坡和半阴坡杂类草地上生物量显著低于半阳坡和阳坡(P<0.05)。

表2 不同坡向山地草甸植物功能群生物量Table 2 Plant group functional of mountain meadow at different aspect
从植物功能群地上生物量的组成比例来看(图2),随着阴坡向阳坡的变化,莎草科地上生物量比例依次降低。禾本科地上生物量比例先升高后降低。而豆科地上生物量比例呈波动性下降。杂类草地上生物量比例依次升高。

图2 不同坡向山地草甸植物功能群组成比例Fig.2 Plant group functional composition ratio of mountain meadow at different aspect
2.3 土壤含水量和地上植物含水率
在阴坡-阳坡梯度上,山地草甸土壤含水量和地上植物含水率变化规律相同,依次降低(图3),阴坡的土壤含水量和植物含水率显著高于阳坡(P<0.05),其他坡向之间差异不显著。
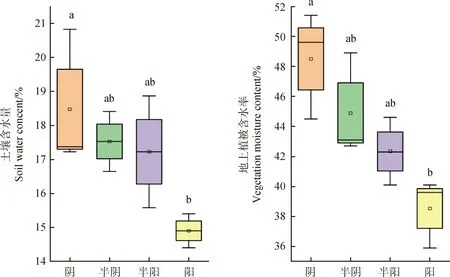
图3 不同坡向山地草甸土壤含水量和地上植物含水率Fig.3 Soil water content and plant water content of mountain meadow at different aspect
2.4 植被生物多样性变化
从阴坡到阳坡,山地草甸植物Shannon-Wiener多样性指数先升高后降低(图4A),四个坡向之间无显著差异。Margalef丰富度指数变化和Shannon-Wiener多样性指数基本类似(图4D)。山地草甸植物Pielou均匀度指数呈现出先降低后上升的变化趋势(图4B),半阴坡Pielou均匀度指数显著低于阴坡、半阳坡和阳坡(P<0.05),阴坡、半阳坡和阳坡之间无显著差异。山地草甸植物Simpson优势度指数呈现出半阴坡>阴坡>阳坡>半阳坡的变化趋势(图4C),呈波动性上升,四个坡向之间无显著差异。

图4 不同坡向下山地草甸植被生物多样性Fig.4 Vegetation biodiversity of mountain meadow at different aspect
2.5 植物群落稳定性
阴坡的稳定性拟合曲线与直线交点坐标为(32.16,67.84) (表3,图5),与植物群落稳定参考点(20,80)的欧式距离为17.20。半阴坡和半阳坡的稳定性拟合曲线与直线的交点坐标分别为(33.36,66.64)和(32.17,67.83),与植物群落稳定参考点(20,80)的欧式距离分别为18.89和17.22(表3,图5),半阴坡和半阳坡的稳定性拟合曲线与直线的欧氏距离均大于阴坡。阳坡的稳定性拟合曲线与直线的交点坐标为(52.09,47.91),交点坐标与植物群落稳定参考点(20,80)间的欧式距离为45.38(表3,图5),大于阴坡、半阳坡和半阴坡的欧式距离。

表3 坡向对山地草甸植物群落稳定性的影响(Godron法)Table 3 Effects of slope aspect on plant community stability in mountain meadow method (Godron’s)

