小亚璃眼蜱CPL截短基因的多克隆抗体制备及其应用研究
2022-08-24翟雪洁郝蕴伟阿力木江加帕尔宋瑞其诺明达来何文文郑会珍巴音查汗
翟雪洁,郝蕴伟,阿力木江·加帕尔,宋瑞其,诺明达来,蒋 倩,何文文,郑会珍,巴音查汗*
(1.新疆农业大学动物医学学院,新疆 乌鲁木齐 830052;2.石河子大学医学院基础医学系,新疆 石河子 832003)
硬蜱是一种可以通过吸食宿主血液传播疾病的体外寄生虫。小亚璃眼蜱(Hyalomma anatolicum,H. anatolicum)属于硬蜱科璃眼蜱属,其幼蜱和若蜱主要寄生于啮齿类动物,成蜱主要寄生于骆驼、牛、马、羊等家畜。H.anatolicum在国外主要分布于印度、巴基斯坦、伊朗、苏丹、以色列等地[1-2],国内仅在中国新疆有报道,其是新疆的优势蜱种[3-4],其除传播动物梨形虫[5],还可传播克里米亚-刚果出血热、立克次体病、莱姆病和森林脑炎等人畜共患病的病原[6-7]。防治蜱虫主要使用有机磷和拟除虫菊酯类化学药物,但有研究显示因滥用杀虫剂使得蜱虫对部分化学药物产生了耐药性,且耐药性逐年增强[8-9],而抗蜱疫苗可有效避免环境污染和蜱虫耐药性的产生,因此筛选鉴定抗蜱疫苗候选分子就显得尤为重要。
组织蛋白酶L(Cathepsin L,CPL)是木瓜蛋白酶家族中的半胱氨酸蛋白水解酶,有研究显示CPL 在长角血蜱(Haemaphysalis longicornis)和蓖子硬蜱(Ixo-des ricinus)中参与多酶级联反应等过程[10],沉默CPL基因会影响H. longicornis中天冬氨酸蛋白酶(Longep-sin)、天冬酰胺内肽酶(HlLgm-1 和HlLgm-2)和丝氨酸羧肽酶(HlSCP-1)等的表达,从而使雌蜱的饱血重量下降[11],所以CPL 是具有研究价值的抗蜱疫苗候选分子[12-13]。基于此,本研究对H. anatolicum的CPL 截短基因(HanCPL)进行克隆,并使用原核、真核系统分别表达出了原核重组HanCPL 蛋白(rHanC-PL)和真核重组CPL 蛋白(EGFP-HanCPL)。利用western blot 对rHanCPL 和EGFP-HanCPL 鉴定,并通过免疫组化试验和间接免疫荧光试验(IFA)对CPL 定位,为后续基于rHanCPL 为候选抗原的抗小亚璃眼蜱的亚单位疫苗研发提供参考依据。
1 材料与方法
1.1 主要实验材料新西兰兔购自新疆医科大学实验动物中心。H. anatolicum、pET-28a、pEGFP-C3载体和HEK-293T 细胞均由新疆农业大学动医学院寄生虫实验室提供;FastKing cDNA 第一链合成试剂盒、PCR 相关试剂、BL21(DE3)感受态细胞均购自天根生化科技(北京)有限公司;TRIzol 试剂、EcoR I、XhoI、T4 DNA连接酶、pMD19-T载体和DH5a感受态细胞均购自TaKaRa 公司;兔抗His tag、HRP 标记羊抗兔IgG H&L 购自北京博奥森生物技术有限公司;Cy3 标记羊抗兔IgG(H+L)购自艾克索生物科技股份有限公司。
1.2 引物设计与合成利用Antibody Epitope Predic-tion(http://tools.immuneepitope.org/bcell/)对本实验室前期获得的H. anatolicum转录组数据中的CPL 基因序列进行B 细胞抗原表位预测,从中选取B 细胞抗原表位较多的氨基酸区域对应的碱基序列,利用Prim-erPremier5.0 和Oligo 6.24 软 件 设 计HanCPL 截 短 基 因扩增引物,引物由上海生工生物工程技术服务有限公司合成,具体引物序列信息见表1。
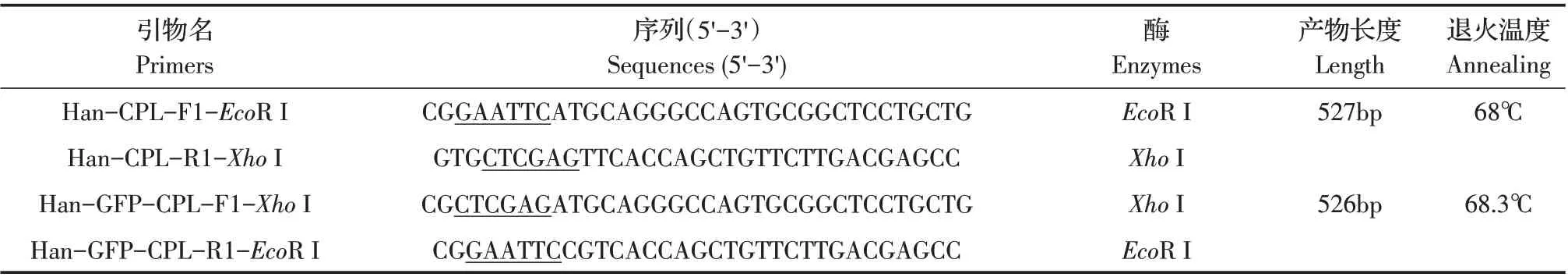
