宁夏针茅属植物叶片碳稳定同位素特征及其影响因素
2022-08-31刘万弟李小伟
刘万弟, 李小伟
(宁夏大学农学院, 宁夏 银川 750021)
植物叶片δ13C值记录了植物生长过程中的环境因子信息,能够揭示植物对降水、热量因子等环境因子变化的响应[1-2]。由于该值易于测试,目前已经广泛用于植物水分利用效率、养分循环以及全球气候变化等研究领域,稳定同位素技术已成为研究植物与环境关系的有力工具[3]。叶片δ13C差异是由于植物光合作用对碳同位素的分馏所引起的,自身遗传特性是影响因素之一,例如:C4植物叶片δ13C值通常高于C3植物,而CAM植物则在二者之间[4-5]。另外,环境因子也是引起植物δ13C值变化重要原因[6],植物叶片光合作用和气孔导度受许多环境因子影响,造成细胞内与大气中CO2浓度比值的变化[1],如降水减少和温度增加会引起植物减少蒸腾作用,提高自身水分利用效率。
植物叶片δ13C响应环境梯度变化的规律能够反应植物的适应策略。但植物叶片δ13C与环境的关系尚没有统一的结论,比如:多数研究表明降水与植物δ13C呈负相关[7-8],但也有研究显示不同的结论,如在青藏高原草甸草原降水并不是植物δ13C的关键因子,而是温度[9];而更高海拔区域大气压强成为限制因素[9]。不同区域环境因子对植物δ13C影响不同,仍需更广泛的验证。此外,许多研究仅考虑了气候因素,未考虑土壤因子,如贺玲威等通过研究沙棘与气候的关系表明水蒸气压、生长季温度和太阳辐射是影响叶片δ13C的主要因素[10],而土壤水分对植物叶片δ13C影响较大[11],植物生长发育主要受土壤和气候共同影响,结合气候和土壤揭示植物δ13C对环境变化适应性至关重要。
宁夏草原地处干旱与半干旱过渡区,生态环境脆弱,南北水热梯度明显,针茅属植物作为建群种由旱生到湿生呈替代分布。鉴于δ13C是植物水分利用效率的指示指标,而水分是影响针茅属植物生长发育的限制因素之一[12-13]。在此背景下,探究针茅属植物叶片碳稳定同位素特征与气候和土壤因子的关系,可揭示针茅属植物δ13C值的环境变化响应机制,对针茅属植物响应未来气候变化研究具有重要意义。本研究通过测定6种针茅属植物叶片δ13C及环境因子数据,拟回答以下问题:(1)宁夏针茅属植物叶片δ13C基本特征;(2)宁夏针茅属植物叶片δ13C与环境因子间关系如何?(3)土壤因子和气候因子对δ13C的解释程度如何。
1 材料与方法
1.1 研究区域概况
在宁夏温带草原(35°14′~39°23′ N,104°17′′~107°39′ E)由北向南梯度上选择15个自然条件下以针茅属植物为优势种的样地,范围北至宁夏灵武市,南到宁夏固原市原州区,西至中卫沙坡头区,东为宁夏盐池县,涵盖了宁夏境内针茅属主要分布区域(图1)。宁夏属温带大陆性气候[14],生长季年降水量在141~249 mm,年均温在4.14~10.72℃之间,土壤以沙砾质及砾质土壤为主。

图1 样地调查分布Fig.1 Survey distribution of sample plot
1.2 研究方法
1.2.1植物取样 由于南北物候差异,采样于2016年7月中旬到8月中旬(植物生长旺季)进行,共15个样地,样地优势种针茅见表1。每个样地选取3个大样方,每个大样方内部取五个样本混合,每个样地按照五点取样法随机选择成熟完整无病斑的叶片,混为一个样本重复,3次重复。将叶片带回实验室,清洗表面灰尘杂质后,放入烘箱中,65℃条件下烘至恒重,研磨后过0.1 mm筛后进行碳稳定同位素测定。

表1 样地基本概况Table 1 Basic situation of the sample land