图5 坡向对山地草甸植物群落Godron稳定性拟合曲线Fig.5 Godron stability simulated curves of plant community under slope aspect in mountain meadow
2.6 相关性分析
相关性分析发现(6),TC与GH,Cyp,PWC和SWC呈极显著正相关(P<0.01),与AGB呈显著正相关(P<0.05),与Forbs和S呈极显著负相关(P<0.01)。GH与Cyp和SWC呈极显著正相关(P<0.01),与AGB呈显著正相关(P<0.05),与Forbs呈显著负相关(P<0.05)。AGB与Cyp呈极显著正相关(P<0.01),与Forbs和S呈显著负相关(P<0.05)。Pielou指数与Simpson指数呈极显著负相关(P<0.01),与Margalef指数呈显著负相关(P<0.05)。Margalef指数与Simpson指数呈极显著正相关(P<0.01),与Forbs呈极显著负相关(P<0.01)。Cyp与PWC呈极显著正相关(P<0.01),与Leg和SWC呈显著正相关(P<0.05),与Forbs和S呈极显著负相关(P<0.01)。Gra与S呈显著负相关(P<0.05)。Leg与SWC呈显著正相关(P<0.05),与S呈极显著负相关(P<0.01)。Forbs与PWC呈显著负相关(P<0.01)。PWC与SWC呈显著正相关(P<0.05),与S呈显著负相关(P<0.05)。SWC与S呈显著负相关(P<0.05)。
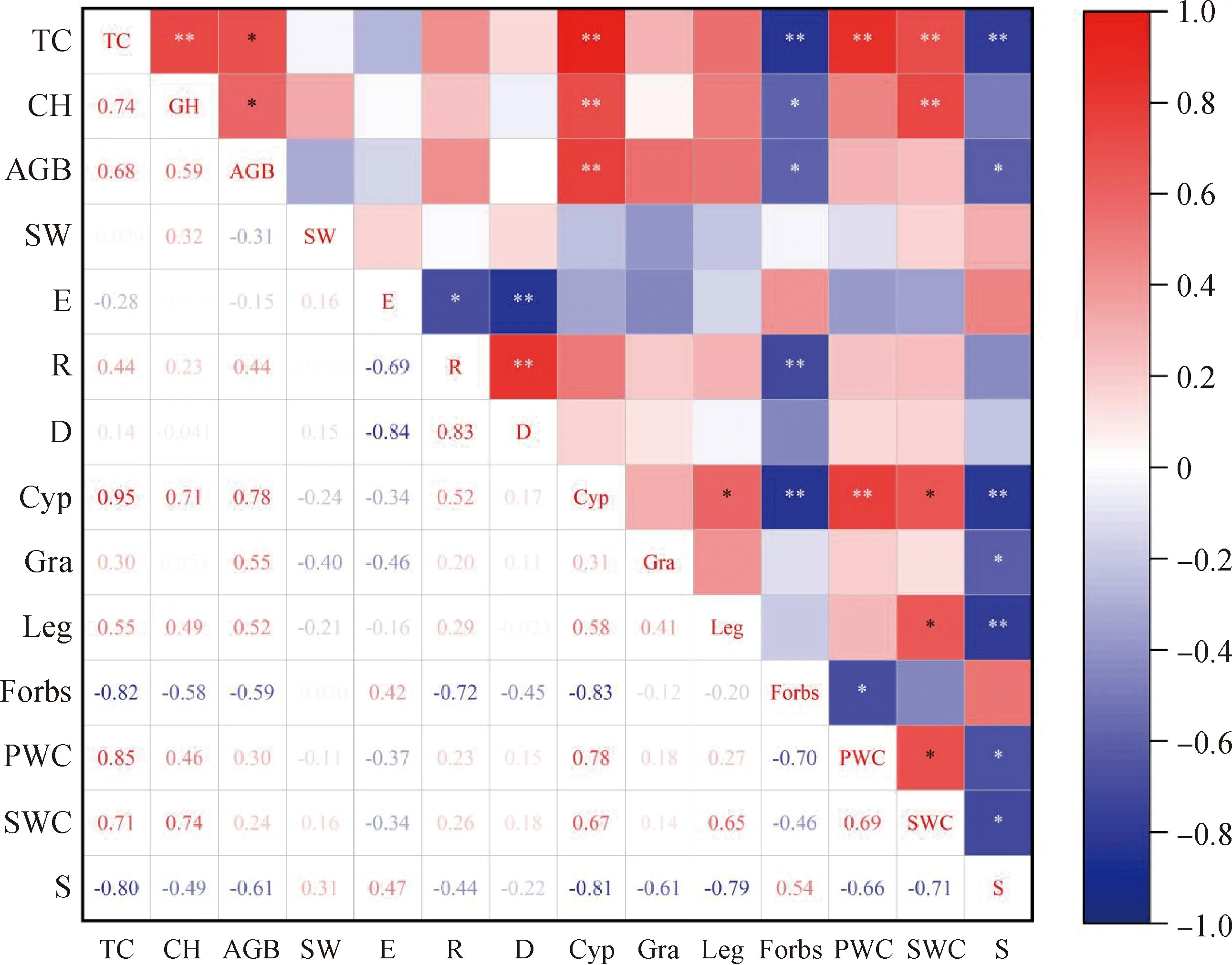
图6 不同坡向山地草甸植被指标的相关性分析Fig.6 Correlation analysis of vegetation indicators in different aspect注:*和**分别表示在P<0.05 and P<0.01。TC,总盖度;GH,高度;AGB,地上生物量;PWC,植被含水率;SW,Shannon-Weiner多样性指数;E,Pielou物种均匀度;R,Margalef物种丰富度;D,Simpson物种优势度;Gra,禾本科;Cyp,莎草科;Leg,豆科;Forbs,杂类草;SWC,土壤含水量;S,群落稳定性Note:* and ** indicate significance at the 0.05 and 0.01 level. TC,Total Coverage;GH,Grass layer height;AGB,Above-ground biomass;PWC,Plant water content;SW,Shannon-Weiner index;E,Pielou Evenness;R,Margalef Richness;D,Simpson Dominance;Gra,Graminea;Cyp,Cyperace;Leg,Legumios;Forbs,Forbs;SWC,Soil water concent;S,Community stability
3 讨论
坡向是影响山地草甸植物分布和生长的重要因子[31]。本研究中,在阴坡-阳坡梯度上,土壤含水量和物种的盖度、高度、地上生物量、植被含水率从阴坡到阳坡呈递减的趋势,这说明阴坡有充足的水分及养分资源供给植物,植物生长良好,植株个体较大,生物量高。阳坡因为温度和光照度较高,蒸发量大,致使土壤含水量较低,降低了物种生长速率,植株矮小,这与Baher[32]的研究结果一致。由此可见,在从阴坡到阳坡的坡向梯度上,各植被特征由于坡向的不同而存在差异,且存在明显的梯度变化。
