烤烟上部烟叶成熟生理特性研究进展
2022-08-22贾中林尹启生戴华鑫梁太波张艳玲张仕祥
贾中林,尹启生,戴华鑫,梁太波,张艳玲,刘 阳,张仕祥
中国烟草总公司郑州烟草研究院,郑州高新技术产业开发区枫杨街2号 450001
烟叶成熟度是决定烤烟品质的核心要素,田间烟叶成熟采收是优质烟叶生产的重要环节,直接影响烤后烟叶品质。左天觉[1]、宫长荣[2]认为田间烟叶成熟采收对烟叶品质的贡献占整个烤烟生产环节的1/3。烤烟上部烟叶包括上二棚和顶叶在内的5~6片叶,占单株总产量的30%~40%,优质的上部烟叶对卷烟香味和风格有很大影响,是生产高档卷烟的重要原料,在烟叶生产中占有重要地位。长期以来,我国许多烟区上部烟叶存在成熟度不够、叶片僵硬、化学成分不协调、刺激性大、杂气重以及工业可用性差等问题,导致多数上部烟叶不能直接用于一、二类卷烟配方中[3]。自2009年朱尊权院士提出烤烟上部4~6片烟叶一次性成熟采收技术后,许多产区相继开展了提高上部烟叶可用性的技术研究与应用,并取得了显著效果,部分上部烟叶已应用到高端卷烟品牌配方中[4-5]。然而,目前关于烤烟上部烟叶成熟进程中的物质转化规律和相关作用机理等仍不明确,这在一定程度上限制了相关技术的应用。为此,从烟叶物质代谢、生物酶活性、基因表达和蛋白功能等方面综述了近年来烤烟上部烟叶成熟生理特性的研究进展,旨在为烟叶成熟度调控和优质上部烟叶开发提供参考。
1 上部烟叶成熟过程中物质代谢规律
1.1 碳水化合物
1.1.1 碳水化合物种类及测定方法
烟叶碳代谢产物主要是碳水化合物(糖类),按结构组成分为单糖、低聚糖和多糖。20世纪60年代起,国外开始关注烟叶中的碳水化合物,主要采用酸水解法测定烟叶淀粉含量,Somogyi法测定烟叶中可溶性糖含量,离子交换色谱法来分离烟叶中的糖组分[6-7]。20 世纪80 年代起,国内也开始相关研究,起初采用二硝基水杨酸比色法测定还原糖和总糖含量,酸水解-二硝基水杨酸比色法或酸水解-碘比色法测定淀粉含量[8-9]。随着分析技术的不断发展,出现了光谱法、液相色谱法、气相色谱法、离子色谱法和电泳法等更加快速精准的检测方法[10-11],目前行业内对烟叶中总糖和淀粉的测定一般采用连续流动分析法,测定糖组分一般采用高效液相色谱法(表1)。

表1 烟叶中碳水化合物的检测方法Tab.1 Detection methods for carbohydrates in tobacco leaves
1.1.2 碳水化合物与烟叶成熟度的关系
烟叶成熟过程中碳代谢产物的研究主要集中于总糖、还原糖和淀粉(表2)。在烟叶成熟过程中,叶片光合生理功能开始衰退,碳水化合物合成减弱而降解增强,物质含量逐渐降低[12]。国外研究表明,随烟叶成熟度增加,淀粉、总糖和可溶性糖含量大致呈抛物线形变化,国内研究也发现同样的规律,并认为烟叶淀粉含量的最大值出现在烟叶工艺成熟时期,之后进入过熟阶段,烟叶淀粉含量逐渐降低[13-14]。烟叶成熟过程中物质含量的变化可为生产上烟叶成熟度的判断提供参考,如美国通过提前1周测定烟叶中化学成分含量来判断烟叶的成熟度,以此来确定烟叶采收期[15]。20世纪80年代起,国内学者也提出将淀粉含量作为烟叶成熟度的判断指标,淀粉含量达到最高值的时间即为烟叶工艺成熟期[18]。烟叶碳代谢产物在成熟过程中的变化受品种、地域环境等因素的影响。不同品种烤烟成熟过程中,烟叶淀粉和还原糖含量虽然整体上表现为先升高后降低的变化趋势,但达到峰值的时间存在差异[22]。同一烤烟品种在不同地域环境下,烟叶糖含量在成熟过程中呈现的变化规律也略有差异[23]。目前对烟叶成熟过程中糖含量的研究主要集中在总糖和还原糖总量方面,而对糖组分如葡萄糖、果糖、蔗糖和麦芽糖等的关注相对较少。

