我国余甘子新害虫锡兰玻壳蚧的形态及生物学特性
2022-08-13黄佳聪杨晏平李归林
蒋 华, 黄佳聪, 杨晏平, 李归林
(1. 云南省保山市林业和草原技术推广站, 云南保山 678000; 2. 云南省保山市林业和草原科学研究所, 云南保山 678000)
锡兰玻壳蚧Drepanococcuschiton隶属半翅目(Hemiptera)蚧总科(Coccoidea)蜡蚧科(Coccidae)玻壳蚧属Drepanococcus,是我国近年来新记录的一种入侵害虫。该虫最早发现于印度南部锡兰岛(现斯里兰卡)番石榴Psidiumguajava的果实上(Green, 1908),1909年由Green描述置于Ceroplastodes属(Green, 1909),1990年由Williams和Watson重描归于Drepanococcus属(Williams and Watson, 1990)。近年来锡兰玻壳蚧已在印度、马来西亚、泰国、越南和老挝等国传播扩散,严重危害香蕉Musanana、杨桃Averrhoacarambola、番石榴Psidiumguajava、番木瓜Caricapapaya、榴莲Duriozibethinus和龙眼Dimocarpuslongan等热带水果植物(Ibrahim, 1994; Manietal., 1997; Sharma, 2011; Choietal., 2018; Tranetal., 2019)。2020年国内首次报道广州南沙区白楸Mallotuspaniculatus上发现该害虫(仇玲, 2020)。2019年,笔者在调查滇西地区余甘子病虫害时采得此虫,该虫主要寄生于枝干、脱落枝、叶柄等部位,以雌成虫和若虫吸食汁液危害,造成受害植株长势衰弱,枝条生长缓慢或失水干枯,导致果实品质和产量降低。 在滇西海拔770~1 750 m余甘子适生栽培区受害株率达32%~56%,为害面积约213 hm2,呈逐年上升趋势,对滇西地区农业生产及生态环境具有潜在的巨大威胁。
目前国内外已有关于锡兰玻壳蚧形态学和生物学方面的研究,Ibrahim(1994)介绍该虫在马来西亚29℃环境下可存活50 d,产卵量达1 200粒/雌。Tran等(2019)报道该虫在越南可严重为害番木瓜,导致果实完全丧失商品价值。Mani(1995)在印度班加罗尔杨桃和番石榴果园中调查出6种该害虫的寄生昆虫。仇玲(2020)对其末龄若虫和雌成虫的形态、蜡腺和蜡泌物超微特征进行了系统的研究。而关于锡兰玻壳蚧在我国果树上发生为害、各虫态特征及相关生物学等研究尚处空白。本研究详细调查了锡兰玻壳蚧在我国滇西热带、亚热带干热河谷地区的发生为害、空间分布、形态特征、生活史及相关生物学特性,以期能为该害虫的预测预报及科学防控提供参考依据。
1 材料与方法
1.1 样地概况及试虫来源