1.2.2碳稳定同位素测定 样品在EA-HT元素分析仪(Thermo Fisher Scientific,Inc.,Waltham,USA)中高温燃烧后生成CO2,通过DELTA V Advantage同位素比率质谱仪(Thermo Fisher Scientific,Inc.,Waltham,USA)检测CO2的13C与12C比率,并与国际标准物(Pee Dee Belnite,PDB)比对后计算出样品的δ13C值,计算公式如下:
δ13C=[(Rsa-Rst)/Rst]×1000‰
式中:Rsa和Rst分别为叶片样品以及标准样品的13C/12C值[15]。
1.2.3气候数据 本研究气候数据来自世界气候数据网站(www.worldclim.org),包括年均温度(Mean annual temperature,MAT)、年均太阳辐射(Solar radiation,SRAD),年均降水量(Mean annual precipitation,MAP),通过经纬度坐标提取每个样地的月份数据,并由此计算年均数据。
1.2.4土壤数据 在每个样地随机选取3个点采集0~40 cm土样混为一个样本,重复3次。土样带回实验室自然风干后,除去土壤中的细根及石块,研磨过0.15 mm筛后,用于土壤养分指标的测定:土壤全氮(Soil total nitrogen,TN)采用全自动凯氏定氮仪测定;土壤全磷(Soil total phosphorus,TP)采用HClO4-H2SO4法消煮后,采用钼锑抗比色法测定;土壤有机质(Soil organic matter content,SOC)采用重铬酸钾外加热法测定;土壤含水量(Soil water content,SWC)用差减法测定;土壤容重(Soil bulk density,BD)采用环刀法测定。
1.3 数据处理
使用Excel 2016和SPSS 22对数据进行整理和统计分析,使用单因素方差分析比较了不同针茅物种之间的碳稳定同位素差异,探究了针茅属植物叶片碳稳定同位素丰度的空间特征。通过Pearson相关分析了其与环境因子的关系;利用Canoco 5.0进行冗余分析(Redundancy analysis,RDA)以及蒙特卡洛检验,通过方差分解探讨了环境因子对碳稳定同位素δ13C影响的解释程度,通过蒙特卡洛检验筛选影响叶片δ13C的主要环境因素,绘图使用Origin 2021。
2 结果与分析
2.1 针茅属植物叶片δ13C基本特征
针茅属叶片δ13C平均值为-26.31‰,变化范围为-24.56‰~-28.07‰,变异系数为0.23~3.63%(表2、图2)。其中,戈壁针茅δ13C最大,沙生针茅δ13C最小,长芒草δ13C变异系数最小,短花针茅变异系数最大。沙生针茅和长芒草叶片δ13C与其他针茅间存在显著性差异(P<0.05)(表2)。
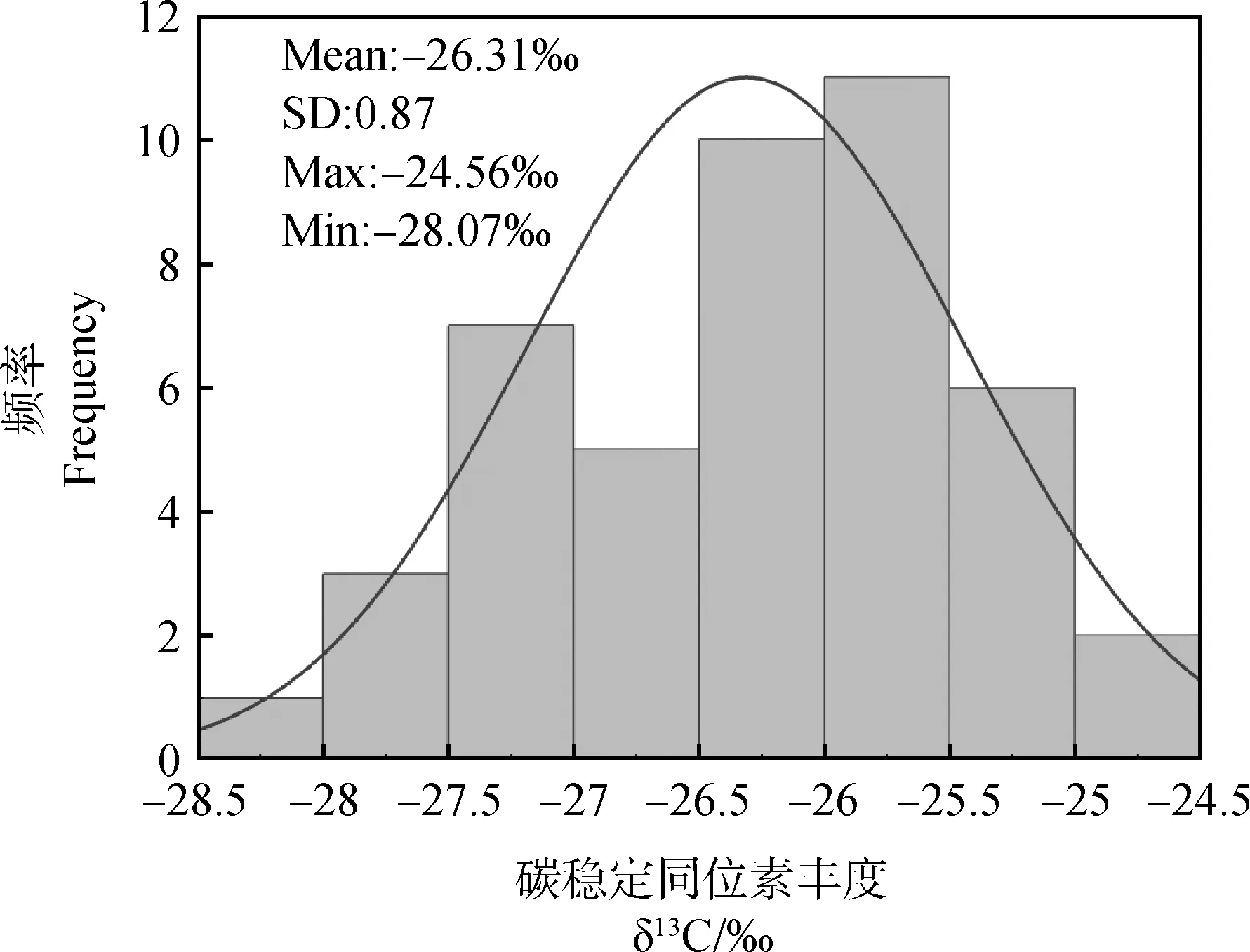
图2 针茅属植物叶片δ13C分布特征Fig.2 δ13C distribution trait in Stipa leaves注:Mean表示平均值;SD表示标准偏差;Max:表示最大值;Min表示最小值Note:Mean indicates the mean value;SD indicates the standard deviation;Max:indicates the maximum value;Min indicates the minimum value

表2 针茅属植物叶片δ13C基本特征Table 2 Basic characteristics of δ13C in Stipa leaves
2.2 针茅属植物叶片δ13C的空间特征
