金黄色葡萄球菌处理对奶牛乳腺上皮细胞外泌体表征的影响
2022-08-23朱晓艳王梦玲刘子豪熊本海
蔡 萌,朱晓艳,王梦玲,刘子豪,熊本海,杨 亮
(中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193)
乳房炎是奶牛养殖中最重要的疾病之一,给全球牧场带来巨大的经济损失[1]。乳房炎造成奶牛产奶量显著下降、乳品质量下降、兽医护理成本增加及过早淘汰的损失约占总经济损失的70%[2]。乳房炎通常由微生物感染引起,按乳房症状及乳汁变化分为临床型和亚临床型乳房炎,临床型乳房炎的主要症状表现为乳房患处出现不同程度的充血、肿胀、手感温热伴有疼痛,乳汁变黄、质地变稠、偶有乳凝块等[3]。亚临床型乳房炎一般没有明显的临床症状,肉眼无法看出异常,但是产奶量出现下降且乳中病原菌、脓球、白细胞含量增加而乳脂、蛋白质、乳糖、钾、钙、镁、磷、铁和锌含量下降[4]。相对于临床型乳房炎,亚临床型乳房炎更难治疗,潜在危害更大。奶牛乳腺上皮细胞(bovine mammary epithelial cell,BMEC)是乳腺感染时的第一道防线,是抵御病原微生物入侵的天然屏障,可以启动机体对病原微生物最早的免疫识别和免疫应答,并协调后续免疫分子、免疫细胞应答等[5]。金黄色葡萄球菌(以下简称金葡菌)、无乳链球菌、乳房链球菌、大肠杆菌等是乳房炎的主要致病菌。其中,金葡菌是最常见且最重要的病原菌之一[6]。由于金葡菌的抵抗力及其逃避固有免疫和获得性免疫反应的能力,经常引起亚临床乳房炎和慢性乳房炎[7]。金葡菌可以黏附侵入BMEC诱导细胞内自噬体的形成,阻止胞内细菌清除,从而导致持续感染并引起慢性疾病[8-10]。
外泌体是一种直径大小在30~150 nm之间的纳米级小囊泡,它可以特异性地包裹一些蛋白质、脂质、核酸或miRNA等物质。外泌体具有生物活性,能够被受体细胞吸收,实现细胞间的物质运输和信息传递[11]。不同细胞来源的外泌体在免疫调节中起着不同的作用[12]。外泌体的来源非常广泛,几乎所有类型的细胞均可分泌,如凋亡细胞、巨噬细胞、淋巴细胞、树突状细胞以及星形胶质细胞等[13-17]。外泌体普遍存在于各种体液中,如血清、血浆、尿液和母乳等[18-20]。研究发现,母乳中富含外泌体,但其来源不能确定,可能来源于BMEC、巨噬细胞、淋巴细胞,甚至来源于身体其他可以通过血液循环到达母乳的细胞[21-22]。外泌体与许多生理病理学功能有关,如信号传导、免疫和感染等[17]。外泌体可以直接将病原体相关分子从细胞转移到细胞,从而影响感染进程。结核分枝杆菌感染的巨噬细胞释放的外泌体,可将分枝杆菌蛋白、脂质和核酸等传递给幼稚巨噬细胞,从而激活或抑制免疫反应[17]。