以云南省保山市隆阳区潞江镇(丛岗村、孟柳村)、蒲缥镇双河村余甘子栽培林(表1)为样地进行锡兰玻壳蚧种群空间分布格局调查,并采集锡兰玻壳蚧雌成虫,以余甘子无病虫新鲜嫩枝叶和锡兰玻壳蚧雌虫寄生枝条嫁接作为寄主,分别进行室内培养和仿生饲养。
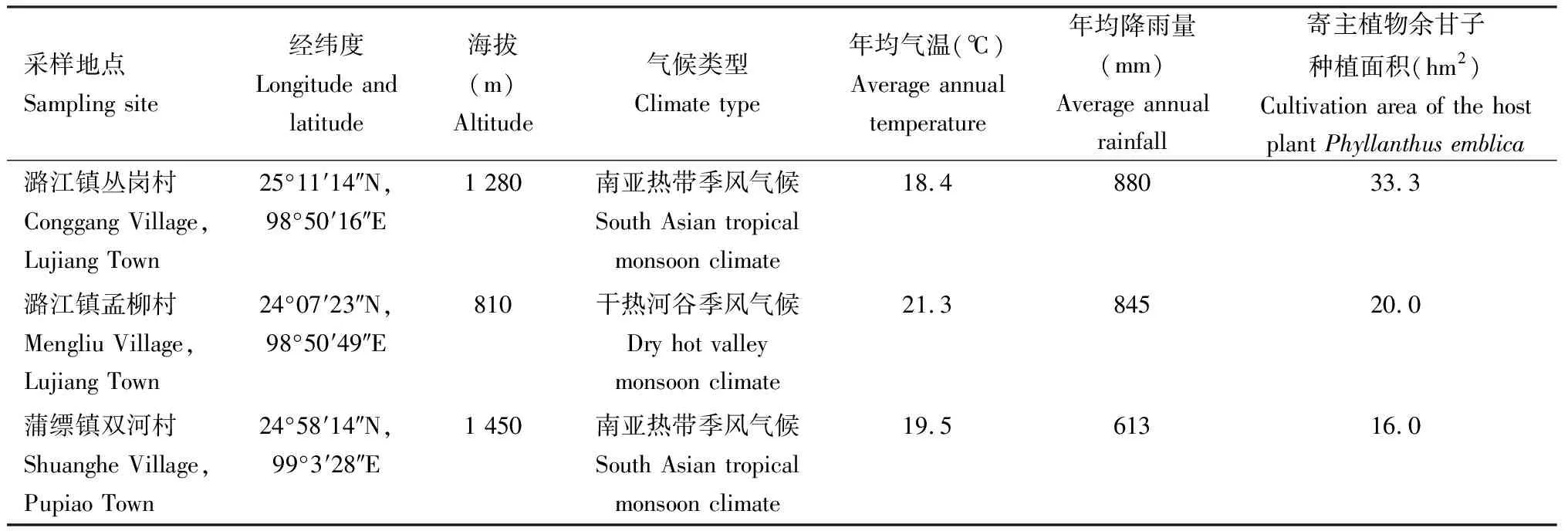
表1 云南保山样地概况Table 1 Overview of sampling sites in Baoshan, Yunnan
1.2 林间发生为害及空间分布型调查
1.2.1调查方法:2019年5-9月在3块虫源余甘子栽培林中,每林地采用五点取样方法,分别设置单块面积大于800 m2具有代表性的调查样地5个,每样地内随机调查30株,详细记录每株所有枝条寄生锡兰玻壳蚧雌成虫的数量及为害情况。
1.2.2空间分布型分析方法:采用聚集度指标法(于新文和刘晓云, 2001),计算各林地调查样株的锡兰玻壳蚧平均虫口密度(m)和方差(S2),计算分析各项聚集度指标,分析测定空间分布型(表2)。

表2 聚集度指标计算公式及判别标准Table 2 Calculation formula and criterion of aggregation indexes
1.3 形态特征及生物学特性观察
1.3.1室内培养:于2020年6月在蒲缥镇双河村余甘子栽培林样地内剪取受害枝条,网袋收集密封后带回实验室。从中挑取60头锡兰玻壳蚧产卵雌成虫,在OLYMPUS SZ体视显微镜下拨开蜡壳观察记录其产卵量。挑取初产卵540粒置于垫有湿润纱布直径9 cm的培养皿(180粒/皿),将培养皿置于HWS培养箱(均温26±1℃, RH 70%±7%, 光周期8L∶16D),每1 d取盖置于体视显微镜下观察胚胎发育、孵化和死亡情况。挑取280头健康活跃的1龄若虫,接种到余甘子鲜嫩枝叶上,将寄主枝条置于垫有湿润纱布直径15 cm的培养皿,寄主枝条基部用湿润纱布包裹以保鲜。将培养皿置于相同条件的HWS培养箱内,每5 d喷水保湿,每7 d更换新鲜寄主,每2 d置于体视显微镜下观察各虫态体征变化、发育历期、雌雄分化、交尾产卵。雌雄分化后设置两组新的培养皿和寄主,其中一组接种健康3龄雌若虫和雄蛹各10头并标记为两性生殖组,另一组则接种10头健康3龄雌若虫并标记为孤雌生殖组,每组设置6次重复;待发育至雌成虫阶段时,分别观察、记录两组生殖环境中的雌成虫是否产卵以此判断其生殖方式。整个室内培养试验观察过程中,采用OLYMPUS Cellsens Standard显微成像系统分别对各龄期、各虫态60个样本进行体长、体宽测量和拍照,记录发育历期。
1.3.2仿生饲养:采用蒋华等(2021a)介壳虫迁地生活史观测研究方法,于2019年12月将丛岗村虫源地内的越冬受精锡兰玻壳蚧雌成虫寄生枝条嫁接至砧木,成活后移栽至自然环境相同、方便观察的试验地进行栽培管护,设置120目/cm2养虫网(100 cm×100 cm×150 cm)。2020年1-12月定期进行观察,记录各虫态发生期,并结合室内培养和野外调查,描述生活习性,绘制生活史。