针茅属植物叶片δ13C与经度呈显著负相关(P<0.01,R2=0.38);在纬度梯度上,随纬度增加呈先增加后减小的单峰趋势(P<0.01,R2=0.49);在海拔梯度上,随海拔增加呈先增加后减小的单峰趋势(P<0.01,R2=0.46)。

图3 针茅属植物叶片δ13C与经度、纬度、海拔的关系Fig.3 Correlation of Stipa leaf δ13C with longitude,latitude and altitude
2.3 针茅属植物叶片δ13C与环境因子的关系
如图4所示,针茅属植物叶片δ13C与年均温(MAT)、太阳辐射量(SRAD)、容重(BD)极显著负相关,与土壤全磷(TP)极显著正相关;而与年均降水量(MAP)、土壤有机质(SOC)、土壤全氮(TN)、土壤含水量(SWC)相关性不显著。


图4 针茅属植物叶片δ13C与环境因子的关系Fig.4 The relationship between leaf δ13C and environmental factors in Stipa
2.4 环境因子对针茅属植物叶片δ13C的综合影响
为了分析土壤和气候因子对针茅属植物叶片δ13C的影响程度,我们对环境因子和δ13C进行方差分解分析。环境因子总共解释了74.58%的方差,其中土壤因子单独解释量为32.87%,气候因子单独解释了7.54%,二者共同解释量为34.17%,表明土壤较气候影响更大。

图5 环境因子对δ13C方差解释量Fig.5 Amount of variance explained by environmental factors on δ13C
如图6所示,通过冗余分析排序图,环境梯度在RDA轴1正方向呈热量增加,土壤容重越大,环境干旱胁迫越大;负方向土壤养分呈增加趋势。整体来说,针茅叶片碳稳定同位素丰度与土壤养分呈正相关,与年均温、太阳辐射和土壤容重呈负相关,与水分因子相关性较弱。

图6 不同针茅叶片δ13C与环境的排序图Fig.6 Sequencing diagram of δ13C of different Stipa with environment注:1. 短花针茅,用圆圈表示;2. 沙生针茅,用正方向表示;3. 长芒草,用三角形表示;4.戈壁针茅,用星形表示;5.大针茅,用十字表示;6. 甘青针茅,用叉表示Note:1. S. breviflora,represented by circle;2. S. caucasica subsp. Glareosa,represented by positive direction;3. S.bungeana,represented by triangle;4. S. tianschanica var. gobica,represented by star;5. S. grandis,represented by cross;6. S. przewalskyi,represented by X mark
通过蒙特卡洛检验对环境因子进行筛选,结果表明:土壤全磷(TP)、土壤容重(BD)、年均温(MAT)、太阳辐射量(SRAD)对针茅属植物叶片δ13C贡献率最大(P<0.05),且依次减小(表3)。

表3 蒙特卡洛检验筛选结果Table 3 Screening results of Monte Carlo test
3 讨论
3.1 针茅属植物叶片δ13C基本特征
