山羊干扰素刺激基因15克隆、生物信息学分析及亚细胞定位
2022-08-23唐井玉杜汉宇孟春春刘光清
唐井玉,杜汉宇,2,孟春春,刘光清
(1.中国农业科学院上海兽医研究所,上海 200241;2.新疆农业大学,乌鲁木齐 830052)
干扰素刺激基因15 (interferon-stimulated gene 15,ISG15) 是干扰素诱导最强烈最快的ISG分子之一。所有物种的ISG15具有相似的结构,均由2个泛素样蛋白 (Ubiquitin-like protein,Ubl) 结构域组成并都含有特征性的β-折叠,这2个结构域被称为“铰链”的多肽序列连接[1]。但与其他泛素蛋白相比,ISG15序列差异大,具有多样性的特征。ISG15有2种发挥作用的形式,一种是通过ISG化修饰与蛋白质中的赖氨酸残基结合发挥作用,整个过程与泛素化过程相似,包括了E1激活酶、E2结合酶和E3连接酶,USP18解聚酶可以将ISG15从被修饰的蛋白质上移除[2],目前鉴定出的可以被ISG化修饰的蛋白已有上百种[3];另一种作用形式是未结合的处于游离状态的ISG15。
ISG15具有多种生物学功能,包括调节宿主对病原微生物感染的应答[4-5]、调节宿主信号通路[6-7]、发挥免疫调节作用[8],同时还参与调节宿主损伤和DNA修复[9-10]、肿瘤的发生[11]、凋亡和自噬[12],在生命活动过程中发挥重要的作用。目前关于羊ISG15的研究较少,主要研究集中在ISG15对反刍动物妊娠期的调节,ISG15被认为是决定子宫对胚胎接受性的重要因素之一。牛、水牛和绵羊在妊娠早期ISG15会短暂表达[13]。羊ISG15在抑制病毒感染、调节山羊乳腺上皮细胞凋亡和炎症等方面有了初步研究[14-15]。
ISG15从发现至今已有40多年,但由于不同宿主物种之间ISG15功能和结构之间存在差异,因此研究进展缓慢。目前关于人和小鼠ISG15的作用、空间结构等研究较多,但对于山羊的研究报道较少。本研究克隆表达山羊ISG15并通过生物信息学分析了解其基本的信息、结构和功能,对其亚细胞定位初步研究,旨在进一步理解山羊ISG15蛋白质的功能,为后续研究山羊ISG15的功能奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株、载体及细胞 大肠杆菌DH5α感受态细胞购自南京诺唯赞生物科技股份有限公司;真核表达质粒pCMV-Myc载体、HEK-293T细胞和山羊子宫内膜上皮细胞(EEC)均由上海兽医研究所伴侣动物生物安全与防控技术团队保存。
1.1.2 主要试剂与仪器 反转录试剂、胶回收试剂盒和无缝连接试剂盒均购自南京诺唯赞生物科技股份有限公司;Trizol、Lipofectamine 3000脂质体转染试剂盒、蛋白Marker、ECL显色液、羊抗兔荧光二抗594和羊抗鼠荧光二抗594均购自Thermo公司;无内毒素质粒小提中量试剂盒购自天根生化科技(北京)有限公司;EcoRⅠ和XhoⅠ限制性内切酶均购自NEB公司;DMEM-F12、DMEM和Opti-MEM培养基均购自Gibco公司;鼠源Myc单克隆抗体、牛血清白蛋白均购自西格玛奥德里奇贸易有限公司;兔源ISG15多克隆抗体由上海兽医研究所伴侣动物生物安全与防控技术团队制备并保存;蓝色荧光染料DAPI购自上海碧云天生物技术有限公司。倒置荧光显微镜购自Nikon公司。