1.4 数据分析
采用SPSS 21.0(IBM,美国纽约州阿蒙克市)统计软件对数据进行整理和分析,应用GraphPad Presm 8.0软件绘制统计图,应用Adobe Illustrator CS6软件进行图片编辑。
2 结果
2.1 锡兰玻壳蚧的发生为害及空间分布型
调查结果显示,锡兰玻壳蚧在云南保山调查的余甘子栽培林中的为害率为32%~56%,虫口密度为10.95~94.26头/株,二者具有显著相关性(R2=0.94)(图1)。对发生为害的3块余甘子林雌虫种群的空间分布型分析结果表明,聚集度指标C>1,I>0,Ca>0,m*>1,m*/m>1,并且0
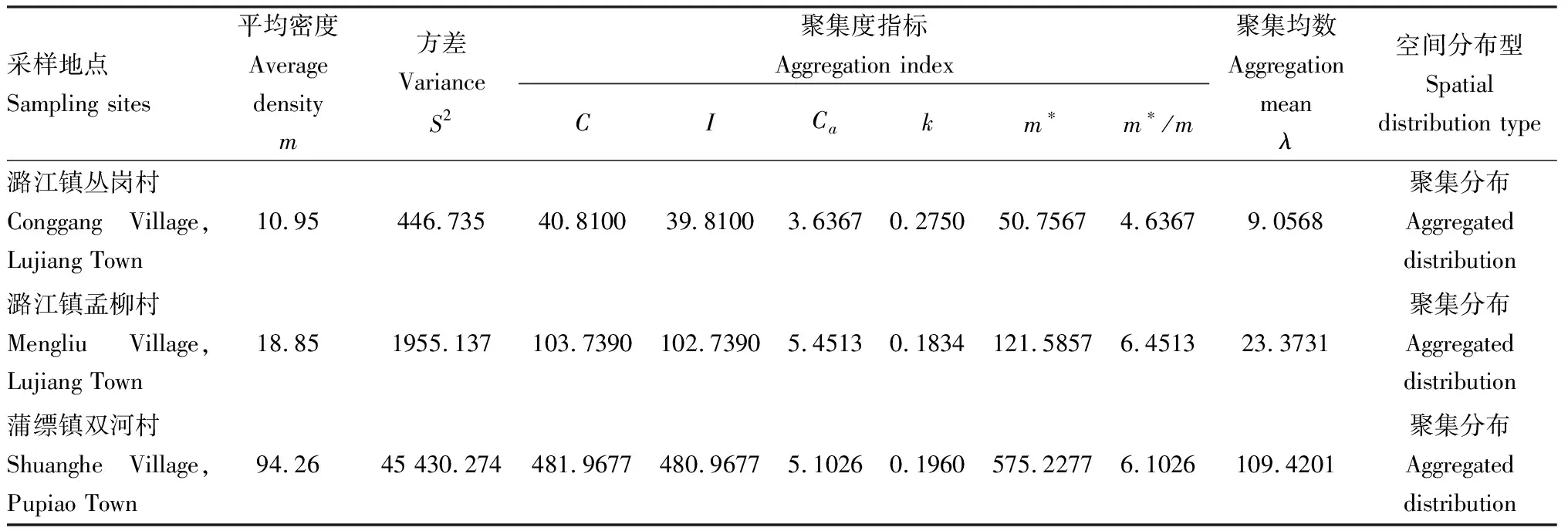
表3 锡兰玻壳蚧雌成虫的空间分布型(云南保山, 2019)Table 3 Spatial distribution pattern of female adults of Drepanococcus chiton (Baoshan, Yunnan, 2019)
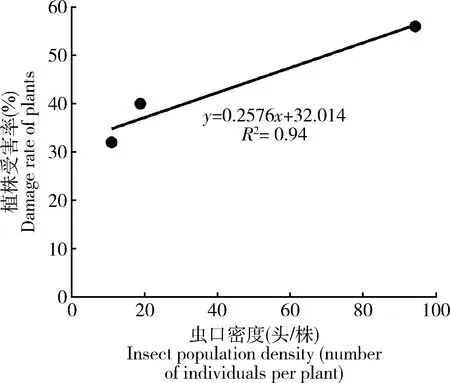
图1 锡兰玻壳蚧种群密度与植株受害率的关系(云南保山, 2019)Fig. 1 Relationship between the population density ofDrepanococcus chiton and the damage rateof plants (Baoshan, Yunnan, 2019)
2.2 锡兰玻壳蚧的形态特征
2.2.1卵:长椭圆形,长0.34±0.01 mm,宽0.17±0.01 mm,初产卵为橘黄色,后期灰黄色,卵壳与胚胎分层明显,透过卵壳可见虫体及黄褐色的单眼(图2: A; 图7)。
2.2.21龄若虫:体表光滑,橘黄色半透明,体型扁平或卵形,体长0.41±0.02 mm,体宽0.20±0.02 mm,体缘毛稀疏可见,头、胸、腹、足等部位区分明显,触角发达共5节,单眼球形棕褐色,口器细长,臀裂明显,臀板黄褐色,两侧各有1根细蜡丝,蜕皮前体表逐渐分泌稀疏白色蜡泌物(图2: B, C)。
2.2.32龄若虫:体背稍向上隆起,体色加深呈黄褐色,体长1.38±0.37 mm,体宽0.98±0.34 mm,触角5~6节,臀裂加深,体表分泌出齿状蜡质突起逐渐形成灰白色薄蜡层(图2: D, E)。发育后期分化雌雄,体型宽厚的为雌性3龄若虫,体型圆窄椭圆状的为雄性预蛹。

图2 锡兰玻壳蚧的卵和1-2龄若虫形态特征Fig. 2 Morphological characteristics of eggs and the 1st-2nd instar nymphs of Drepanococcus chitonA: 卵Eggs; B, C: 1龄若虫1st instar nymphs; D, E: 2龄若虫2nd instar nymphs.
