崂山奶山羊无角间性综合征生殖缺陷基因的基因型分析
2022-08-23张权威都萌萌戴正浩刘开东林晓坤高而尚秦至莉赵金山李和刚
张权威,都萌萌,程 明,戴正浩,刘开东,林晓坤,高而尚,秦至莉,赵金山,李和刚
(1.青岛农业大学动物科技学院,青岛 266109;2.青岛市畜牧兽医研究所,青岛 266100)
羊角是羊的标志性特征之一,但并非经济性状。在野生情况下,羊角是羊为了吸引配偶、争夺地位等进行争斗的重要工具。在现代畜牧业养殖中,有角山羊打斗明显且占有优势地位,饲喂时抢料严重,而且打斗还会造成流产甚至骨折、乳房撕裂等情况,给饲养管理造成极大的困难,从而增加经济成本。而无角羊与有角羊相比不仅减少了个体之间相互争斗带来的经济损失,而且也有利于对家畜的大批量圈养,更利于养殖和管理。因此,培育无角羊品系已经成为羊育种发展的趋势。
据报道,松弛素家族肽受体2(relaxin family peptide receptor 2,RXFP2)是影响绵羊角的有无、大小和角型的主要候选基因[1-4],关于绵羊多角性状的候选基因较多,主要有同源框基因家族D(homeobox gene family D,HOXD)和RXFP2基因等[5-8]。在山羊中同样存在无角变异,但与绵羊不同的是山羊的无角性状与间性性状是紧密连锁的,称为无角间性综合征(polled intersex syndrome,PIS)。出现这种现象的原因是由于山羊1号染色体上约129 Mb处的11.7 kb缺失,并影响叉头转录因子2(fokhead box L2,FOXL2)和PIS区调控的第一个转录产物(PIS regulated transcript number 1,PISRT1)的转录造成的[9]。有学者认为间性奶山羊中11.7 kb片段的缺失不是完全的而是部分的[10-11],其中198 bp替换和108 bp缺失也可能引发奶山羊的间性[10]。Simon等[12]研究揭示了一种更为复杂的结构变异,认为间性奶山羊的出现是由于1号染色体上约129 Mb区域总长度为约10.1 kb的缺失和1个反向插入的约480 kb大小的重复片段导致的。尽管目前的研究定位了山羊PIS基因的可能突变区,但具体作用机制还不清楚,严重阻碍了无角山羊的品种培育进程。因此,本试验以崂山奶山羊为研究对象,分析崂山奶山羊PIS基因的遗传变异特点,为培育崂山奶山羊无角新品系提供参考。
1 材料与方法
1.1 样品采集
采集青岛地区155只崂山奶山羊全血样品,其中间性山羊36只,有角公羊9只,有角母羊34只,无角母羊29只,无角公羊47只,用血液基因组DNA提取试剂盒(天根生化科技(北京)有限公司)提取血液样品基因组DNA。
1.2 方法
1.2.1 间性山羊的表型特征分析 根据36只间性崂山奶山羊外生殖器的形态特点,对其进行形态学分类。