由于长期的自然选择,植物群落对各自的生态位均具有适应性。不同坡向对水热异质性的分派,导致草地植被功能群随着坡向也发生着变化[33]。本研究发现,阳坡的主要功能群是杂类草,而阴坡的主要功能群为莎草科,这与刘玉祯等[29]发现阴坡以禾本科植物为主不一致。可能是阴坡生长季前期均以各自群落中对水热条件要求相对较低或生长季相对较短的禾本科和部分菊科物种为主,取样时由于部分物种的生长季已完成,导致阴坡草地群落则以莎草科等为主[34]。在阴坡-阳坡梯度上,阳坡的莎草科、禾本科和豆科的地上生物量均低于阴坡、半阴坡和半阳坡,杂类草比例阳坡高于阴坡、半阴坡和半阳坡,这与张倩[12]的研究结论类似。
草地植被的物种多样性能够很好地体现群落的结构类型、组织水平、演替阶段、稳定程度和生境差异性[35-36]。在本研究中,在阴坡-阳坡梯度上,Shannon-Wiener指数和Margalef指数变化一致,表现为半阴坡>半阳坡>阴坡>阳坡。坡向对Margalef指数、Simpson指数、Shannon-Wiener指数的影响较小,而对Pielou指数具有一定的影响,半阴坡显著高于阴坡、半阳坡和阳坡(P<0.05),这与曹运强等[37]对油松中龄林林下植物多样性的研究结果不同,这主要是植被类型不同所致。各测定指标部分组内差异较大,可能由于取样点之间差异较大引起。
植物群落稳定性可反映植物的种间竞争及群落抵抗环境压力和人为扰动的能力[38]。在阴坡-阳坡梯度上,应考虑植物群落结构和组成变化对相应稳定性的影响,因为这些变化会对生态系统服务能力产生强大的影响[40]。本研究中,4个群落稳定性排序为阴坡>半阳坡>半阴坡>阳坡,从整体看,阴坡、半阳坡和半阴坡处于稳定状态,阳坡处于不稳定状态。这意味着祁连山山地草甸随阴坡-阳坡梯度转变会显著降低生态系统服务功能。其中,阳坡群落稳定性较差,这可能与阳坡的恶劣的生境条件有关,阳坡土地裸露率高,土壤蓄水能力差,养分和水分匮乏,植物聚集在局部适宜的的斑块中共享稀缺的资源,因此加剧了种内和种间竞争,导致群落稳定性较差。另一方面,我们发现阳坡群落受到一定程度的人为活动干扰,如祭扫和挖种植穴等,对群落稳定性产生不利影响,因此,应加强对山地草甸阳坡的保护与管理,诱导其进行顺向演替更新,进而提高植物群落的稳定性。
在本研究中,由于同一群落多样性大小与其稳定性并不存在相关关系,尚不清楚背后的生态学机制的相对贡献,这需结合更多生态影响因子(如太阳辐射,土壤温度等)深入分析对该各坡向群落稳定性的影响及机制,从而对祁连山山地草甸的植被恢复与重建、管理以及生物多样性保护等提供参考依据。
4 结论
山地草甸不同坡向草地植被特征和土壤含水量不同且差异显著,坡向影响草地生产力和群落稳定性,但对植被生物多样性无显著影响。Godron稳定性结果表明,阳坡相较于阴坡、半阴坡和半阳坡处于不稳定状态。同一群落多样性大小与其稳定性并不存在相关关系。建议在草地管理中考虑不同坡向草地植被特征变化,在禁牧的基础上,可采取适当的抚育措施(如灌溉、施肥和割草等),使植被进行正向演替更新,保障其生态恢复功能,提高山地草甸群落的生物多样性和稳定性,增加其生态系统服务能力。