表2 国内外烟叶成熟过程中碳代谢产物的变化主要研究结果Tab.2 Domestic and foreign researches on variations of carbon metabolites during tobacco leaf ripening
1.2 含氮化合物
1.2.1 含氮化合物种类及测定方法
烟叶中氮代谢产物主要是含氮化合物,包括蛋白质、游离氨基酸和生物碱等,含氮化合物对烟叶的感官质量和吸食者健康有重要影响。国外对烟叶含氮化合物的研究始于20世纪60年代,最初主要采用凯氏定氮法测定蛋白质,该方法具有操作简单、测量准确等优势,也是较长一段时期国内测定蛋白质的常用方法。目前,行业内总氮的测定主要采用连续流动分析法[24]。1958 年,Moore 等[25]发明了氨基酸自动分析的方法,逐渐成为国内外测定氨基酸的常用方法。随着研究的深入和分析仪器的发展,出现了电泳法、光谱法、色谱法、质谱联用等更加快速准确检测烟叶游离氨基酸的方法[26-27](表3)。国外对生物碱的测定方法研究较早,Bertrand 和Javillier 采用硅钨酸重量法测定了烟碱;之后Markw 提出了1种烟碱比色测定的方法[28]。国内对烟碱含量的测定最初采用酸碱滴定法,之后出现了光度法、电泳法、液相色谱、气相色谱、液质联用或气质联用等更精准的测定方法[28-30],目前行业内主要采用连续流动分析法大批量检测样品中生物碱含量(表4)。
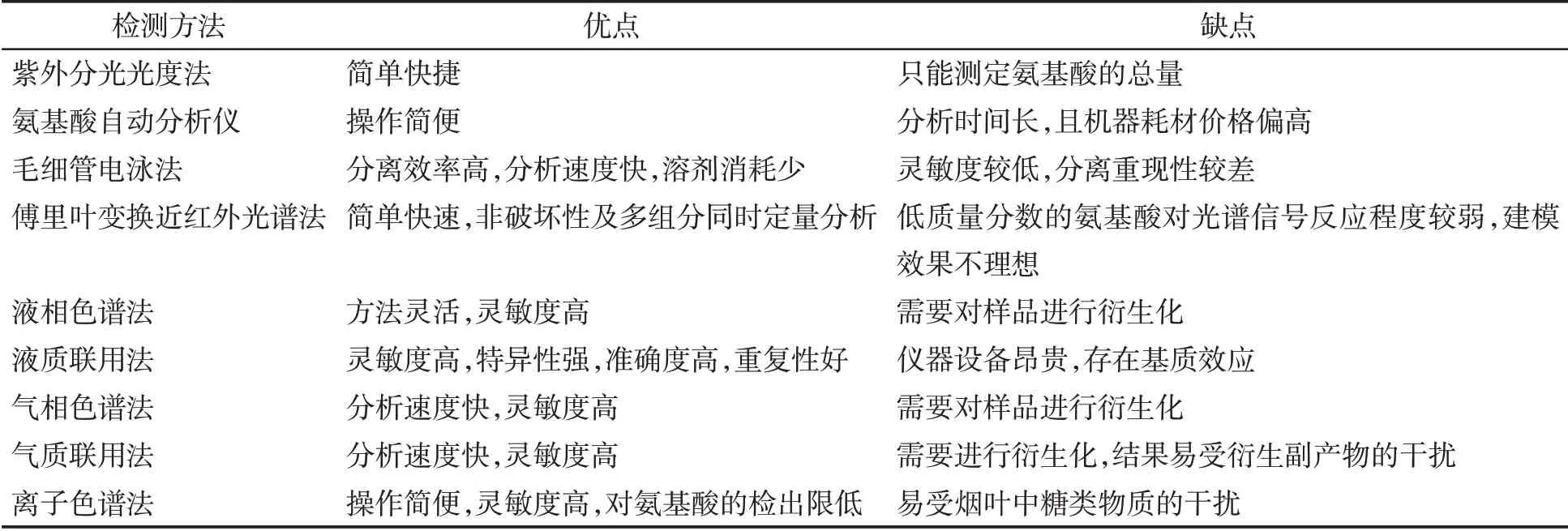
表3 烟叶中游离氨基酸的检测方法Tab.3 Detection methods for free amino acids in tobacco leaves
1.2.2 含氮化合物与烟叶成熟度的关系
烟叶成熟过程中氮代谢产物的研究多集中在总氮、蛋白质、氨基酸和生物碱(表5)。随着烟叶成熟度增加,总氮、蛋白质和游离氨基酸含量逐渐降低,这一变化与烟叶衰老过程中氮素的再分配和再利用有关[31]。叶片中的氮素大部分以蛋白质形式存在于叶绿体中,在叶片进入衰老阶段,蛋白水解酶活性显著增强,蛋白质被水解为氨基酸等物质,然后通过韧皮部输送到新生器官中以维持其生长,因此蛋白质等含氮化合物含量在烟叶成熟衰老过程中逐渐降低[32]。在烟叶生长发育过程中,生物碱在叶片中逐渐积累,各组分(烟碱、降烟碱、新烟草碱和假木贼碱)含量均随成熟度增加而呈上升趋势[33]。游离氨基酸可作为前体物参与烟碱的生物合成,随着烟叶叶龄的增加,氮在游离氨基酸中的分配比例逐渐降低,而在烟碱中的分配比例逐渐提高,成熟后期烟叶游离氨基酸含量降低,也可能与其参与了烟碱的生物合成有关[31,34]。有研究表明在烟叶成熟后期,烟碱含量出现下降趋势,可能是由于烟叶达到了过熟状态,叶片开始衰老,导致部分内含物质发生降解而损耗[35]。有研究认为,烟叶游离氨基酸含量与烟叶品质呈负相关,在烟叶成熟过程中游离氨基酸含量呈先降低后升高的变化规律,最低值出现在烟叶工艺成熟时期,烟叶中游离氨基酸含量可以作为判断烟叶成熟度的依据之一[18]。与碳水化合物相似,烟叶成熟过程中含氮化合物的变化规律在不同品种、生态环境和栽培条件下也略有差异。因此,在实际生产中应针对产区特点,探究烟叶成熟过程中化学成分与成熟度的关系,确定烟叶最佳采收期,将烟叶化学成分控制在适宜范围内。