表1 HanCPL截短基因扩增的引物信息Table 1 HanCPL truncated gene prokaryotic and eukaryotic expression primer information
1.3 HanCPL 截短基因的PCR 扩增及表达载体的构建取吸血后第5 d 半饱血雌性H. anatolicum成蜱,用预冷PBS 清洗后放入无菌袋,置于液氮中研磨,利用TRIzol提取蜱虫总RNA,利用FastKing cDNA 第一链合成试剂盒将RNA 反转录为cDNA,以其为模板,采用引物Han-CPL-F1-EcoR I/Han-CPL-R1-XhoI 和Han-GFP-CPL-F1-XhoI/Han-GFP-CPL-R1-EcoR I,经PCR 扩增CPL 截短基因(HanCPL),扩增产物由上海生工生物工程技术服务有限公司测序鉴定。PCR 反应条件为:94 ℃4 min,94 ℃30 s、68 ℃/68.3 ℃30 s、72 ℃45 s,35 个循环,72 ℃10 min。PCR 产物纯化回收测序正确后分别连接至pET-28a、pEGFP-C3中,构建重组质粒pET-28a-HanCPL 和pEGFP-C3-HanCPL,重组质粒经双酶切(EcoR I/XhoI)鉴定正确后备用。
1.4 rHanCPL 表达及纯化的检测将重组质粒pET-28a-HanCPL 转 化 至BL21(DE3)感 受 态 细 胞 中,37 ℃180 r/min 培养至OD600nm为0.4~0.6 时加入终浓度0.8 mmol/L IPTG 诱导6 h,收集菌体超声破碎后,分别收集上清和沉淀,利用SDS-PAGE 检测rHanCPL 的表达情况。利用His Trap 亲和层析柱纯化,再利用尿素透析法复性后,通过SDS-PAGE 分析rHanCPL 的纯化情况;进一步以兔抗His tag(1∶5 000)为一抗,HRP 标记羊抗兔IgG H&L(1∶10 000)为二抗,进行western blot 鉴定,并采用紫外分光光度计测定蛋白浓度后置于-80 ℃保存。
1.5 rHanCPL多克隆抗体的制备及效价检测将6只新西兰兔随机均分为2 组,取200 μg rHanCPL 与等体积氢氧化铝佐剂乳化后,采用背部皮下多点注射免疫实验组兔,对照组兔注射等体积的PBS 混合氢氧化铝佐剂,免疫后间隔14 d二次免疫。二免后14 d经耳缘静脉采血,分离血清制备rHanCPL 多克隆抗体。以制备的rHanCPL 多克隆抗体(1∶100)为一抗,HRP 标记羊抗兔IgG H&L(1∶10 000)为二抗,进行western blot检测。进一步以纯化的rHanCPL(2 μg/mL)为包被抗原,以2 倍倍比稀释(1∶200~1∶409 600)的rHanCPL多克隆抗体为一抗,同时以对照组兔血清作为阴性对照,以HRP 标记羊抗兔IgG H&L(1∶10 000)为二抗,采用间接ELISA 检测rHanCPL 多克隆抗体效价。
1.6 天然CPL 蛋白在半饱血H. anatolicum组织中的定位检测 将半饱血H. anatolicum雌蜱按常规方法制成石蜡切片,脱蜡后利用柠檬酸抗原修复缓冲液修复抗原,然后置于3%过氧化氢溶液中阻断内源性过氧化物酶,3% BSA 中封闭后,以制备的rHanC-PL 多克隆抗体(1∶500)为一抗, HRP 标记的羊抗兔IgG(1∶1 000)为二抗,经免疫组化(IHC)检测半饱血H.anatolicum中天然CPL 蛋白的定位情况。