在黏附侵袭性大肠杆菌感染肠上皮细胞后,细胞分泌的外泌体可以将特定的miRNA(如miR-30c和miR-130a)转移到受体细胞,从而抑制自噬介导的细胞内细菌清除过程[23-24]。上述研究证实感染细胞释放的外泌体参与宿主对病原感染的免疫反应。牛奶衍生的外泌体含有大量与乳腺和免疫相关的miRNA。试验性或临床性金葡菌感染导致的奶牛乳房炎会引起牛奶中外泌体miRNA表达谱的差异,差异miRNA包括miR-142-5p、miR-223、miR-223、miR-378和miR-185[25-27]。然而,BMEC感染乳房炎病原菌金葡菌与细胞释放外泌体之间的关系仍然未知。本研究将探究不同金葡菌与BMEC数量比值(MOI)和处理后无外泌体培养时间对外泌体总浓度的影响,并对金葡菌诱导BMEC释放的外泌体进行分离和鉴定,旨在确立一套金葡菌诱导BMEC释放外泌体的细胞模型,为进一步探究其在感染进程中的作用奠定基础。
1 材料与方法
1.1 材料
奶牛乳腺上皮细胞(MAC-T细胞系)及金葡菌均由中国农业科学院北京畜牧兽医研究所智慧畜牧业团队保存。
1.2 主要试剂及仪器
LB肉汤、琼脂粉均购自博奥星生物公司;CHROMagar培养基购自科玛嘉公司;胎牛血清、胰蛋白酶、双抗(青霉素、链霉素)均购自Gibco公司;DMEM/F12培养基、PBS均购自HyClone公司;ECL发光液购自Tanon公司;庆大霉素、SDS-PAGE凝胶制备试剂盒均购自索莱宝科技有限公司。BCA试剂盒购自Thermo Fisher Scientific公司;兔抗CD9、CD81、TSG101单克隆抗体均购自Abcam公司。
主要仪器有超速离心机(Optima XE-90,贝克曼库尔特公司);透射电子显微镜(HT7700,日立公司);扫描电子显微镜(LSM 900,蔡司公司)。
1.3 无外泌体胎牛血清的制备
将胎牛血清于4 ℃、110 000×g离心18 h,去除外泌体。缓慢收集上层血清,避免扰动底部的沉淀,收集的上层血清即为无外泌体血清,在细胞间无菌超净台中使用0.22 μm滤器过滤后分装,-20 ℃保存备用。
1.4 建立金葡菌处理BMEC模型
1.4.1 准备菌株 使用LB肉汤复苏金葡萄菌株,使用CHROMagar培养基纯化菌株,取直径1 mm单菌落增菌4 h,4 ℃保存待用。
1.4.2 准备细胞 细胞接种至T75细胞瓶中,每组15瓶,待细胞汇合度达到85%后做攻菌处理,此时每瓶约含2.3×106个细胞。
1.4.3 金葡菌处理BMEC 攻菌前更换细胞培养基为感染培养基(含5%无外泌体血清的DMEM/F12培养基,无双抗)。MOI=1组加入金葡菌2.3×106/瓶;MOI=10组加入金葡菌2.3×107/瓶,于37 ℃恒温培养箱中培养3 h。同时以未处理细胞作为空白对照组。