1.2.2 引物设计及合成 本研究涉及的引物共分为3部分:遗传性别判定引物、PIS区198 bp替换和108 bp缺失的基因型鉴定引物以及PIS区10.1 kb缺失/480 kb重复的基因型鉴定引物。遗传性别判定引物的设计参考性别决定区Y基因(sex determination region Y,SRY)序列(JN561348);内参引物则根据GAPDH基因序列(NM_001190390)设计。PIS区198 bp替换和108 bp缺失的基因型鉴定引物PIS wt-1则参考Li等[10]11.7 kb片段变异检测引物,其验证引物是在PIS区设计1对包含PIS wt-1的引物PIS wt-3;PIS区10.1 kb缺失/480 kb重复的基因型鉴定引物则根据山羊1号染色体PIS区10.1 kb缺失/480 kb重复特征,结合NCBI公布的山羊基因组DNA的完整序列信息(RefSeq:NC_030808.1)进行设计。利用 Primer Premier 5.0 软件设计引物。各鉴定引物在1号染色体上的位置如图1所示,引物序列信息见表1。引物均由生工生物工程(上海)股份有限公司合成。

图1 PIS基因型鉴定引物在1号染色体上的位置Fig.1 Location of PIS genotype identification primers on chromosome 1
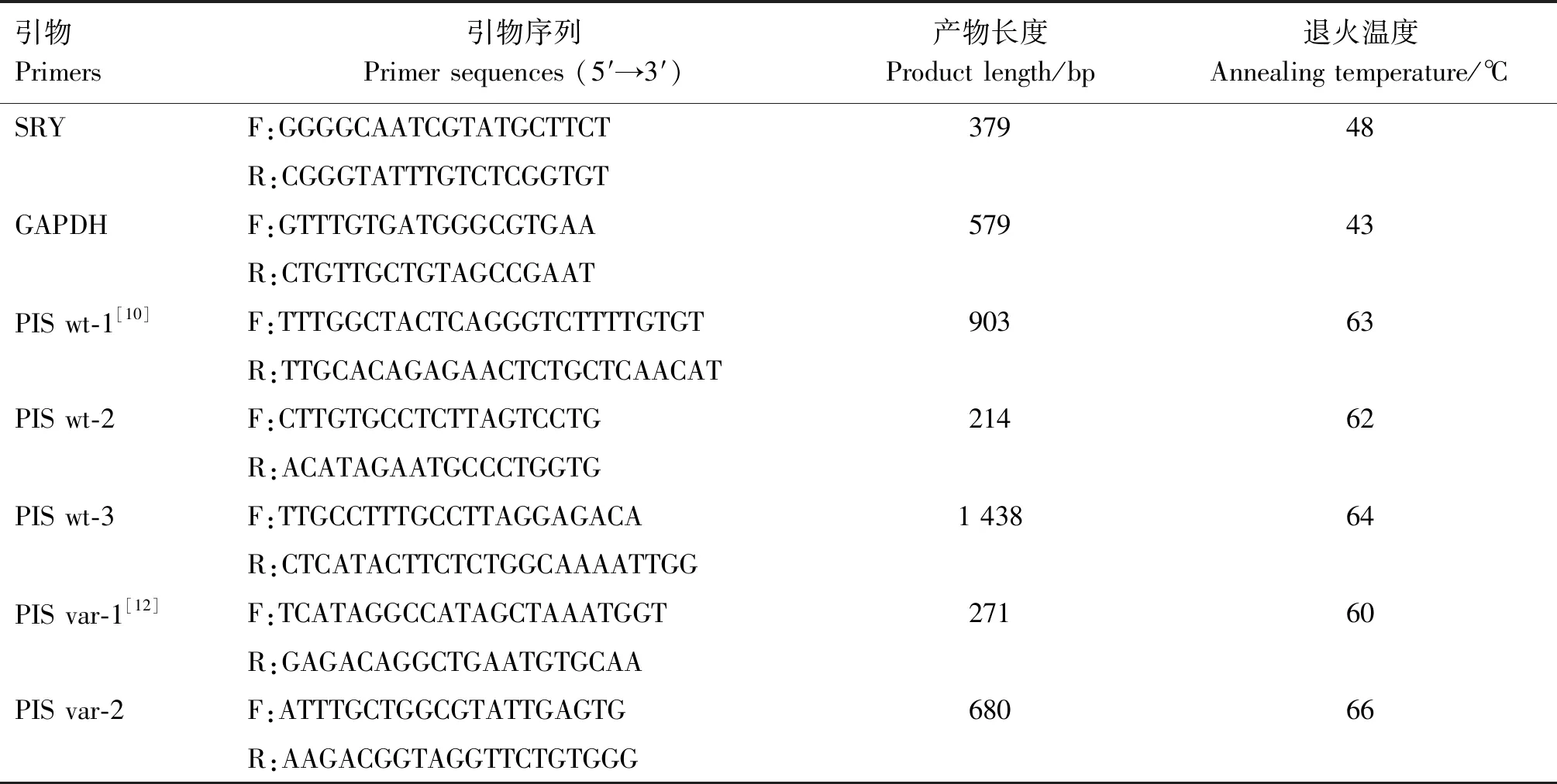
表1 引物信息
1.2.3 遗传性别鉴定 PCR扩增SRY和GAPDH基因片段,通过检测2个特异性片段的有无确定155只山羊的遗传学性别。2个特异性片段(379、579 bp)同时存在则为雄性,只存在579 bp则为雌性。PCR扩增体系25 μL:1.1×Golden Star Super PCR Mix 22 μL,DNA 1 μL,上、下游引物(SRY-F/R、GAPDH-F/R)各1 μL。PCR反应程序:95 ℃预变性5 min;95 ℃变性30 s,退火30 s(退火温度见表1),72 ℃延伸30 s,共34个循环;72 ℃延伸5 min;4 ℃保存。2.0%琼脂糖凝胶电泳检测PCR产物。
1.2.4 PIS区198 bp替换和108 bp缺失的基因型鉴定 利用PIS wt-1检测有角、无角和间性山羊,分析间性山羊在129 427 003~129 427 905 bp区域是否存在198 bp替换和108 bp缺失;同时利用PIS wt-3验证山羊PIS区是否存在不完全缺失。PCR扩增体系10 μL:1.1×Golden Star Super PCR Mix 8.8 μL,DNA 0.4 μL,上、下游引物(PIS wt-1-F/R、PIS wt-3-F/R)各0.4 μL。PCR反应程序:95 ℃预变性5 min;95 ℃变性30 s,退火30 s(退火温度见表1),72 ℃延伸1 min,共34个循环;72 ℃延伸5 min;4 ℃保存。1.5%琼脂糖凝胶电泳检测PCR产物。
1.2.5 PIS区10.1 kb缺失/480 kb重复的基因型鉴定 PIS区10.1 kb缺失/480 kb重复的基因型鉴定具体涉及3对引物(PIS wt-2-F/R、PIS var-1-F/R、PIS var-2-F/R),通过检测3个特异性片段的有无来确定155只山羊的PIS基因型。扩增PIS var-1和PIS var-2片段是为了验证山羊PIS基因的缺失类型。不同基因型山羊的扩增片段组合如表2所示。PCR扩增体系及反应程序同1.2.3。