2.2.43龄雌若虫:体背大幅隆起,具透明薄蜡壳,体色变为黄绿色或棕黄色,体长2.57±0.30 mm,体宽1.64±0.20 mm。触角9节,臀裂加深明显,体表蜡壳增厚变硬逐渐覆盖虫体背部(图3: A, B)。
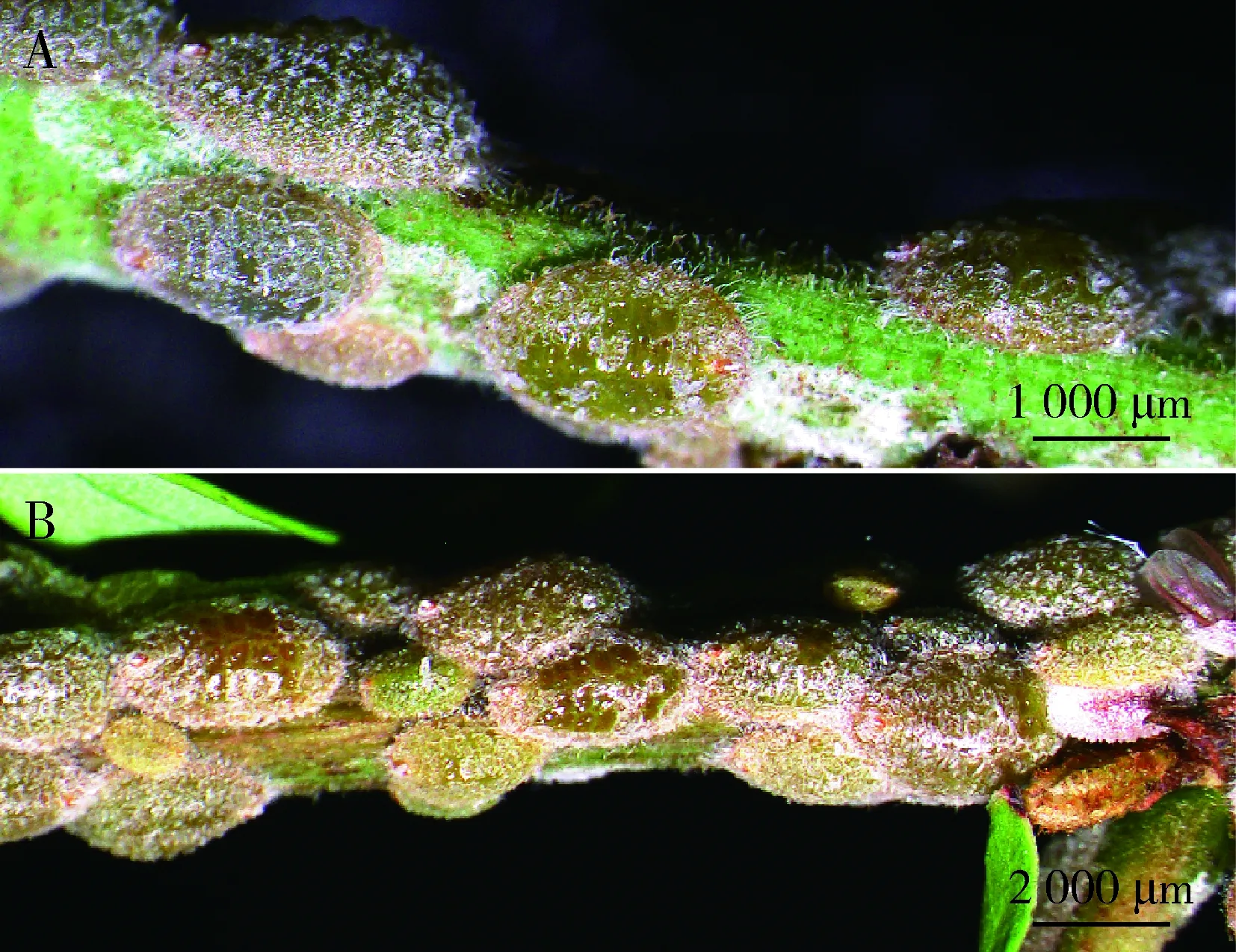
图3 锡兰玻壳蚧3龄雌若虫形态特征Fig. 3 Morphological characteristics of the 3rd instarfemale nymphs of Drepanococcus chitonA: 初蜕皮的3龄雌若虫Newly molted 3rd instar female nymphs; B: 近成熟的3龄雌若虫Nearly mature 3rd instar female nymphs.