1.2 方法
1.2.1 引物设计及合成 根据GenBank数据库中预测的山羊ISG15基因序列(登录号:XM_005690795),利用SnapGene软件设计扩增山羊ISG5基因的特异性引物,上游引物ISG15-F:5′-tggaggcccgaattcGGATGGGCGGGGACCTGAAG-3′,下游引物ISG15-R:5′-ggccgcggtacctcgagCTACCC-ACCCCGCAGACG-3′,上、下游引物分别加入pCMV-Myc载体中EcoRⅠ和XhoⅠ两个酶切位点两端的同源臂(小写字母部分),预期扩增片段大小为474 bp。引物由北京擎科生物科技有限公司合成。
1.2.2 山羊ISG15基因PCR扩增 将EEC细胞样品按照Trizol说明书提取总RNA。根据反转录试剂盒说明进行反转录得到的cDNA。以cDNA为模板,用高保真DNA聚合酶进行PCR扩增,PCR反应体系50 μL:2×Phanta Max MasterMix 25 μL,上、下游引物各2 μL,模板cDNA 2 μL,ddH2O补足至50 μL。PCR反应条件为:95 ℃预变性3 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s,共35个循环;72 ℃终延伸10 min。PCR产物经2.0%琼脂糖凝胶电泳检测。
1.2.3 pCMV-Myc-ISG15重组真核表达载体的构建及表达 将pCMV-Myc质粒用EcoRⅠ和XhoⅠ两个限制性内切酶37 ℃酶切2 h,使其线性化,酶切产物经1.0%琼脂糖凝胶电泳检测。将PCR产物与酶切后的载体进行胶回收,测定浓度。将回收后的酶切产物和线性化的pCMV-Myc载体通过ClonExpress Ⅱ One Step Cloning Kit (C112) 进行同源重组。插入片段与线性化载体摩尔比为2∶1,同源重组反应体系为:线性化载体1 μL,目的片段3 μL,5×CE Ⅱ Buffer 2 μL,ExnaseⅡ 1 μL,ddH2O 3 μL。PCR仪中37 ℃连接30 min,然后将连接产物转化大肠杆菌DH5α感受态细胞,挑选菌落,PCR鉴定正确后送至北京擎科生物科技有限公司测序。测序正确后根据无内毒素质粒小提中量试剂盒说明书提取质粒并测定浓度。将构建完成的pCMV-Myc-ISG15质粒按照Lipofectamine 3000脂质体转染说明转染至HEK-293T细胞中,转染pCMV-Myc空载为对照组。转染24 h收取细胞样品进行Western blotting检测pCMV-Myc-ISG15质粒的表达。