表2 3种PIS生殖缺陷基因型的片段大小
2 结 果
2.1 间性山羊的表型特征分析
按照间性山羊外生殖器的形态特点可将其分为3种类型:①大阴蒂雄性假间性有17只,其阴户缩小且阴蒂增大而突出阴户之外,性腺为睾丸组织;②拟雌性假间性有12只,其阴蒂增大,状如阴茎,阴户缩小严重且与肛门之间的距离增大;③短阴茎间性有7只,其外表为雄性,但阴茎发育不全,内部有两性生殖管道与睾丸。
2.2 遗传性别鉴定
由图2可知,正常母羊的扩增产物只有1条579 bp条带,正常公羊的扩增产物同时具有579和379 bp 2条带。间性山羊中只检测出579 bp条带,并未检测到379 bp条带,说明间性山羊的性染色体为XX。

M,DL2000 DNA Marker;1、2,有角母羊;3,有角公羊;4、5,无角母羊;6,无角公羊;7~9,间性山羊M,DL2000 DNA Marker;1 and 2,Horned ewes;3,Horned rams;4 and 5,Hornless ewes;6,Hornless rams;7-9,Intersex goats图2 遗传性别判定结果电泳图Fig.2 Electrophoretic diagram of genetic sex determination results
2.3 PIS区198 bp替换和108 bp缺失的基因型鉴定
利用PIS wt-1检测无角山羊(图3A)和有角山羊(图3B)都显示出903和713 bp条带,而间性山羊只显示出713 bp条带(图3B)。利用PIS wt-3检测间性山羊时并未检测出1 438 bp条带(图4),说明在1号染色体129 427 003~129 427 905 bp区域并不存在198 bp替换和108 bp缺失。
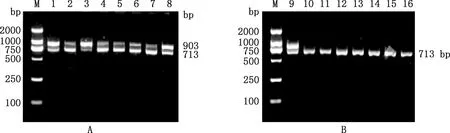
①A,无角山羊检测结果;B,有角和间性山羊检测结果。②M,DL2000 DNA Marker;1~8,无角山羊;9,有角山羊;10~16,间性山羊①A,Test results of hornless goats;B,Horned and intersex goats test results.②M,DL2000 DNA Marker;1-8,Hornless goats;9,Horned goats;10-16,Intersex goats图3 引物PIS wt-1的PCR检测结果Fig.3 PCR detection results of primer PIS wt-1
2.4 PIS区10.1 kb缺失/480 kb重复的基因型鉴定
PCR扩增结果显示,有角山羊只有214 bp片段(图5A、5B),其基因型为野生型纯合子,并没有发生10.1 kb缺失/480 kb重复,说明有角山羊并不会出现复杂的结构变异;而间性山羊不仅检测出了下游断点的680 bp片段(图5A),还检测出了上游断点的271 bp片段(图5B),其基因型为缺失突变纯合子,同时发生了10.1 kb的双缺失和480 kb的双重复。在检测中发现10只基因型为双缺失的纯合子无角公羊(表3),其余无角山羊同时具有214 bp片段以及下游断点的680 bp片段和上游断点的271 bp片段(图5A、5B),其基因型为突变杂合子,突变基因包含10.1 kb缺失/480 kb重复。