2.2.5雌成虫:蜡壳玻璃状银白色,壳长4.27±0.33 mm,壳宽2.41±0.47 mm;产卵前虫体棕褐色,体长3.87±0.45 mm,体宽2.35±0.62 mm,体背高度隆起,体缘扁平具2对锥形体缘刺,臀裂加深内缘凹陷;触角9节,柄节粗壮、梗节和鞭节很小,端部着生刺毛;单眼很小或不可见。产卵前虫体色变为黑褐色,腹末端贴着蜡壳内壁向背前部压缩,直至腹面与前胸面呈70°~90°,体长2.51±0.18 mm,体宽2.33±0.23 mm(图4: A~F)。
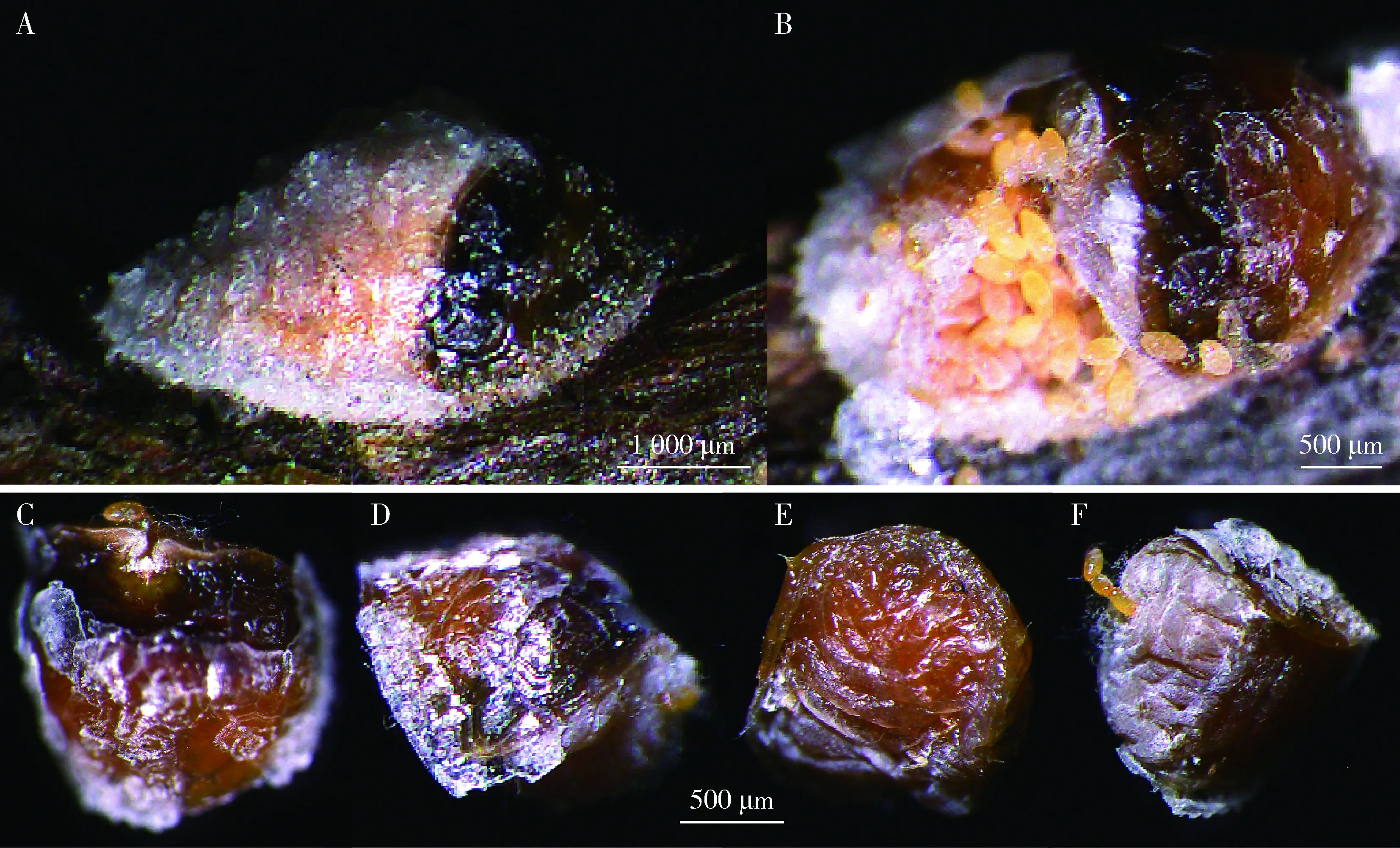
图4 锡兰玻壳蚧雌成虫特征Fig. 4 Characteristics of female adults of Drepanococcus chitonA, B: 雌成虫外观及产卵状Appearance and oviposition of female adults; C, D: 虫体背面和侧面观Dorsal and lateral views of the insect; E: 头、胸部腹面观Ventral view of head and thorax; F: 腹部Abdomen.
2.2.6预蛹:体被透明蜡壳,壳长1.80±0.13 mm,壳宽0.81±0.08 mm(图5: A)。虫体表光滑,黄褐色,长椭圆形,体长1.76±0.04 mm,体宽0.74±0.03 mm,头部圆润,单眼逐步蜕化或隐约可见,触角、足紧缩,臀背板扁平,臀裂明显相互靠拢(图5: B, C)。

图5 锡兰玻壳蚧雄蛹形态特征Fig. 5 Morphological characteristics of male pupae of Drepanococcus chitonA: 预蛹蜡壳Wax shell of prepupa; B, C: 预蛹虫体背面和腹面观Dorsal and ventral views of prepupae; D: 蛹蜡壳Wax shell of pupa; E, F: 蛹体背面和腹面观Dorsal and ventral views of pupa.