1.2.4 山羊ISG15基因的生物信息学分析 从NCBI-Uniport(https:∥www.uniprot.org)和GenBank两个数据库中获得其他19个物种的ISG15序列(表1),利用MegAlign和Mega 6.0软件对ISG15基因核苷酸和氨基酸序列相似性比对及系统进化树进行分析;同时利用在线软件ProtParam (https:∥web.expasy.org/protparam/)对山羊ISG15理化性质进行分析;利用TMHMN Server v.2.0 (https:∥services.healthtech.dtu.dk/service.php?TMHMM-2.0) 和SignalP 5.0 (https:∥services.healthtech.dtu.dk/service.php?SignalP-5.0)、NetNGlyc-1.0 (https:∥services.healthtech.dtu.dk/service.php?NetNGlyc-1.0)、YinOYang (https:∥services.healthtech.dtu.dk/service.php?YinO Yang-1.2)和NetPhos-3.1程序(https:∥services.healthtech.dtu.dk/service.php?NetPhos-3.1)预测山羊ISG15蛋白的跨膜结构、信号肽、N-糖基化、O-糖基化位点和磷酸化位点;利用SPOMA(https:∥npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html) 和SWISS-MODEL软件(https:∥swissmodel.expasy.org)预测山羊ISG15蛋白的二级结构和三级结构;利用STRING软件(https:∥cn.string-db.org)预测与山羊ISG15相互作用的蛋白;通过PredictProtein (http:∥predictprotein.org) 预测山羊ISG15蛋白的生物学功能;应用在线软件PSORT Ⅱ Prediction (http:∥psort.hgc.jp/form2.html) 预测山羊ISG15蛋白在细胞中的定位。
1.2.5 山羊ISG15蛋白在EEC细胞中的亚细胞定位 将pCMV-Myc-ISG15质粒转染至EEC细胞,36 h后进行间接免疫荧光试验(indirect immunofluorescence assay,IFA),检测外源性ISG15的亚细胞定位。同时用小反刍兽疫病毒(PPRV)感染EEC细胞,36 h后进行IFA,检测内源性ISG15的亚细胞定位。
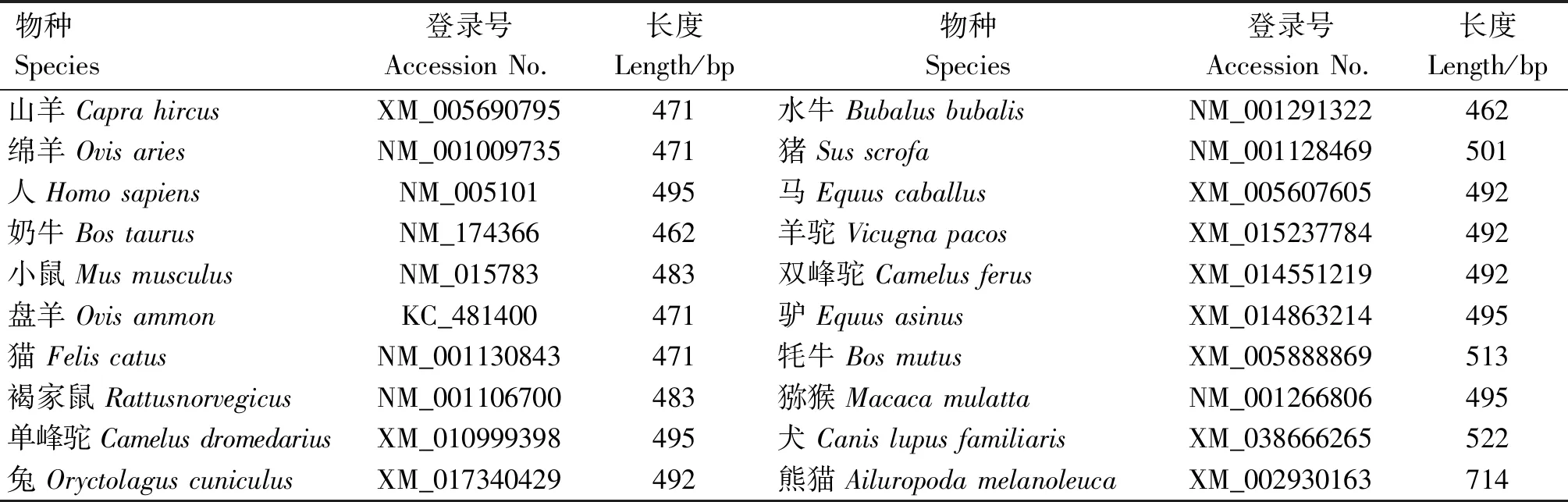
表1 GenBank中不同物种ISG15基因序列信息
2 结 果
2.1 山羊ISG15基因的克隆、真核表达载体的构建和表达