1.4.4 收集细胞上清 攻菌结束后彻底弃掉感染培养基,PBS清洗多次,以保证清洗掉未黏附的细菌。更换无外泌体培养基(含10%无外泌体血清、含1%双抗、200 μg/mL庆大霉素)。将攻菌后的细胞分别培养9、12、24 h,收集细胞上清并用0.22 μm滤器过滤。
1.5 金葡菌处理BMEC后细胞形态学观察
将细胞爬片提前置于6孔细胞培养板底部,然后进行细胞接种培养。PBS清洗细胞并压碎细胞爬片。将细胞爬片碎片放入到4 ℃预冷的3%戊二醛中,4 ℃过夜固定。弃掉固定剂后,用PBS清洗细胞爬片碎片2次,每次10 min,加入预冷的1%锇酸4 ℃固定1 h,再次清洗细胞爬片碎片。脱水,干燥。使用扫描电子显微镜观察并拍照。
1.6 外泌体的分离纯化及鉴定
1.6.1 外泌体分离纯化 将收集的上清于4 ℃、110 000×g离心90 min,缓慢不彻底弃上清,避免扰动底部的沉淀,使用PBS重悬沉淀、再次离心,用PBS清洗外泌体。向外泌体沉淀中加入200 μL PBS重悬,并转移到1.5 mL离心管中,彻底吹匀后-80 ℃保存备用。
1.6.2 外泌体形态观察 取10 μL外泌体悬液滴在封口膜上,将铜网膜面置于外泌体悬滴上保持3 min,再置于蒸馏水悬滴上,保持1 min,然后置于醋酸双氧铀悬滴上,保持5 min。膜面朝上标记好样品名称置于平皿中,室温干燥,保持洁净。待铜网干燥后,在透射电子显微镜下观察外泌体形态并拍照。
1.6.3 外泌体粒径分析 取外泌体20 μL,PBS稀释5 000倍,通过粒径分析仪(Zeta View,Particle Metrix)实时对悬浮液中50~1 000 nm直径范围内特定的外泌体等小囊泡进行逐个直接成像和观察,最后通过软件Zeta view 8.04.02计算外泌体粒径分布。
1.6.4 外泌体标记蛋白CD9、CD81、TSG101的鉴定 每组取外泌体10 μL,使用RIPA裂解液提取总蛋白;总蛋白通过15% SDS-PAGE分离后转移至PVDF膜,将PVDF膜置于5% BSA中37 ℃封闭1 h;1×PBST缓冲液洗涤2次,每次10 min。加入兔抗CD9(1∶500稀释)、CD81(1∶500稀释)、TSG101(1∶500稀释),4 ℃过夜。1×PBST缓冲液洗涤2次,每次10 min,加入HRP标记的抗兔IgG(H+L)(1∶5 000稀释),37 ℃反应50 min;1×PBST缓冲液洗涤2次,每次10 min。ECL显影,根据蛋白大小调节曝光时间,保存图像备用。
1.7 数据统计分析