2.2.7雄蛹:体被透明银白色蜡壳,蛹体赤褐色,体长1.59±0.03 mm,体宽0.69±0.03 mm。触角透明发达,沿着体缘两侧向腹末端分布;足基节至股节伸至体侧缘,前足胫节至跗节弯曲呈弧形伸向头部,中足和后足胫节至跗节沿着体侧缘伸向腹末端。头部略呈扁圆形,头顶左右两侧具4对黑色单眼,中间2对大,顶部和基部2对小。前胸背板发达,翅芽宽舌状光滑透明,紧贴腹背两侧伸至腹中部。腹部发达与前胸同宽,分节不明显,外生殖突发达从腹部末端伸出,呈尖锥状(图5: D~F)。
2.2.8雄成虫:体长1.43±0.04 mm,体宽0.49±0.03 mm,翅展3.23±0.06 mm。头淡褐色至黑褐色,胸腹部赤褐色。触角丝状,共10节,柄节粗短着生头顶两侧,梗节卵形端部着生数根小刺毛,鞭节细长着生稀疏长刺毛。单眼4对,黑色发达。胸部发达,宽过头部,前胸背板硬化基部具1月牙状突起。足细长,棕褐色,各节均着生稀疏长刺毛。前翅发达近于透明,浅褐色,翅脉2条发达棕褐色,后翅退化。腹部8节,腹末端中部伸出1根刺状外生殖突,两侧具2根白色蜡丝(图6: A~C)。各虫态形态体征见表4。
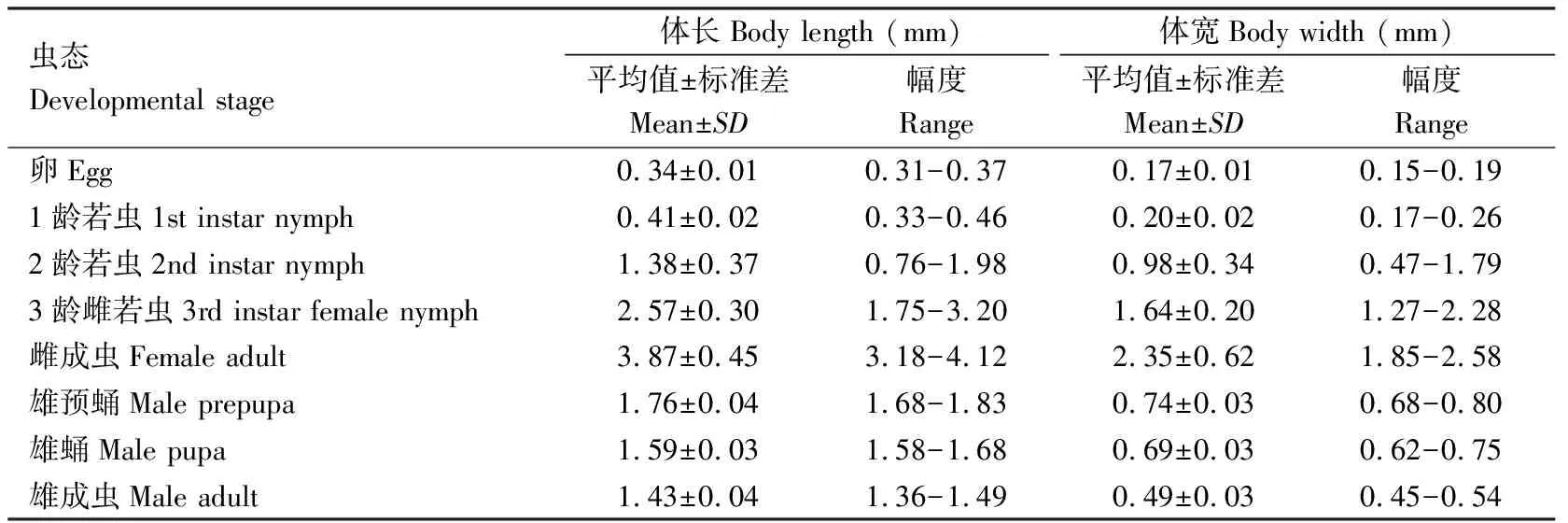
表4 锡兰玻壳蚧各虫态体长和体宽Table 4 Body length and width of various developmental stages of Drepanococcus chiton
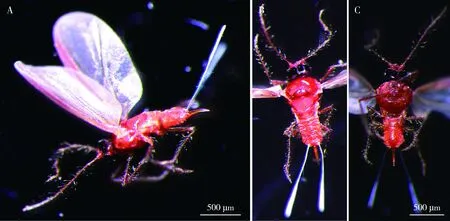
图6 锡兰玻壳蚧雄成虫形态特征Fig. 6 Morphological characteristics of male adults of Drepanococcus chitonA: 侧面观Lateral view; B: 背面观Dorsal view; C: 腹面观Ventral view.