以EEC细胞cDNA为模板扩增山羊ISG15基因,获得了474 bp的目的条带(图1A),与预期结果相符。回收PCR产物连接至pCMV-Myc酶切后的空载体(图1B),转化大肠杆菌DH5α感受态细胞进行阳性克隆筛选,并将PCR鉴定正确的pCMV-Myc-ISG15质粒送测序后,对比序列发现克隆的山羊ISG15与NCBI数据库中预测的序列一致,且pCMV-Myc-ISG15质粒构建成功。将pCMV-Myc-ISG15质粒转染至HEK-293T细胞,24 h后收取细胞样品,经Western blotting鉴定发现构建的pCMV-Myc-ISG15质粒成功表达,大小约为17 ku(图1C)。

A,山羊ISG15基因CDS区PCR扩增产物:M1,DL2000 DNA Marker;1,ISG15基因扩增产物;2,阴性对照。B,pCMV-Myc载体EcoRⅠ和XhoⅠ双酶切图:M2,DL5000 DNA Marker;1-2,pCMV-Myc空载EcoRⅠ和XhoⅠ双酶切。C,山羊ISG15真核表达产物:M3,蛋白质分子质量标准;1,pCMV-Myc-ISG15质粒表达产物;2,pCMV-Myc质粒表达产物A,PCR amplification product of ISG15 gene CDS in goats:M1,DL2000 DNA Marker;1,ISG15 gene amplification product;2,Negative control.B,pCMV-Myc plasmid digested by EcoRⅠ和XhoⅠ:M2,DL5000 DNA Marker;1-2,pCMV-Myc empty EcoRⅠ and XhoⅠ double digest.C,Eukaryotic expression product of ISG15 in goats:M3,Protein Marker;1,Expression product of pCMV-Myc-ISG15 plasmid;2,Expression product of pCMV-Myc plasmid图1 山羊ISG15基因扩增及表达Fig.1 Amplification and expression of ISG15 gene in goats
2.2 相似性比对和系统进化树构建
2.2.1 相似性比对 核苷酸和氨基酸序列相似性分析结果发现,山羊ISG15与盘羊核苷酸和氨基酸序列相似性最高,分别为99.2%和100%,与绵羊的相似性次之,分别为98.9%和99.4%,与小鼠的核苷酸和氨基酸序列相似性最低,分别仅有72.6%和62.8%,与人的核苷酸和氨基酸序列相似性分别为75.8%和65.2%(表2)。MegAlign分析20个物种ISG15的氨基酸保守性,结果发现不同物种的ISG15序列长度不同,起始和终止位置也不相同,但末端均含有LRLRGG。与山羊ISG15相比,不同物种在不同位置氨基酸保守性有差异,在山羊ISG15的50-80位氨基酸区域突变较为集中。从整体上来看不同物种的ISG15序列具有多样性。