表5 国内外烟叶成熟过程中含氮化合物的变化主要研究结果Tab.5 Domestic and foreign researches on variations of nitrogen metabolites during tobacco leaf ripening
1.3 致香物质
1.3.1 致香物质的种类及测定方法
烟叶致香物质含量是衡量烤烟品质的重要因素之一。国外早在20 世纪50 年代就已经从烤烟中分离鉴定出多种重要的香味成分如茄尼醇、新植二烯和挥发性脂肪酸等[41]。1976年,Lloyd等[42]借助蒸馏萃取和气相色谱/质谱联用技术(GC/MS)从烤烟烟丝中分离鉴定出酮类、羧酸类、醇类、醛类、酚类和氮杂环类等共计323 种化合物。国内对烤烟致香成分的研究较晚,1989年,李云等[43]利用气相色谱技术从烤烟中鉴定出12种低级脂肪酸;1992年,冼可法等[44]从云南烤烟中鉴定出129 种中性致香成分。随着前处理技术以及检测技术的不断发展,同时蒸馏萃取、固相微萃取、液液微萃取、超临界CO2萃取、加速溶剂萃取以及基于气质和液质联用的代谢组学分析手段逐渐应用到烟叶致香成分分析中,大量致香成分被分离和鉴定出来[45]。
1.3.2 致香物质与烟叶成熟度的关系
一般认为在烟叶成熟过程中,类胡萝卜素、类西柏烷和苯丙氨酸等致香前体物的降解产物以及棕色化反应产物含量总体呈升高趋势,达到最大值后开始下降,但由于烟叶致香物质受生态因素影响较大,不同产区的研究结果存在较大差异,例如在四川省攀西地区,随着成熟度增加,烟叶棕色化产物含量逐渐降低[46],而在陕西省延边地区,烟叶棕色化产物总量则呈上升趋势[47]。此外,不同品种的致香物质成分组成也存在差异,在烤烟NC89烟叶成熟过程中检测到糖苷类香气成分共28种,包括醛、酮、醇、内酯和酚类等成分,而在云烟87烟叶中仅分离鉴定出21种糖苷类香气成分,主要由脂肪醇、芳香族成分、倍半萜类和多酚类物质组成,两个烤烟品种烟叶中糖苷类香气成分总量均随成熟度的提升而增加[48-49]。目前,关于烟草致香物质的研究大多集中在烤后烟叶中,对于田间鲜烟叶尤其是上部烟叶成熟过程中的变化研究较少,虽然在烟叶烘烤过程中致香物质含量还会发生新的变化,但田间采收烟叶中致香物质及前体物的基础含量对烟叶品质起着决定性作用。因此,选择合理的采收期,保持适宜的烟叶致香物质及前体物含量,对改善烤后烟叶香气品质具有重要意义。
1.4 质体色素
烟叶成熟过程中最直观的变化是颜色的改变,生产上叶片颜色是判断成熟度的重要依据。烟叶颜色的变化实质上是质体色素含量变化的外在表现,质体色素主要包括叶绿素和类胡萝卜素等。叶绿素含量下降而导致的叶片失绿被认为是植物衰老最显著的特征,其与叶绿体中类囊体膜逐渐崩解有关[50]。观察细胞超微结构(图1)发现,在烟叶成熟过程中,细胞内叶绿体首先出现衰老症状,具体表现为细胞空隙变大,叶绿体肿胀呈不规则形状,基粒个数和类囊体数量逐渐减少,类囊体膜结构丧失,淀粉粒和嗜锇颗粒数量增多、体积增大,并向细胞中部游离[50-51]。