2.3 锡兰玻壳蚧的生活史
试验地饲养观察结果表明,锡兰玻壳蚧在我国滇西热带干热河谷地区余甘子果园内1年发生2代,以受精雌成虫在寄主枝条上越冬,越冬期卵和低龄若虫自然死亡现象突出。翌年1月上旬-2月中上旬越冬雌成虫产卵,第1代1龄若虫于1月中旬开始活动,1月下旬-2月中旬为1龄若虫分散活动和2龄若虫固定刺吸为害高峰期。雌雄分化始于2月中旬,雌性3龄若虫持续固定刺吸发育,雄性进入预蛹,3月中旬达到化蛹高峰,3月中下旬为雄虫羽化和雌雄交尾高峰期。4月上旬-5月中旬为受精雌成虫个体发育和孕卵期,5月下旬开始产卵,产卵高峰期在6月下旬-7月中旬。第2代1龄若虫于6月上旬开始活动,6月下旬-7月中旬为1-2龄若虫活动高峰期,6月下旬化蛹,雄成虫羽化始于7月中上旬,7月下旬-8月下旬为雄虫羽化和雌雄交尾高峰期。交尾高峰前的受精雌成虫于10月上旬产卵,11月上旬低龄若虫逐步死亡。交尾高峰期后的受精雌成虫于10月下旬进入越冬(表5)。
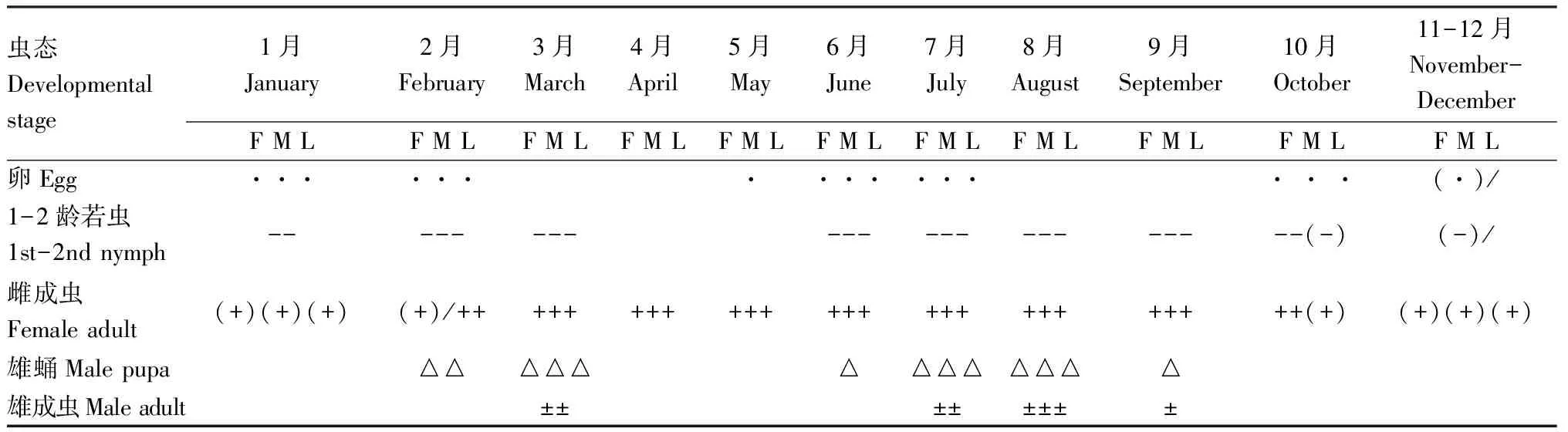
表5 锡兰玻壳蚧年生活史(云南保山, 2020)Table 5 Life history of Drepanococcus chiton(Baoshan, Yunnan, 2020)
2.4 锡兰玻壳蚧的产卵能力、生殖方式及胚后发育类型
室内培养观察发现,锡兰玻壳蚧雌成虫单雌产卵量为847.03±13.72粒,卵孵化率为97.59%;卵单产堆积于母体腹外被蜡壳保护,靠卵壳吸收营养孕育胚胎形成若虫破壳而出,属于典型的卵生发育类型(图7)。室内培养种群经个体发育分化为雄性预蛹106头、雌性3龄若虫136头,雌雄性比为1.28∶1,两性分化特征明显;进一步观察两性生殖环境下雌虫均产卵,而孤雌生殖环境下雌虫均未产卵,故锡兰玻壳蚧生殖方式为两性生殖(图8)。

图7 锡兰玻壳蚧的胚后发育类型Fig. 7 Types of postembryonic development ofDrepanococcus chiton
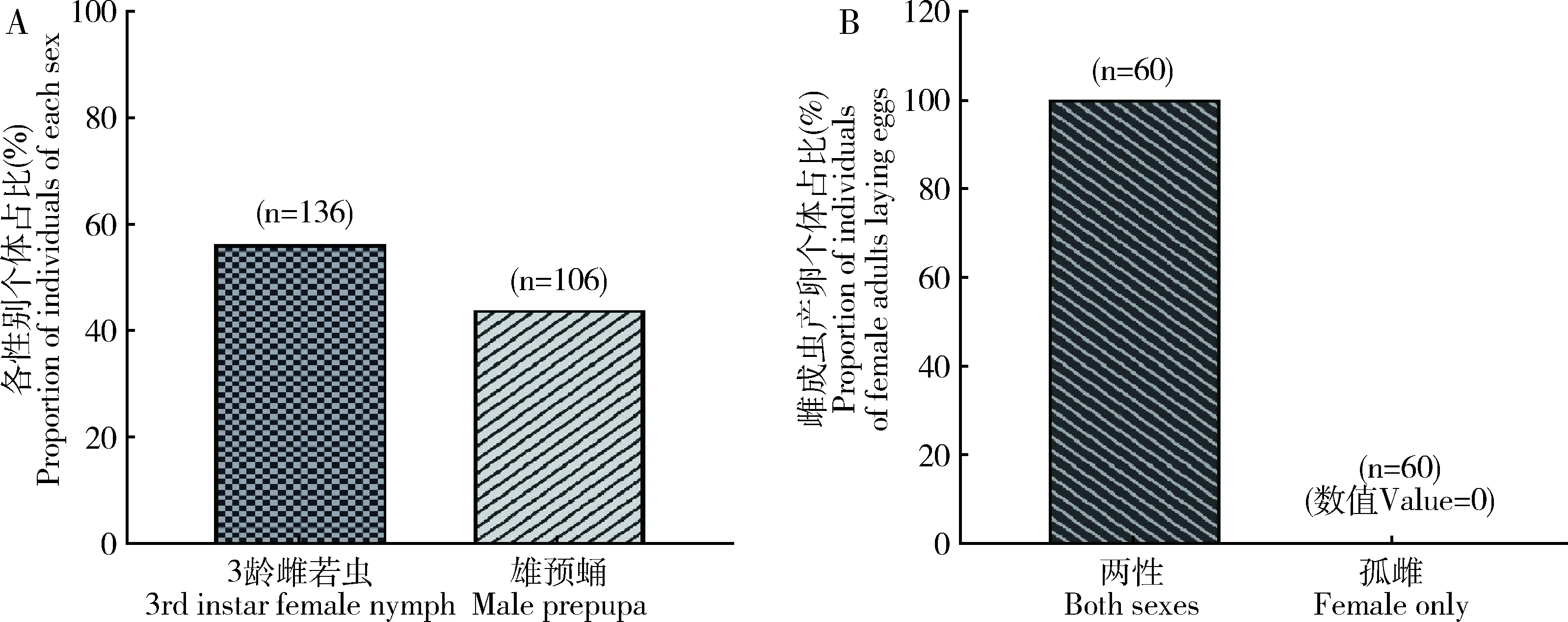