2.2.2 系统进化树构建 系统进化树分析结果表明,山羊与绵羊和盘羊属于一个分支,与奶牛、牦牛和水牛属于一个大分支;羊驼、单峰驼和双峰驼属于一个小分支,人和猕猴属于一个分支;大鼠和小鼠则属于另一个分支。从进化树图可以看出羊与牛类亲缘性较近,与灵长类和啮齿类亲缘性较远(图2)。
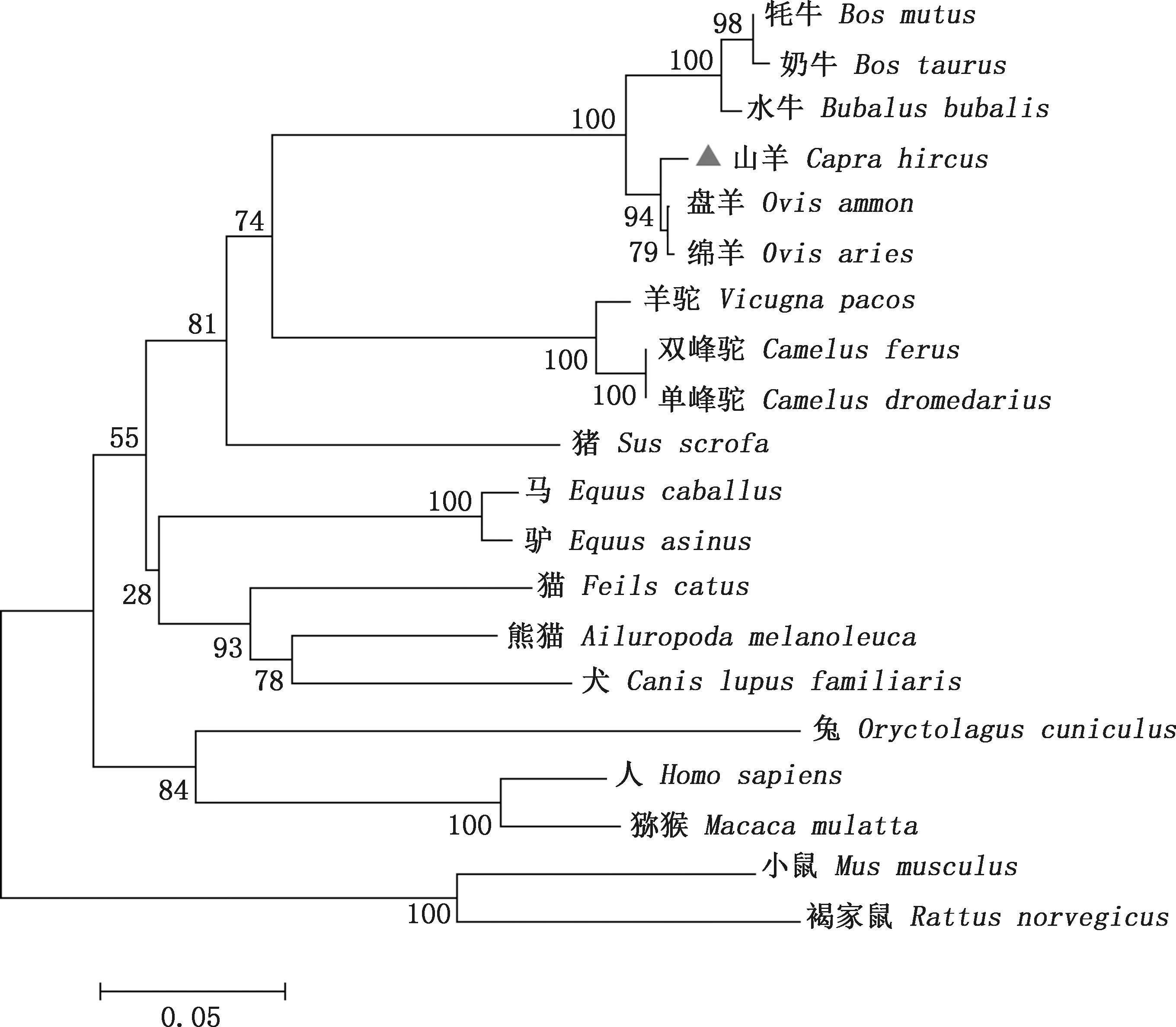
图2 ISG15基因系统进化树Fig.2 Phylogenetic tree of ISG15 gene
2.3 生物信息学分析
2.3.1 理化性质分析 根据NCBI数据库预测发现,山羊ISG15基因位于16号染色体上,CDS全长474 bp,编码区从第128位开始,止于601位,编码157个氨基酸。ProtParam分析发现,ISG15蛋白分子质量约为17.47 ku,等电点为6.83,分子式为C766H1252N216O299S10,由20种氨基酸组成,其中亮氨酸(Leu,L)有21个,含量最高,占13.4%,色氨酸(Trp,W) 含量最低,仅有1个,占0.6%;其中带正电荷残基(Asp+Glu)与带负电荷残基(Arg+Lys)数目相等,均为17(表3)。失稳指数(instability index,II) 为41.73,由于该值>40,因此该蛋白归为不稳定蛋白。脂肪族指数(aliphatic index)为96.75。亲水性平均值(grand average of hydropathicity,GRAVY) 为-0.148,该值<0,因此判断山羊ISG15蛋白为亲水性蛋白。
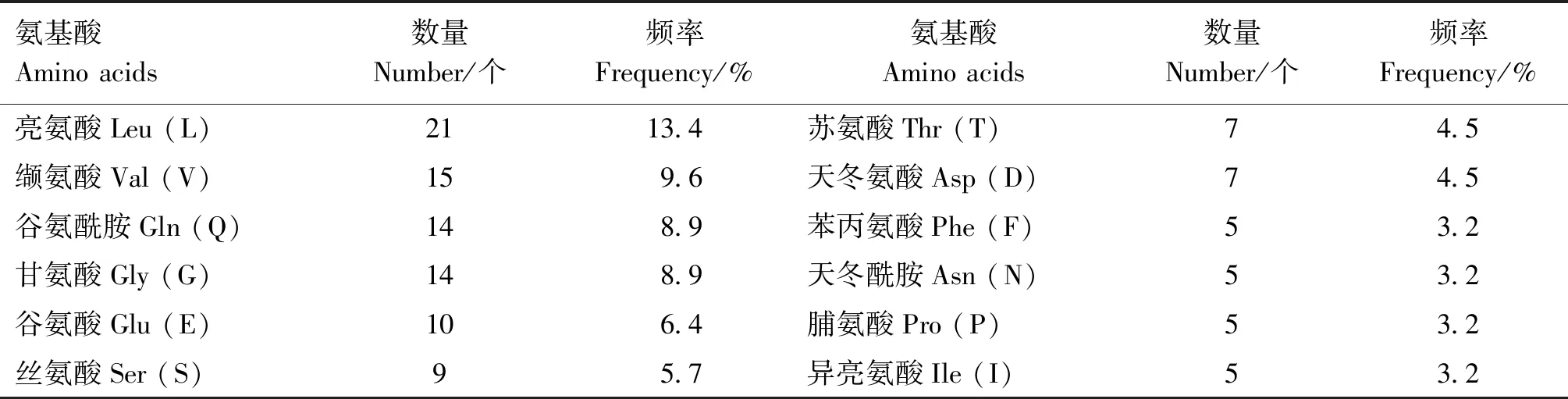
表3 山羊ISG15蛋白氨基酸组成

续表
2.3.2 跨膜结构及信号肽预测 在线软件TMHMN Server v.2.0和SignalP 5.0预测结果显示,山羊ISG15蛋白没有跨膜结构(图3A),也没有信号肽(图3B)。

图3 山羊ISG15蛋白跨膜结构(A)及信号肽(B)预测Fig.3 Prediction of transmembrane structure (A) and signal peptide (B) of ISG15 protein in goats
2.3.3 糖基化和磷酸化位点预测 预测结果显示,山羊ISG15蛋白存在1个潜在的N-糖基化位点,位于第77位氨基酸处(图4A);存在16个潜在的O-糖基化位点,分别位于第22、26、50、69、70、79、81、93、94、95、101、103、125、143、144和147位氨基酸处(图4B);存在10个潜在的磷酸化位点,其中包括6个丝氨酸位点(位于第22、69、93、94、95及105位氨基酸处)和4个苏氨酸位点(分别位于第101、103、144及147位氨基酸处)(图4C),结果显示该蛋白的磷酸化可能与蛋白激酶C (PKC)、肌酸激酶Ⅰ(CKⅠ)、DNA依赖性蛋白激酶 (DNAPK)、蛋白激酶A (PKA)、肌酸激酶Ⅱ(CKⅡ) 等有关。