在烟叶成熟过程中,叶绿素、类胡萝卜素和质体色素总量均随成熟度增加而逐渐降低,其中叶绿素较类胡萝卜素含量下降速率更快,降解量更大,二者之间的比例变化使不同成熟度烟叶颜色产生差异[52]。由于烟叶叶绿素含量在成熟过程中变化显著且易于测定,所以常被作为衡量烟叶成熟度的重要指标。叶绿素仪可以快速检测叶片中叶绿素相对含量(Soil and Plant Analyzer Development,SPAD),生产上可将烟叶田间SPAD值与其外观颜色特征相结合,作为判断烟叶成熟度的依据[53-54]。然而,SPAD仪虽然操作简单、使用方便,但只能进行单点测量,测量误差和样本代表性的缺陷,限制了其大范围推广使用。近年来,多种现代信息获取技术如计算机视觉技术、无人机遥感技术、高光谱技术和近红外光谱技术等逐渐在烟草生产中得到应用,通过建立光谱信息与烟叶成熟度的关系模型,可以实现烟叶成熟度的可视化分析[55-57]。无人机遥感和高光谱等技术虽然具有测量范围大、精准度高等优势,但技术门槛和成本较高,现阶段并未广泛应用于烟叶成熟度判断。因此,根据田间烟叶理化指标的变化选择更加科学合理的成熟度量化指标和便捷高效的检测方法、检测设备,将是今后需重点解决的问题。
2 上部烟叶成熟过程中生物酶活性变化
2.1 碳代谢相关酶
植物的碳氮代谢过程离不开各种生物酶的参与,在烟叶生长发育过程中,参与碳代谢的生物酶有淀粉合成酶(Starch synthase,SS)、淀粉酶(Amylase,AM)、蔗糖转化酶(Invertase,INV)和蔗糖合成酶(Sucrose synthase,SUS)等[58]。AM 是碳水化合物积累代谢过程中的关键酶,可以将烟叶中积累的淀粉降解为麦芽糖和少量葡萄糖,目前关于烟叶AM 活性在成熟过程中的变化主要有3 种观点:①双峰变化,但达到峰值的时间还存在分歧[9,12];②先升高后降低的单峰变化[58-59];③先降低后升高的“U”形曲线变化,最低值时间与淀粉含量达到最高值的时间相吻合[21],结果的差异主要与烤烟品种、生态环境及取样标准等不同有关。蔗糖是植物体中碳水化合物运输的主要形式,INV 可以催化蔗糖水解为葡萄糖和果糖,其活性大小反映了烟叶对光合产物的利用程度。杨志晓等[59]研究表明,烟叶INV 活性在成熟前期较低,呈缓慢增加趋势,在成熟期活性迅速上升,不同品种间INV活性存在较大差异。INV活性前期较低,可能是因为烟叶前期氮代谢活动占主导,碳代谢活动强度较弱,进入叶片功能盛期后,氮代谢活动开始减弱,转为以碳代谢为主,转化酶活性增强有利于促进光合产物的合理分配。
2.2 氮代谢相关酶
植物的氮代谢包括氮素的吸收、同化、转运、利用和调节等过程。参与氮代谢的生物酶有硝酸还原酶(Nitrate reductase,NR)、谷氨酰胺合成酶(Glutamine synthetase,GS)、谷氨酸合成酶(Glutamate synthase,GOGAT)和谷氨酸脱氢酶(Glutamate dehydrogenase,GDH)等[59]。1952 年,Evans 等[60]在红色链孢霉中首次发现NR,次年又在高等植物中发现。NR 是植物氮代谢的关键酶和限速酶,其作用是将植物吸收的硝态氮还原为亚硝态氮以供后续利用。2000年,Becker等[61]将植物GS分为2 类,细胞质型(GS1)存在于根部细胞质中,主要同化植物根系系统中硝酸还原作用产生的氨;质体型(GS2)存在于绿色组织质体中,负责同化硝酸还原和光呼吸所产生的氨。GS 与GOGAT 联合作用下,将植物吸收的无机态氮同化为有机态的谷氨酰胺和谷氨酸,其活性大小可反映植物对氮素的同化利用情况[62]。