M,DL2000 DNA Marker;1~3,有角山羊;4~6,无角山羊;7~9,间性山羊。下同M,DL2000 DNA Marker;1-3,Horned goats;4-6,Hornless goats;7-9,Intersex goats. The same as below图4 引物PIS wt-3的PCR验证结果Fig.4 PCR validation results of primer PIS wt-3
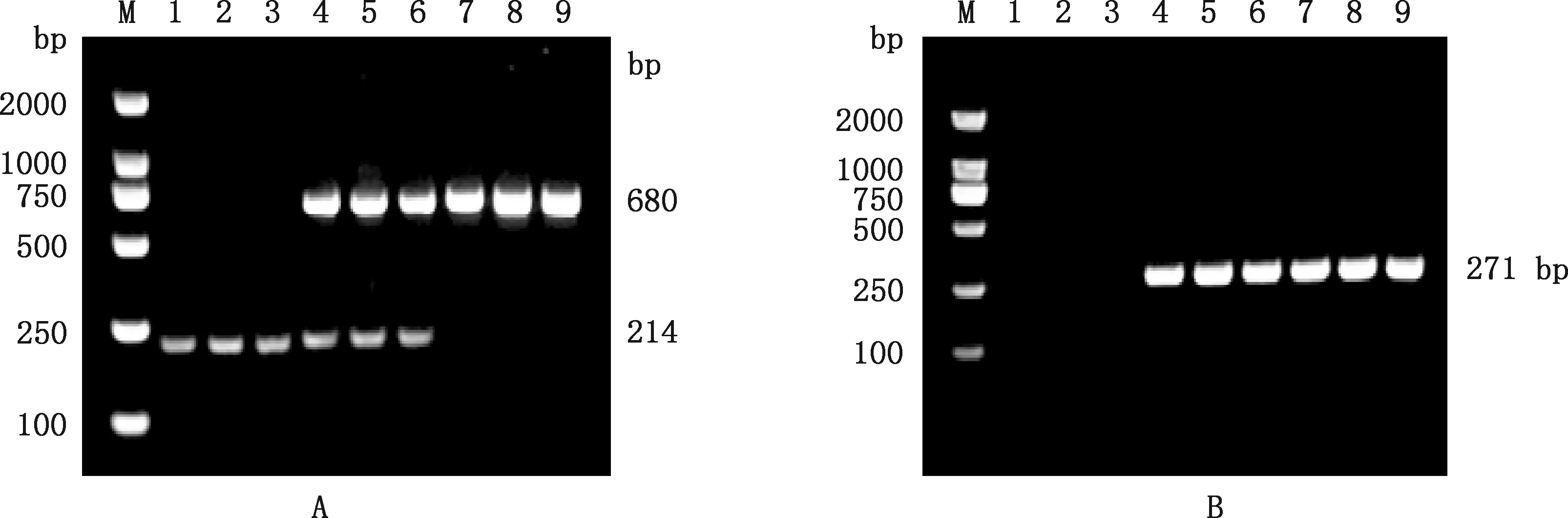
A,引物PIS wt-2和PIS var-2的PCR结果;B,引物PIS var-1的PCR结果A,The PCR results of primers PIS wt-2 and PIS var-2;B,The PCR results of primer PIS var-1图5 崂山奶山羊的2个结构变异PCR检测结果Fig.5 PCR detection results of two structural variations in Laoshan dairy goats

表3 崂山奶山羊群体PIS生殖缺陷基因型检测结果
3 讨 论
在山羊中,无角性状是显性性状,而与其紧密连锁的间性性状则是隐性性状,间性山羊表现出一定的发育障碍和卵巢畸形,无法应用于育种。尽管在20年前Pailhoux等[9]就已经明确了造成山羊雌性性别反转这一现象的原因,但是在此后的研究中却发现间性山羊的出现并不是简单的11.7 kb的纯合缺失[10-11,13]。而最近的一项研究中使用长读测序技术精准地将PIS缺失区的长度从11.7 kb缩短至10 159 bp,而且原位于1号染色体150 334 286~150 818 098 bp的约480 kb的重复序列反向插入到PIS缺失区的位置中[12],这种变异在中国唐山奶山羊和金堂黑山羊中也得到了印证[14-15]。本研究结果表明间性崂山奶山羊的PIS区是完全缺失的,且符合10.1 kb缺失/480 kb重复插入这一变异特征。另外,在1号染色体129 427 003~129 427 905 bp区域并不存在198 bp的替换和108 bp的缺失,这与198 bp替换和108 bp缺失也可能引发山羊间性性状的结论并不相同[10]。对155只崂山奶山羊进行基因分型时发现所有有角山羊等位基因都是纯合的,没有缺失和重复;而间性山羊等位基因的缺失和重复都是纯合的,这与先前大部分报道的结果一致[12,14-15],但是与个别研究结果不同[10-11]。而在检测的无角公羊中发现大多数基因型都是杂合的,只有10只等位基因为纯合型的双缺失无角公羊,与预期结果一致。以上结果证实了突变杂合子无角母羊是可育的,而间性性状仅发生在突变纯合子母羊中[16-17]。