1.7 HEK-293T 细胞中EGFP-HanCPL 表达检测将HEK-293T 铺于96 孔细胞培养板,待细胞密度达60%左右,使用lipofectamine2000 试剂分别将pEGFPC3 和pEGFP-C3-HanCPL 转染至HEK-293T 细胞,于37 ℃、5% CO2培养48 h 后随机收集各孔中的细胞,超声破碎后,于4℃离心收集上清液,采用1.4 中的westen blot 方法检测EGFP-HanCPL 蛋白与rHanCPL多克隆抗体的反应原性,试验设转入pEGFP-C3 的细胞裂解液作为阴性对照。培养24 h 后使用4%多聚甲醛固定各组细胞,以制备的rHanCPL 多克隆抗体(1∶100)为一抗,以Cy3 标记的羊抗兔IgG(H+L)(1∶400)为二抗,置于倒置荧光显微镜观察,采用Adobe Photoshop 2020 进行Merge 处理,经IFA 检测rHanCPL 多克隆抗体与真核表达EGFP-HanCPL 蛋白的反应性及其在细胞中的定位。
2 结 果
2.1 重组表达载体的鉴定结果PCR 扩增HanCPL结果显示,扩增到约530 bp 的目的片段,与预期相符(图略),经测序比对,结果显示其与小亚璃眼蜱源靶基因(KC707940)HanCPL 同源性99.60%,表明扩增获得HanCPL。构建pET-28a-HanCPL 和pEGFPC3-HanCPL重组质粒,经双酶切鉴定插入基因片段大小均与预期一致(图1),表明重组质粒pET-28a-HanCPL和pEGFP-C3-HanCPL正确构建。

图1 pET-28a-HanCPL和pEGFP-C3-HanCPL的双酶切鉴定Fig.1 Double enzymes digestion identification of pET-28a-HanCPL and pEGFP-C3-HanCPL
2.2 rHanCPL 表达、纯化的检测结果重组质粒pET-28a-HanCPL 转化至BL21(DE3)感受态细胞,采用IPTG 诱导rHanCPL 的表达,经SDS-PAGE 后结果显示,在27 ku 处出现与理论值相符的目的条带,且rHanCPL 以包涵体形式表达,经His Trap 亲和层析柱纯化后利用尿素透析法复性,得到了高纯度的rHanCPL,浓度为3 mg/mL。经western blot 分析结果显示,rHanCPL 可以被兔抗His tag 抗体特异性识别,且表达蛋白大小与预期相符(图2)。结果表明正确表达且得到了高纯度rHanCPL。

图2 rHanCPL表达及纯化的鉴定结果Fig.2 The identification results of rHanCPL expression and purification
2.3 rHanCPL 多克隆抗体的制备及效价检测结果经western blot 分析制备的rHanCPL,结果显示,rHanCPL 可以被制备的rHanCPL 多克隆抗体特异性识别,而对照组的兔血清不与rHanCPL 反应,表明rHanCPL 具有较好的反应原性(图3A)。进一步以纯化的rHanCPL 为包被抗原,利用间接ELISA方法测定rHanCPL 多克隆抗体效价,结果显示,rHanCPL 多克隆抗体OD450nm值/免疫PBS 血清OD450nm>2.1的最大稀释倍数达1∶204 800(图3B),可用于后续试验。

图3 rHanCPL的western blot鉴定及多克隆抗体的效价测定Fig.3 Western blot identificantion and determination of rHanCPL polyclonal antibody titer
2.4 天然CPL 蛋白在半饱血H. anatolicum组织中的定位检测结果 半饱血H.anatolicum雌蜱组织经IHC检测结果显示,染色反应分布于中肠肠腔的中肠上皮细胞内,并呈棕黄色或褐色(图4,白色箭头),细胞核呈蓝色(图4,黑色箭头)。表明rHanCPL 多克隆抗体可以与H. anatolicum的CPL 天然蛋白发生反应,且其定位于H.anatolicum的中肠上皮细胞。