随着烟叶成熟度增加,NR活性先升高后降低,在移栽后30~60 d活性较强,在60~70 d后活性逐渐减弱,亚硝酸还原酶活性与NR活性变化规律大致相同[40]。贾保顺等[63]研究发现,烤烟K326 和NC71的GS活性在移栽后70~110 d内呈先升高后下降的变化,最高值出现在移栽后80 d,叶片中GDH活性、NH4+浓度与GS活性变化趋势相同。氮代谢相关酶活性前期升高可能与叶片正处在功能盛期有关,此时氮代谢活动较强,后期碳代谢活动逐渐增强,氮代谢活动开始减弱。在烟叶成熟进程中,碳氮代谢的强度、协调程度以及动态变化会直接影响烟叶中各类化学成分的含量和比例,进而影响烟叶品质。优质烟叶生产的关键是在适当发育时期,及时由以氮代谢为主转变为以碳积累代谢为主。因此,生产上可采用生物、化学和农艺等措施有针对性地对碳氮代谢相关酶活性进行调节,以协调碳氮代谢进程,提高上部烟叶化学成分协调性,改善烟叶品质。
2.3 衰老相关酶
衰老是植物发育过程中细胞、组织、器官和个体死亡的过程。在众多衰老机制学说中,生物自由基导致衰老的学说比较受重视[64]。植物体内的超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT)和过氧化物酶(Peroxidase,POD)等是植物应对胁迫时重要的防御酶系,可以有效阻止活性氧自由基的积累[65]。SOD是生物体转移清除超氧阴离子自由基(O2-·)的酶,能特异性地将O2-·歧化为H2O2和O2[66]。CAT 是一种包含血红素的四聚体酶,在烟草中分离到的3个CAT同工酶中,Cat1主要负责清除光呼吸产生的H2O2,Cat2 则清除氧化胁迫产生的H2O2,而Cat3 主要清除乙醛酸循环体中产生的H2O2[67]。植物中的过氧化物酶有很多种,最常见的有抗坏血酸过氧化物酶、谷胱甘肽过氧化物酶和辣根过氧化物酶等。利用电泳法从辣根组织中分离出2 种不同的辣根过氧化物酶,在活性氧代谢过程中,POD可以清除光合作用过程中产生的过量H2O2,起到保护叶绿体的作用[68]。
植物衰老过程其实也是抗氧化酶活性下降、活性氧自由基积累、膜脂过氧化不断加剧的过程。在烟叶衰老过程中,SOD、CAT和POD的活性一般呈逐渐降低的趋势,但也有研究发现,在烟叶成熟前期抗氧化酶活性有短暂升高的现象[69-70]。此外,一些参与烟株防御体系的生物酶,如脂氧合酶和多酚氧化酶等在烟叶成熟过程中也呈现先升高后下降的变化趋势[70-71]。成熟前期烟株代谢旺盛,活性氧的少量积累对植物细胞损害较小,反而诱导了抗氧化酶活性的升高,但在成熟后期,烟株生理功能逐渐衰退,活性氧过量积累造成细胞大量损伤,抗氧化酶活性也随之降低[72]。外界环境因素的改变会影响烟叶衰老进程,如高温、冷害、盐胁迫等在一定程度上会加速烟叶衰老。喷施外源调节物质,如水杨酸、硅、2,4-表油菜素内酯等可以调节植物叶片抗氧化酶活性,完善植物体内的防御体系,缓解恶劣环境胁迫对烟株造成的伤害[73-74]。因此,生产上遭遇环境胁迫条件时,可以根据大田烟株生长实际情况,选择适宜的外源调节物质进行喷施,以增强成熟期烟株的抗逆性。
3 上部烟叶成熟过程中基因表达变化