采用SPSS 20.0软件进行非参数分析(Kruskal-Wallis),考察不同MOI及处理后培养不同时间对外泌体总蛋白浓度的影响。结果以平均值±标准误表示,用GraphPad Prism 6.0软件作图。P<0.05表示差异显著。
2 结 果
2.1 金葡菌处理BMEC后细胞形态学分析
通过扫描电镜观察细胞表面超微结构(图1)。由图1A可知,空白对照组BMEC形态饱满、结构完整,呈现椭圆形,表面微绒毛排列有序。金葡菌呈典型圆球状,处理BMEC 3 h后,大量黏附在细胞膜表面;MOI=1和MOI=10组BMEC表面微绒毛均消失,细胞骨架萎缩、凹陷,后者细胞受损程度更严重(图1B和图1C)。
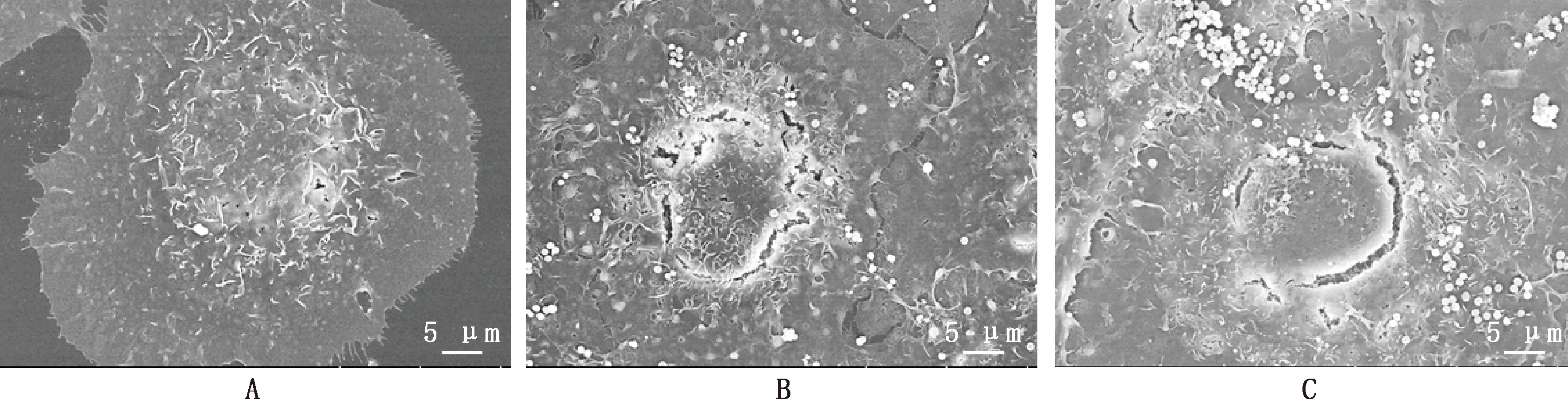
A,空白对照组;B,MOI=1;C,MOI=10。图2同A,Control group;B,MOI=1;C,MOI=10. The same as fig.2图1 不同处理组BMEC表面超微结构观察(2 000×)Fig.1 Ultrastructural observation of BMEC surface in different processing group (2 000×)
2.2 外泌体形态学分析
采用透射电镜观察BMEC外泌体的形态(图2)。由图2可知,各组外泌体均为大小类似、形态一致的小囊泡,直径在100~150 nm之间,均可见明显双层膜、中间凹陷的茶托样结构,各组形态无明显差异。其中,MOI=10组(图2C)外泌体密度大于MOI=1组(图2B),且两组均大于空白对照组(图2A)。
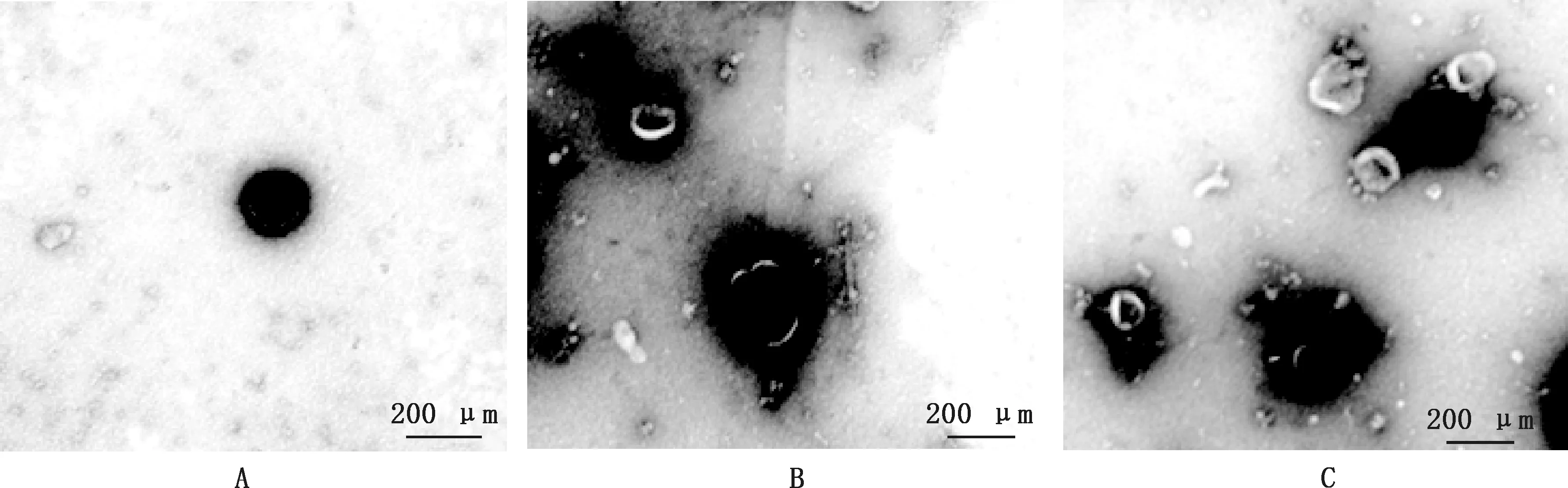
图2 不同处理组BMEC释放的外泌体形态观察(50 000×)Fig.2 Morphological observation of exosomes released by BMEC in different processing group (50 000×)
2.3 外泌体总蛋白浓度分析
由图3可知,在各培养时间点,MOI=10时细胞上清外泌体总蛋白浓度高于其他2组,但各组间差异不显著(P>0.05)。感染后继续培养12 h,空白对照组及MOI=1、MOI=10攻菌组细胞上清外泌体总蛋白浓度均值分别为499.65、611.75和781.68 μg/mL,其中以MOI=10组获取的外泌体总蛋白浓度最高。
根据外泌体总蛋白浓度结果,确定攻菌量MOI=10和攻菌后培养时间为12 h。后续外泌体鉴定只保留空白对照组和MOI=10组。
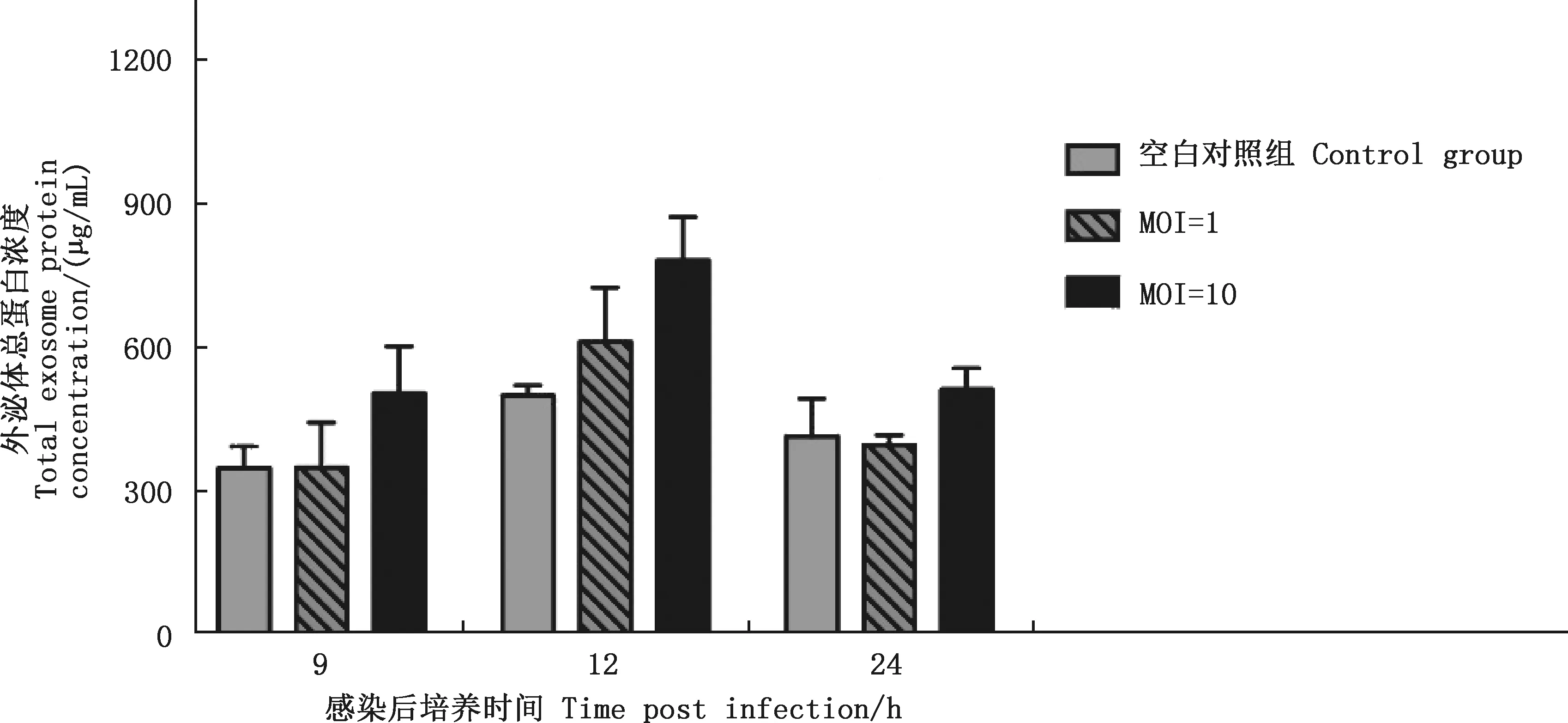