图4 山羊ISG15蛋白糖基化(A、B)和磷酸化位点(C)预测Fig.4 Prediction glycosylation (A and B) and phosphorylation (C) sites of ISG15 protein in goats
2.3.4 山羊ISG15蛋白二级结构、三级结构及互作蛋白预测 利用在线软件SPOMA分析山羊ISG15的二级结构,发现该蛋白由4种结构组成,分别为无规则卷曲、延伸链、α-螺旋和β-转角,所占比例分别为34.39%、31.21%、21.66%和12.74%(图5)。根据SWISS-MODEL分析得到的山羊ISG15的三级结构与二级结构相似,也是由α-螺旋、β-转角、无规则卷曲和延伸链构成(图6)。与参考的绵羊ISG15模板相似性较高,达到了98.72%。利用STRING预测与山羊ISG15蛋白相互作用的蛋白,结果发现预测中的蛋白主要为干扰素和泛素化相关的蛋白。其中已经试验证明与其相互作用的蛋白有泛素激活酶 (Ubiquitin-activating enzyme,UBA7)、USP18和DDX解旋酶58 (DEDA-box RNA helicase,DDX58)。预测ISG15还与抗黏液病毒1 (Myxovirus resistance protein 1,MX1)、抗黏液病毒2(Myxovirus resistance protein 2,MX2)、寡腺苷酸合成酶1 (2′,5′-oligoadenylate synthetase 1,OAS1)、干扰素调节因子7 (interferon regulatory factor 7,IRF7)、干扰素诱导蛋白6 (interferon-inducible protein 6,IFI6)、干扰素诱导蛋白44 (interferon-inducible protein 44,IFI44)相互作用 (图7)。

h,α-螺旋;e,延伸链;c,无规则卷曲;t,β-转角h,Alpha helix;e,Extended chain;c,Random coil;t,Beta turn图5 山羊ISG15蛋白二级结构预测Fig.5 Secondary structure prediction of ISG15 protein in goats

图6 山羊ISG15蛋白三级结构预测Fig.6 Tertiary structure prediction of ISG15 protein in goats
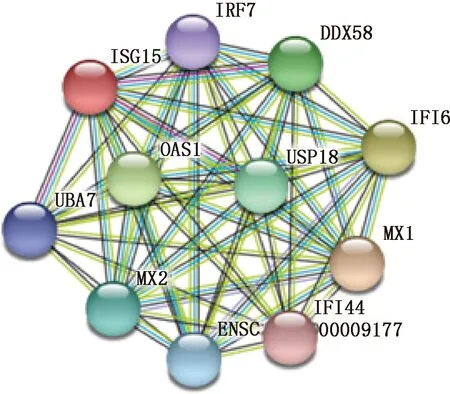
图7 山羊ISG15互作蛋白预测Fig.7 Prediction of proteins interacting with ISG15 in goats
2.3.5 蛋白功能预测 通过PredictProtein在线软件预测和分析山羊ISG15蛋白的功能,发现ISG15可能对细菌和病毒具有防御作用,通过负向调节病毒基因组的复制来达到防御病毒的目的;负向调节蛋白质泛素化;正向调节红细胞分化和造血过程;调节Ⅰ型干扰素和γ干扰素;调控白细胞介素10的产生;参与调节多种信号通路。