转录组是指细胞中所有参与蛋白质翻译的mRNA 总和,自20 世纪90 年代起,随着分子生物技术和高通量测序技术等的快速发展,转录组学技术已广泛应用于各个学科领域,借助转录组学分析技术,科研人员在植物叶片衰老的分子机制方面取得了重大进展。1995 年,Gan 等[75]将农杆菌ipt基因在衰老启动子SAG12 的驱动下在烟草中表达,延缓了烟叶衰老的进程;2001 年,Yang 等[76]发现烟草中两种碱性亮氨酸拉链(b-ZIP)家族的转录因子表达会促进叶片衰老。目前,已从烟草和拟南芥等模式植物中克隆出大量与衰老相关的基因,根据这些基因在叶片衰老期间表达量的变化,可将其分为衰老上调基因和衰老下调基因[77]。相关基因在烟叶成熟过程中的功能主要集中在蛋白质降解、碳氮代谢、色素代谢和激素代谢等方面。
3.1 蛋白降解相关基因
半胱氨酸蛋白酶是一种多功能水解蛋白酶,在植物衰老和细胞程序性死亡过程中起关键作用,烟草中NtCP1、NtCP2、NtCP23和MC是编码半胱氨酸蛋白酶的基因[50]。2006 年,Beyene 等[78]从烟草衰老叶片和非衰老叶片中分离得到两个与衰老相关的半胱氨酸蛋白酶基因NtCP1和NtCP2,NtCP1是一种特异性表达基因,仅在自然衰老的叶片中表达,且不受外界不良条件的诱导,被认为是烟草中良好的衰老标记基因,NtCP2与NtCP1表达模式不同,其在成熟绿叶中表达量较高,而在衰老叶片中表达量显著降低。在衰老叶片中,包括NTCP23在内的许多编码蛋白酶基因上调以参与氮素的转移活动。2000 年,Ueda等[79]从烟草中分离出编码半胱氨酸蛋白酶的基因NtCP23,其表达模式与NtCP1相似,在衰老过程中上调表达,但在叶片生长发育初期也能检测到NtCP23的表达。PSA1是编码26S 蛋白酶体的非催化型亚基,在烟叶衰老过程中其表达量逐渐降低,有学者推测蛋白酶体在植物细胞中的作用可能是调节蛋白水平来控制发育过程,而不是在衰老过程中降解蛋白质[50]。在烟叶衰老过程中,NtCP1、NtCP23、PSA1和MC在不同烤烟品种中的变化规律基本一致,基因NtCP1和NtCP23在衰老过程中上调表达,PSA1和MC基因下调表达。不同烤烟品种之间的差异在于各基因表达量的不同,成熟落黄快的品种NtCP1和NtCP23的表达量较高,PSA1和MC的表达量较低[22]。
3.2 碳氮代谢相关基因
关于烟叶成熟过程中碳氮代谢相关基因的研究主要以编码生物酶的基因为主。颗粒结合淀粉合成酶(GBSS)和淀粉分支酶(SBE)是淀粉合成的关键酶,分别参与直链淀粉和支链淀粉的合成,他们由相应的淀粉合成酶基因GBSS1、SBE等编码。在烟叶成熟过程中,GBSS1和SBE基因的相对表达量呈先升高后下降的变化,与淀粉含量在成熟过程中的变化一致[22]。蔗糖合成酶基因(NtSS)、蔗糖磷酸合成酶基因(NtSPS)和蔗糖转化酶基因(NtINV)参与烟叶的糖代谢过程。在烟叶成熟过程中,NtINV表达量大致呈单峰曲线变化,NtSS和NtSPS表达量均有不同程度上调,且品种之间变化规律一致,说明烟叶进入成熟期后糖代谢活动逐渐增强[80]。在氮代谢方面,NR 是整个氮代谢过程中的限速酶和关键酶,其编码基因在叶片衰老过程中表达量逐渐降低[81]。周健飞等[82]指出,在烟叶成熟过程中,NR 基因的表达丰度仅与NR 活性呈极显著正相关,对成熟期氮效率影响较小,成熟期烟叶氮素运筹主要受GS同工酶基因表达丰度的影响。在烟叶衰老过程中,GS1 调控氮素的转移及再利用,GS2 负责氨同化作用[83],GS1 编码基因表达量呈上升趋势,GS2 编码基因表达量则逐渐下降,表明随着烟叶衰老其氮素转移能力逐渐加强,而氮素同化能力逐渐减弱[22,82]。GDH在烟叶衰老过程中主要负责脱氢,编码GDH的基因有GDH1和GDH2,它们的表达量在烟叶衰老过程中均增加,其转录水平在衰老后期最高[50],但也有研究发现GDH1的相对表达量在衰老过程中呈先升高后降低的趋势[81],相关差异的原因还需进一步研究。