图4 H.anatolicum中肠组织中CPL蛋白定位的IHC结果(40×)Fig.4 Immunohistochemical results of H.anatolicum midgut tissue(40×)
2.5 EGFP-HanCPL 在细胞中表达及定位的检测结果为了解真核表达的重组蛋白EGFP-HanCPL 在真核细胞中的表达和定位情况,收集分别转染质粒pEGFP-C3-HanCPL和空载体pEGFP-C3的HEK-293T细胞,western blot检测结果显示,rHanCPL多克隆抗体可以识别EGFP-HanCPL蛋白,出现特异性条带,并且空载体表达的EGFP蛋白无特异性条带(图5A)。IFA检测结果显示,EGFP蛋白呈绿色荧光,与rHanCPL多克隆抗体不发生反应,DAPI染核后呈蓝色荧光,两者不完全重合;而表达的EGFP-HanCPL呈现的绿色荧光与标记IgG 的Cy3 红色荧光完全重合,与DAPI 染核后的蓝色荧光不完全重合(图5B)。上述结果表明EGFPHanCPL可以与rHanCPL 多克隆抗体特异性结合,并主要定位于细胞质。

图5 EGFP-HanCPL蛋白的western blot(A)和IFA(B)检测结果Fig.5 Western blot(A)and IFA(B)detection results of EGFP-HanCPL protein
3 讨 论
硬蜱在饱血后很长一段时间不进食,在此期间,蜱虫通过消化所摄取的宿主血液为其提供生存、繁殖所必需的能量[14],因此蜱类的血餐消化系统是否正常运行显得非常重要。而蜱中肠中血红蛋白的水解是由多种蛋白酶协同作用的结果[15]。已有研究发现丝氨酸蛋白酶、半胱氨酸蛋白酶和天冬氨酸蛋白酶等参与了硬蜱消化血液的级联反应[16-18]。其中CPL 作为一种半胱氨酸蛋白酶则是参与宿主血液消化级联反应中一个重要的蛋白酶,抑制CPL 可能会影响蜱的吸血、消化血液能力,从而可以减少蜱虫种群规模,甚至降低蜱传病的感染和传播[11]。因此,本研究对新疆优势蜱种H.anatolicumCPL 的截短基因进行了克隆,构建重组质粒pET-28a-HanCPL 和pEGFP-C3-HanCPL。利用原核表达系统获得高表达量的rHanCPL,使用纯化后的rHanCPL 免疫新西兰兔制备rHanCPL 多克隆抗体,在二免后14 d的兔血清效价高达1∶204 800,说明rHanCPL进入兔体后能够有效刺激其免疫系统快速产生免疫应答,证明rHanCPL 具有良好的免疫原性,并且经western blot 鉴定,rHanC-PL 多克隆抗体可与rHanCPL 和EGFP-HanCPL 发生特异性反应,证明rHanCPL 和EGFP-HanCPL 均具有良好的反应原性。
研究发现,在I. ricinus中IrCL1 主要参考半饱血雌蜱的肠道血红蛋白分解[19],本研究利用rHanCPL 多克隆抗体通过IHC 试验发现CPL 同样存在于H.anato-licum半饱血雌蜱的中肠上皮细胞中,提示HanCPL可能在H. anatolicum雌蜱的血餐消化中起到重要作用。本研究进一步将pEGFP-C3-HanCPL 转染至HEK-293T 细胞,通过IFA 试验对EGFP-HanCPL 产生的荧光信号进行分析,结果表明EGFP-HanCPL 主要表达于细胞质,与前人在微小牛蜱(Rhipicephalus microplus)中对CPL 的定位结果一致[20]。
本研究克隆了CPL 截短基因,并利用原核、真核系统对其进行了表达,纯化后制备rHanCPL多克隆抗体,应用于IHC、IFA 和Western blot 检测,证明了rHanCPL具有良好的抗原性,为进一步分析rHanCPL作为H.anatolicum抗蜱疫苗提供参考依据,但rHanCPL在作为疫苗候选抗原的安全性以及rHanCPL 多克隆抗体的抗蜱保护效果方面还有待进一步研究。