2.3.6 亚细胞定位预测 通过在线软件Psort Ⅱ Prediction对山羊ISG15蛋白亚细胞定位预测发现,山羊ISG15主要定位在细胞质中,在细胞质中发挥作用,可能性约占69.6%。同时山羊ISG15还可能分布在细胞核、线粒体和分泌系统的囊泡中,并在其中发挥作用。
2.4 IFA检测山羊ISG15蛋白在EEC细胞中的定位
通过IFA试验发现,转染ISG15后,该蛋白主要在细胞质中表达(图8A)。未用外源刺激物刺激,EEC细胞中无法检测到ISG15,用外源刺激物(PPRV)感染后,EEC细胞中可以检测到ISG15蛋白,并且也主要定位在细胞质中(图8B)。结果证实内源性与外源性ISG15均定位于细胞质中,此结果与软件预测结果一致。
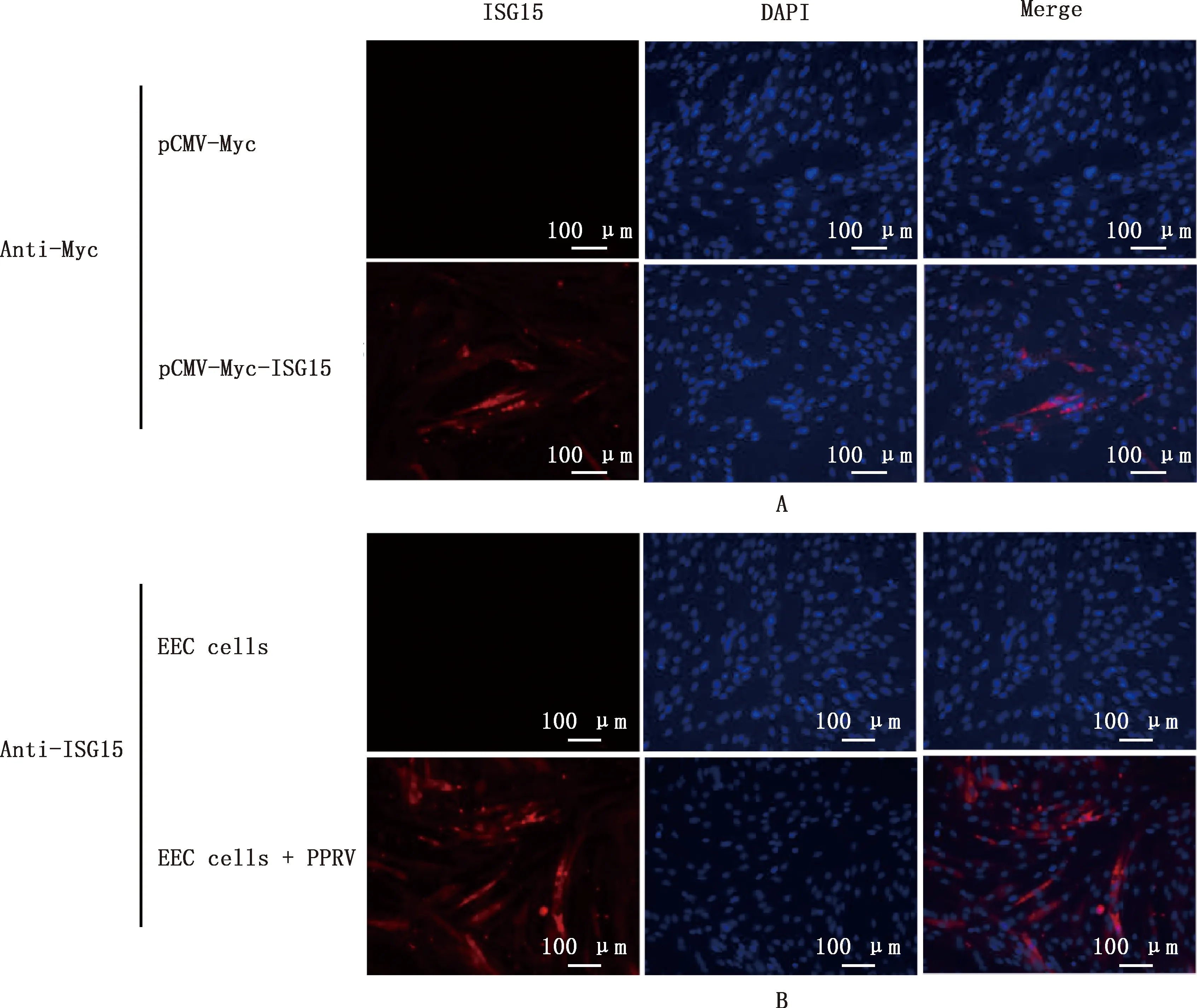
A,外源性ISG15蛋白在EEC细胞中的定位;B,内源性ISG15蛋白在EEC细胞中的定位A,Localization of exogenous ISG15 protein in EEC cells;B,Localization of endogenous ISG15 protein in EEC cells图8 山羊ISG15蛋白在EEC细胞中的定位(200×)Fig.8 Localization of ISG15 protein in EEC cells in goats (200×)
3 讨 论
ISG15是宿主对微生物感染反应的关键成分,本研究克隆了山羊ISG15的CDS区序列,通过氨基酸序列比对发现山羊和人该序列的相似性仅为65.2%,与盘羊相似性最高。以往的研究表明,某些哺乳动物和鱼类之间,相似性仅为30%~35%,即使两个不同的哺乳动物之间序列相似性也可能小于60%[16]。ISG15氨基酸的保守性分析发现,不同物种间ISG15长度不同,但在C-末端均含有LRLRGG序列,这对于翻译后修饰蛋白质至关重要。氨基酸序列差异性大,在山羊ISG15的50-80位氨基酸区域表现出明显的序列变异。序列的差异性导致相似性高的物种之间ISG15的性质也存在差异。鲁海富等[17]报道的新疆野生盘羊ISG15为疏水性蛋白,且有一个跨膜区,本研究通过分析发现,山羊ISG15为亲水性蛋白且没有跨膜结构。崔茹鹏等[18]报道的巴什拜羊ISG15基因编码区全长为522 bp,比本研究克隆的山羊ISG15基因长了48 bp。蔡卓轩等[19]报道牛的ISG15基因编码区为465 bp,比山羊的少了9 bp。序列的差异性会导致不同物种间ISG15的二级结构存在差异。山羊ISG15二级结构中无规则卷曲所占比例最多,约34.39%,β-转角仅占12.74%,而在新疆野生盘羊中存在10个β-转角[18],在牛中β-转角仅占17.53%[19]。