3.3 色素代谢相关基因
在烟叶成熟过程中,与类胡萝卜素合成相关的基因GGPS、PSY、PDS、ZDS和CRTISO均呈现下调表达,相关转录因子ORANGE、HY5、COP1和DET1也呈现下调表达。与之相反,类胡萝卜素转化基因NCED、ZEP、NXS以及与类胡萝卜素降解相关的基因LOX、POD、CCD1则上调表达[84]。叶绿素酶是分解叶绿素的初始酶,CLH是编码叶绿素酶的基因,在烟叶衰老过程中,CLH表达整体增强,说明在烟叶成熟过程中,叶绿素与类胡萝卜素的分解代谢逐渐增强,类胡萝卜素的合成逐渐减弱,这与烟叶中质体色素含量降低的变化规律一致[85]。目前,从拟南芥和水稻中鉴定出6种叶绿素代谢酶[86],相关基因在叶片衰老过程中的变化在其他作物上有较多研究[86-87],而在上部烟叶成熟过程中的变化研究则相对较少。
3.4 激素代谢相关基因
内源激素是植物体内重要的信号物质,对叶片的成熟衰老进程起重要调控作用。烟叶成熟过程中,参与乙烯、脱落酸和茉莉酸合成的相关基因NtEFE26、NtNCED和NtPR1b表达量均呈升高趋势,在衰老烟叶中达到最高值,与相应激素含量的变化趋势相同[88]。生长素(IAA)和细胞分裂素(CTK)被认为是延缓衰老的激素,在烟叶衰老过程中,IAA负调控因子AuX/IAA与SAUR家族蛋白的相关基因表达量均呈升高趋势,IAA合成受到抑制,这与IAA含量在烟叶衰老过程中下降的变化相一致[89]。在衰老叶片中CTK含量显著下降,转录组分析结果表明,与CTK降解有关的基因上调表达,而与CTK合成相关的基因则下调表达[90]。NtCHN50是水杨酸应答基因,在烟叶成熟过程中其表达量逐渐降低,预示着水杨酸合成减弱[88],但对拟南芥的研究发现,在衰老叶片中水杨酸的浓度提高了4倍,衰老叶片中较高的水杨酸含量似乎与叶片衰老过程中几种SAG的上调表达有关[91],差异的原因还需进一步研究。
4 上部烟叶成熟过程中蛋白表达变化
4.1 蛋白组学技术在烟草上的应用
自20世纪90年代起,伴随着蛋白组学技术体系的不断发展,蛋白组学技术已广泛应用于植物不同生长发育阶段以及在各种环境因子和逆境胁迫下的蛋白表达差异研究中[92]。2008年,Hebeler等[93]用蛋白组学技术分析了拟南芥早期衰老叶片的蛋白丰度变化,鉴定出13个差异显著的蛋白。2010年,Bai等[94]研究了牵牛花衰老过程中蛋白组的变化,发现许多衰老上调蛋白参与了防御和应激反应等生物活动。蛋白组学技术在烟草上的应用起步相对较晚,目前主要集中在不同基因型、不同生态环境以及胁迫条件下差异蛋白的研究。2008 年,崔红等[95]对浓香型和清香型典型生态区的烟叶进行蛋白组学比较分析,发现有51个蛋白在两个生态区发生了差异表达,在福建烟区高表达的蛋白主要参与叶绿体发育、色素代谢和光合作用,而在河南烟区高表达的蛋白主要参与糖代谢途径,为从蛋白水平解释不同生态区烟叶品质差异提供了参考。2016 年,徐莹等[96]对云烟87、K326和红花大金元成熟期烟叶蛋白组分析发现,云烟87与烤烟K326烟叶中蛋白质表达图谱相似,仅存在29 个差异表达蛋白,而云烟87 和红花大金元之间则有160个蛋白质差异表达,差异蛋白多与光合作用、物质代谢和抗逆性有关。2019 年,Chen等[97]基于同位素相对标记和绝对定量(iTRAQ)的蛋白组学分析研究表明,豫烟6号和云烟87 在水分胁迫下蛋白组表达水平存在差异,与云烟87相比,豫烟6号表现出更强的保水能力、更好的光合性能、更强的抗氧化防御能力,为研究烟草干旱胁迫反应相关的遗传调控机制奠定了基础。