本研究中,6种针茅属植物叶片δ13C范围为-28.07‰~-24.56‰,均值为-26.31‰。植物存在三种光合途径(C3、C4、CAM),由于植物光合作用在固定CO2产生碳同位素分馏[4],光合羧化酶和羧化的时空差异对13C有不同的识别和排斥,使得δ13C存在差异,因此可以用于区分植物光合途径类型[5]。陆地植物中,C3植物δ13C为-35‰~20‰,均值为-26‰;C4植物δ13C为-15‰~7‰;均值为-12‰,CAM植物δ13C为-22‰~10‰,均值为-16‰[5],因此,这6种针茅属植物均为C3植物。本研究δ13C均值为-26.31‰,略低于全球陆生C3植物,其原因可能为我们的研究对象均为草本,而通常来说不同生活型植物δ13C表现为草本<灌木<乔木[16]。δ13C变异系数范围为0.23%~3.63%,其中短花针茅变异系数最大(3.63%),而沙生针茅和长芒草变异系数较低(0.40%,0.23%)。这和针茅属植物不同生态适应性有关,短花针茅是温性荒漠草原的主要建群种,也可生长于较为湿润的典型草原[17]。在我们选取样地调查过程中,短花针茅分布范围最广(38°05′14″ N~36°39′55″ N),干旱环境到较湿润区域均有分布(图1、图6),能够适应多种环境;长芒草通常被认为是温性典型草原优势种,但由于天然植被遭受破坏,大面积的长芒草群落较为稀少[17],在我们的样地中长芒草样地处于荒漠草原区域;沙生针茅分布于沙地,分布范围狭窄。Zhao等对青藏高原针茅属植物大范围调查研究发现,分布较广的针茅环境适应性较高[18];Chen等对不同生境下的黄囊苔草种群研究表明,能够通过改变其水分利用效率适应不同的土壤水分状况,使其在植物群落中表现出更强的竞争能力和生态适应性[19],因而植物对环境适应力是影响针茅属植物δ13C的重要因子。
3.2 针茅属植物叶片δ13C的空间特征
针茅属植物叶片δ13C与经度呈极显著负相关,通过Pearson相关可知,经度与年均温和太阳辐射显著正相关,热量和太阳辐射是影响植物光合作用的重要因子,从而引起叶片δ13C差异。δ13C随纬度增加先上升后下降,研究区域随纬度增加,降水减少年均温与太阳辐射量增加,因而δ13C呈增加趋势。而较高纬度针茅叶片δ13C降低,其原因为,样地3(长芒草)和4(短花针茅)当年降水较充足,δ13C与降水呈负相关;样地1(短花针茅)和2(沙生针茅)分布于沙地,生长周期较短,而通常来说,叶片δ13C与植物寿命呈正比[20]。针茅属植物叶片δ13C随海拔增加呈先上升趋势后下降,这与大多数研究不完全一致,郑淑霞等通过文献综述得出结论相近:全球高山地带植物δ13C与海拔呈正相关[3]。海拔会重塑水热条件,海拔升高通常伴随着温度下降、水分增加,温度较低会使植物部分气孔关闭,光合作用过程对13CO2识别减少,从而增加δ13C[21],同时随着海拔升高,大气中CO2分压降低,植物对13C的吸收增加[11],这也会使δ13C增大;而在较高海拔区域(本研究为样地15,海拔2 462.52 m),降水增加,这使得叶片δ13C降低。
3.3 针茅属植物叶片δ13C与水分的关系
通常来说植物叶片δ13C与土壤水分和降水量呈负相关[22],当土壤中可利用水分较少时,植物会通过自身生理调节,如关闭部分气孔[23],从而降低水分消耗,提高水分利用效率,使得叶片δ13C变大。本研究中土壤水分和降水量均与叶片δ13C相关性不显著,呈弱正相关,这可能与禾本科植物较短的生活周期及自身特性有关;最近研究表明:黄土高原C3植物(包括禾草、灌木、乔木)叶片δ13C与降水量呈显著负相关,但禾草类植物叶片δ13C与降水量相关性不显著[20],这是因为禾本科植物为须根系,依赖于土壤表层水分,短期降水影响较大,而乔本和灌木植物则可以利用深层土壤水分。其次,随着纬度增加,宁夏年均温越高降水越少,生长环境越恶劣,针茅属植物采用快速生长策略,生长周期较短,而通常来说,叶片δ13C与植物寿命呈正比[16,20],因此叶片δ13C与水分关系不显著。相反,寿命较长的植物经历干旱的机会也越多,其水分利用效率越高,因而寿命长的植物叶片δ13C与水分相关性更为显著[24]。另一方面可能与我们的采样时间有关,我们采集叶片时间为7月中旬到8中旬,宁夏属于温带大陆性气候,雨热同期,生长季降水的补充使得δ13C与水分相关性不显著[25]。