越来越多的证据表明,ISG15序列多样性会影响其三级结构,主要是保守的残基在结构域之间形成疏水界面,从而可能形成一系列的构象。人和小鼠的ISG15蛋白的三级结构整体构象存在着巨大的差异,在人ISG15中Asp79的羧基与Thr101的羟基形成氢键,在ISG15的铰链区形成一个扭结,但在小鼠ISG15中由于Ser77代替了Asp79,因此没有此现象[20]。与绵羊ISG15相比,牛ISG15的铰链区内缺乏3个残基导致牛ISG15在溶液中的稳定性较低,这可能与2个Ubl结构域的空间排列不同所致,后面研究证明牛ISG15两个Ubl结构域的扭曲方向完全不同[21]。因此解析绵羊ISG15的结构对于验证这些铰链区残基对三级结构的作用有着重要价值。
ISG15序列的多样性可以影响其生物学功能。与犬、小鼠和牛等其他物种的ISG15相比,乙型流感的NS1蛋白能够更有效地结合人和非人灵长类动物ISG15,这一发现表明,这可能是决定宿主易感性的主要因素[21]。这种现象同样在内罗病毒和冠状病毒中存在。内罗病毒的卵巢肿瘤家族蛋白(ovarian tumor family,OTU) 在体外与不同物种的ISG15相互作用不同[22]。冠状病毒的木瓜酶样蛋白酶(papain-like proteases,PLP) 与不同物种ISG15相互作用的能力上也有很大的差异,与人ISG15相比,小鼠ISG15的结合方向偏移了27°,蛋白酶结构也发生了相应的变化以适应它。重要的是,它还反映了不同冠状病毒之间已知宿主范围的差异[20]。ISG15对人和小鼠的USP18影响不同,与小鼠ISG15相比,人ISG15与USP18结合更强。研究者认为人和小鼠之间ISG15和USP18的序列差异是这种功能差异的根源[23]。蛋白亚细胞定位预测发现,山羊ISG15主要定位于细胞质中(69.6%),通过外源性转染ISG15真核表达质粒和PPRV感染刺激内源性ISG15的表达,发现ISG15定位于细胞质中,与预测结果相一致。同时本研究发现,未感染PPRV的EEC细胞中,通过IFA无法检测到ISG15的表达,表明PPRV感染可以刺激ISG15的表达。很多研究发现ISG15能够被病毒刺激上调表达,并发挥着其抗病毒作用[4,24]。该结果为后续研究ISG15在PPRV感染中的作用提供了基础。
ISG15是介导和调节宿主对病毒感染反应的重要组成部分。由于作用机理的差异,在不同病原体之间甚至在不同宿主物种之间的重要性有所不同。ISG15的序列和结构多样性增加了探索其在不同物种间作用的难度。不同物种间ISG15的生物信息学分析为后续的研究奠定了基础。
4 结 论
本研究成功克隆了山羊ISG15基因CDS区,全长474 bp,编码157个氨基酸。氨基酸序列比对发现山羊与盘羊ISG15序列相似性最高;山羊ISG15蛋白没有跨膜结构和信号肽区域,含有1个N-糖基化位点、16个O-糖基化位点和10个磷酸化位点;二级结构主要由无规则卷曲和延伸链构成;山羊ISG15蛋白可以与干扰素和泛素化相关蛋白相互作用;亚细胞定位发现内源性和外源性ISG15均定位于EEC细胞的细胞质中。