目前,推测山羊1号染色体129 Mb处的10.1 kb缺失/480 kb重复至少影响FOXL2、PISRT1、钾离子通道15(potassium inwardly rectifying clannel subfamily J member 15,KCNJ15)和ETS转录因子(ETS transcription factor,ERG)基因的表达,进而影响了山羊的性别分化过程。FOXL2基因是哺乳动物的雌性性别决定基因[18-19],具有阻止睾丸形成的功能[20],其缺失可上调SRY转录因子9(SRY-box transcription factor 9,SOX9)的表达[21];研究证明,PIS区的缺失和480 kb的重复不仅影响FOXL2基因的转录[9],还可导致FOXL2基因下游出现几个特定的环状结构[14];在对来自山羊、绵羊和鹿的221个转录组的分析中发现FOXL2基因是一种与角的发育有关的基因[22]。PISRT1基因是一种睾丸抑制因子,可抑制SOX9基因的表达[23];在间性山羊的性腺中PISRT1基因表达下降、SOX9基因表达增加[9],且其基因多态性与间性山羊存在紧密联系[24]。因此,可将PISRT1基因作为调控山羊间性性状的候选基因。480 kb重复片段包含KCNJ15和ERG基因,而这2种基因的额外拷贝可能在角和性腺发育中起重要作用。KCNJ15基因在卵泡上皮细胞中高表达[25],说明KCNJ15基因可能参与卵巢功能的发育。ERG基因不仅是一种致癌基因[26],还参与生物体的骨骼发育[27]。以上这些基因可能在角的发育和生长代谢方面具有潜在的调节作用,并且与间性山羊的出现密切相关。
牛的无角性状受多个复等位基因的控制且不同品种个体间存在差异已被证实[28-31]。绵羊的角性状至少受2个基因位点的调控也已经被证实[3-4,7-8],但是山羊的无角性状和间性性状的分子遗传学基础仍不清楚。尽管对间性山羊和正常山羊的PIS变异区域进行三维结构比较发现了显著的染色体结构差异,但是依然没有强有力的证明PIS变异区域的这些序列是如何相互影响的[14]。FOXL2基因功能的丧失导致雌性山羊胎儿出现性反转[19],而在间性山羊性腺中转基因表达PISRT1基因并不能逆转山羊的间性性状[32];另外,在480 kb处发现14个与山羊角型性状完全相关的SNPs位点[15]。据此推断,山羊的无角显性基因可能位于480 kb上,而间性山羊的出现是由于10.1 kb的缺失影响了FOXL2基因的转录造成的,二者紧密连锁,其双突变导致山羊出现间性性状且仅发生在雌性山羊中,杂合突变仅表现为无角。虽然目前的研究依然没有解释清楚无角性状和间性性状的作用机制,但是明确了在间性山羊中10.1 kb的缺失是完全的,并且在崂山奶山羊中10.1 kb缺失/480 kb重复双突变结合遗传学性别鉴定可以作为诊断间性山羊的依据。
4 结 论
间性山羊的出现是由于1号染色体下游129 Mb区域总长度约10.1 kb的完全缺失和1个反向插入的约480 kb大小的重复片段导致,且间性性状仅发生在纯合子母羊中,而公羊的纯合缺失并不会出现间性性状。本研究结果为进一步揭示山羊无角性状的遗传机制以及培育崂山奶山羊无角新品系提供重要参考。