4.2 烟叶成熟衰老过程中的蛋白组学研究
近年来,在蛋白水平上解释烟叶成熟过程中的成分变化逐渐受到人们的关注。2014 年,张柳等[98]采用蛋白组学技术对云南玉溪地区云烟87旺长期、成熟期和成熟后期烟叶的蛋白组进行了分析,共检测到432 个差异表达蛋白,主要集中在以下几个方面:(1)光合作用,如与光反应相关的蛋白质(如PSⅠ和PSⅡ反应中心蛋白等)和与暗反应中CO2同化相关的蛋白(Rubisco及其活化酶等)在衰老过程中均呈下调表达。(2)糖酵解代谢,如α-葡聚糖焦磷酸化酶、α-葡萄糖苷酶类、与糖类转化及糖核苷酸合成相关酶类均上调表达。(3)蛋白质和氨基酸分解代谢,如天冬氨酸蛋白酶、天冬氨酸转氨酶等上调表达。(4)逆境反应,如热激蛋白和分子伴侣总体上调表达。(5)脂类代谢,如甘油酯水解酶类和磷脂类水解酶表达水平升高。这些现象表明,在烟叶衰老过程中叶片光合作用强度逐渐减弱,呼吸代谢、物质分解代谢活动逐渐增强,同时抗逆相关蛋白表达的升高说明烟叶衰老对植物体本身也是一种逆境反应。何聪莲等[99-100]采用iTRAQ对烤烟K326未熟、适熟和过熟期烟叶的蛋白组进行分析,共鉴定出86个共同差异表达蛋白,这些差异蛋白涉及代谢过程、细胞过程和应激反应等,最终筛选出 包 括Q1WL43、A0A1S4DCM1、A0A1S4C433、A0A1S4AKK6、A0A1S3ZQP6 和A0A140G1P2 在内的6个差异蛋白,可能是烟叶衰老潜在靶标蛋白。
5 总结与展望
目前,我国在上部烟叶成熟度研究和应用方面仍存在一些问题:①关于成熟度与烟叶品质关系的研究,主要集中在田间烟叶成熟度对烤后烟叶品质的影响,而对成熟过程缺乏关注,田间烟叶成熟过程中物质转化规律和内在生理生化研究不够深入。②上部烟叶的采收成熟度判断仍然靠主观经验,缺乏统一的特征指标、判断标准和便捷的测定方法,在生产操作中不易把控,上部烟叶采青现象仍然存在;部分产区由于过分强调成熟度,导致上部烟叶出现物质消耗过度、品质下降等问题。③国内许多产区主要采用延迟采收技术提高烟叶成熟度,由于在烟叶生长后期部分产区气温下降较快,导致烟叶表面易形成蜡质层,使烟叶难以成熟,给烘烤带来困难。烟叶生育后期降水增多带来的病害问题也限制了该项技术的应用。④在外源调节剂调控上部烟叶开片和成熟度方面研究尚不深入,应用也较少。
针对上述问题,未来可先从典型产区和主要栽培品种入手开展相关研究。①借助转录组、蛋白组和代谢组学的相关技术手段,从关键酶活性、基因表达、蛋白质和生物小分子代谢物等多角度出发,系统研究上部烟叶成熟过程中的物质代谢规律,筛选烟叶成熟过程中起重要调控作用的关键基因和代谢产物,并进行功能验证和机制解析。②深入开展烤烟上部烟叶成熟调控研究,研制筛选对烤烟上部烟叶成熟具有较好调控作用的生长调节剂,并揭示其作用机理。③选择适宜种植区域,保证烟株营养均衡,提高烟叶耐熟性,以烟株营养均衡+成熟采收为核心提高上部烟叶品质。④在烟叶田间成熟度特征指标、判断标准、测定方法和测定设备等方面加强研究,针对不同的生态区域,形成上部烟叶成熟度判断和采收标准,制定精准的上部烟叶生产技术规范。