图3 各处理组不同培养时间BMEC上清中外泌体总蛋白浓度Fig.3 Concentration of exosome protein from BMEC supernatant in each treatment group at different cultured time
2.4 外泌体粒径分析和CD9、CD81、TSG101蛋白鉴定
使用纳米颗粒追踪分析技术确定外泌体大小,外泌体大小均一,无明显差异(图4)。其中,空白对照组平均粒径112 nm,粒径在40~185 nm 范围的颗粒数占总颗粒数的96.62%(图4A);MOI=10组平均粒径116 nm,粒径在40~185 nm 范围的颗粒数占总颗粒数的97.73%(图4B)。Western blotting检测结果显示,空白对照组和MOI=10组外泌体标记蛋白CD9、CD81、TSG101表达均呈阳性(图4C)。

A、B,分别为空白对照组和MOI=10组外泌体粒径分析;C,Western blotting检测结果A and B,The detection results of control group,MOI=10;C,Western blotting detection rusults图4 外泌体纳米颗粒追踪分析和标记蛋白表达Fig.4 Nanoparticle tracking analysis and labeled protein expression of exosomes
3 讨 论
乳房炎几乎总是由病原微生物引起的一种奶牛乳房炎症。它严重损害了奶牛养殖和乳制品行业。当BMEC细胞等受到细菌毒素的刺激后,会分泌大量的细胞因子和炎症介质,进而引起乳房炎。乳房炎不仅造成产奶量下降,还降低了乳品质,甚至危害人体健康,严重影响奶牛养殖业的经济效益。乳腺是奶牛防御疾病的第一道防线,BMEC不仅是抵御外来病原微生物入侵的天然屏障,还可以启动机体对病原微生物最早的免疫识别和免疫应答[5]。外泌体通过旁分泌途径在细胞之间进行交流,其携带的物质是启动通信的开关[28]。本研究发现,金葡菌以MOI=10处理BMEC 3 h后,继续无外泌体培养细胞12 h收集上清,利用超速离心法快速获得大量的外泌体。透射电镜观察结果显示,囊泡颗粒具有外泌体典型双层膜中间凹陷的茶托样结构,各组样品整体均一性良好。CD9、CD81和TSG101 蛋白为典型的外泌体标记蛋白,其主要涉及到了外泌体的生物发生过程。CD9、CD81是参与外泌体运输的四跨膜蛋白家族成员,TSG101是转运必需内体分选(ESCRT)复合体相关的蛋白,而ESCRT复合体是膜形成和断裂的驱动器[29]。Western blotting结果显示,本研究分离的囊泡颗粒表达外泌体表面标志物CD9、CD81和TSG101,进一步证明了透射电镜观察到的囊泡状物质为外泌体。粒径分析结果显示,细胞上清中的空白对照组外泌体平均粒径约112 nm,MOI=10组外泌体平均粒径约116 nm,两组外泌体粒径范围在40~185 nm占比高达95%以上,符合外泌体定义大小。