3.4 针茅属植物叶片δ13C与环境因子的关系
总体来看,土壤因子对宁夏针茅属植物叶片δ13C方差的解释量大于气候因子。植物叶片δ13C差异是由于光合作用对碳原子的选择性吸收,影响光合作用的因素(土壤养分、水分、温度等)均会影响叶片δ13C[26-27],如土壤养分条件越好,对植物生长和光合作用越有利,从而使叶片δ13C增高[23]。通过计算土壤因子和气候因子的变异系数以了解梯度差异,结果显示,土壤变异系数为13.71%~83.01%,气候变异系数为3.35%~22.57%,由此可见,在环境梯度差异上土壤因子远大于气候因子,尽管太阳辐射是植物光合作用能量主要来源,但植物光合作用需要的水分及养分绝大部分来源于土壤,因而土壤很大程度决定着植物叶片δ13C值。
本研究中土壤全磷(TP)是影响针茅属植物叶片δ13C最重要的因子,呈显著正相关。在一定范围内,叶片矿质元素含量增加能够提升光合速率,降低CO2分压,从而使得δ13C变大[27];另一方面植物体内磷累积能够增加体内束缚水的含量,增加抗旱性,以保障干旱条件下的生长发育[28],提高了水分利用效率,而叶片δ13C与水分利用效率(WUE)呈正相关[29]。同时,磷是核酸、核苷酸、辅酶等的组成元素,参与植物ATP的反应关键之一[27],而宁夏天然草地植物受土壤磷限制[30];通过纬度和土壤磷含量的关系可得,研究区域由北向南土壤磷含量增加,提升了植物光合作用与水分利用效率,使得δ13C增大。另一方面,宁夏由南向北土壤容重(BD)呈增加趋势[25],伴随着土壤保水保肥能力下降,且降水不足,植物对水分保存尤为重要,而针茅属植物属于须根系植物,许多研究表明须根系植物具有较好的保水能力[31-32],且随着土壤容重减少而增大,因而土壤容重减小针茅根系水分保存及供给越好,提高了水分利用效率[33],叶片δ13C增大。
本研究针茅属植物叶片δ13C与年均温(MAT)、太阳辐射(SRAD)显著负相关,这和任书杰等人研究一致[29]。而大多数研究表明年均温与叶片δ13C正相关[34],如贺玲威等对沙棘属植物叶片δ13C与年均温显著正相关[10],这是由于干旱胁迫,植物通过关闭气孔减少蒸腾作用水分散失,这使得植物对13CO2识别度下降,因而δ13C变大。宁夏由南向北,年均温(MAT)和太阳辐射(SRAD)均呈增加趋势[25],且降水较少,干旱胁迫明显,而针茅属植物在应对干旱胁迫时仍能维持较高的光合系统活性,通过调节气孔调控光合所需的CO2和可利用水资源之间的折中点,能够提高自身水分利用效率,同时降低对碳的吸收[35-36];另一方面较高的温度和太阳辐射使植物光合羧化酶的活性增强,对13CO2识别度增加,δ13C值降低[37]。
4 结论
综上所述,宁夏温性草原针茅属植物叶片δ13C范围为-28.07‰~-24.56‰,均值为-26.31‰,为C3植物。叶片δ13C与经度呈负相关,随纬度和海拔呈先增加后下降的趋势;与水分相关性不显著,土壤因子对叶片δ13C影响大于气候,土壤磷含量、土壤容重、年均温和太阳辐射量是影响叶片δ13C最主要因子,其中土壤磷影响最大,在之后的研究中应多关注土壤磷对植物的影响。本研究结果对揭示温性草原针茅属植物响应气候变化的适应策略,并为大尺度研究提供基础数据支持。