相同培养条件下,与MOI=1相比,MOI=10能刺激BMEC释放更高浓度的外泌体。如克罗恩病患者的回肠黏膜中存在高流行的黏附侵袭性大肠杆菌。从表型上观察,大肠杆菌可以通过与受体细胞的结合来增加肠上皮的通透性,进而促进其在巨噬细胞中存活和增殖。大肠杆菌在巨噬细胞内可以诱导细胞分泌更高浓度的外泌体[30]。本研究发现,在相同MOI时,继续无外泌体培养12 h,可收集分离的外泌体总量最多。推测原因是金葡菌侵染细胞后定植在胞内,培养24 h比12 h会导致更多的细胞死亡、凋亡,且破碎的细胞会释放酶类物质,胞内细菌也会逃出细胞,这些都有可能会破坏上清中外泌体囊膜的完整性,进而影响外泌体的总量。已有研究报道,脂多糖可以刺激奶牛BMEC产生外泌体,引起外泌体中的miRNA差异变化[31-32]。转化生长因子-β1(TGF-β1)刺激牛BMEC产生外泌体通过激活p38-MAPK通路抑制牛巨噬细胞的增殖,并可能干扰奶牛的免疫力[33]。本研究检测不同MOI处理和培养时间的外泌体总量蛋白浓度,结果无显著差异,这可能与奶牛乳腺的天然免疫机制有关。据报道,革兰氏阳性细菌可引起奶牛乳腺较为缓慢或中等程度的免疫反应[34],与金葡菌有关的乳房炎引起的先天免疫反应非常缓慢甚至不可察觉[35]。这种情况下外泌体总量无显著差异,但外泌体功能可能存在一定的差异。自噬是一种参与清除入侵的病原微生物的生物过程,自噬体吞噬病原体并与溶酶体融合以发挥降解作用[36]。然而,一些细菌通过干扰自噬来逃避自噬降解机制,从而为自身的生存创造了有利条件。黏附侵袭性大肠杆菌感染肠上皮细胞后,细胞分泌的外泌体可以将特定的miRNA(如miR-30c和miR-130a)转移到受体细胞,从而抑制自噬介导的细胞内细菌清除过程[30,37]。外泌体也与携带成孔α-毒素的金葡菌的致病性密切相关[38]。上述研究证实细菌感染会促使细胞释放外泌体,但细菌侵入BMEC时自噬是否参与了细菌感染机制有待进一步研究。今后将进一步研究细菌感染对细胞释放的外泌体携带物质组成及功能的影响。
4 结 论
本研究从形态学和分子生物学特征等方面证实试验获得的分离物为外泌体;MOI=10时,金葡菌处理细胞3 h后继续无外泌体培养12 h,收获的细胞上清中外泌体蛋白含量最高。本研究可为金葡菌诱导BMEC释放外泌体的后续研究提供参考。