图8 锡兰玻壳蚧的生殖特性Fig. 8 Reproductive characteristics of Drepanococcus chiton
2.5 锡兰玻壳蚧的发育历期和习性
室内培养和仿生饲养种群观察结果表明,锡兰玻壳蚧雌成虫产卵期长达8.34±1.16 d,卵单个产于母体腹末肛门处,盛期平均每8~10 min产卵1粒,后期产卵量明显下降。卵堆积在母体蜡壳内或挤压顶破蜡壳散落,平均历期为9.60±1.41 d;1龄若虫剧烈蠕动顶破卵壳滞留15~40 min后,开始四处爬行或随风扩散寻找庇护场所,平均历期为13.55±1.82 d;2龄若虫在寄主枝条固定刺吸发育(受惊时会爬行另寻新的庇护场所),7-10 d体征发生变化完成雌雄分化,平均历期7.35±0.80 d;3龄雌若虫永久固定于枝条刺吸发育,13-18 d完成交尾,平均历期13.80±1.52 d;受精雌成虫靠吸收寄主营养完成个体发育和孕卵,35-45 d开始产卵,产卵结束后5-7 d死亡,平均历期56.65±3.26 d;雄性预蛹5-7 d化为完全蛹态,平均预蛹期5.85±0.86 d;雄蛹7-10 d羽化为成虫,平均蛹期8.05±1.13 d;雄成虫羽化后10-24 h内寻找配偶完成交尾,交尾时长30~50 min,可多次寻找不同雌虫个体进行交尾,交尾后1 d内死亡,平均历期为1.77±0.72 d(表6)。

表6 锡兰玻壳蚧室内培养种群各虫态发育历期(云南保山, 2020)Table 6 Duration of various developmental stagesof the indoor cultured population of Drepanococcuschiton (Baoshan, Yunnan, 2020)
3 讨论与结论
蜡蚧科最常见的生殖方式是两性生殖,有时也存在混合产雌单性生殖(王子清, 2001)。本研究表明,锡兰玻壳蚧只营两性生殖,孤雌生殖无法产生后代,与前人在马来西亚寄主杨桃上的观察结果(Ibrahim, 1994)一致。大量研究表明不同地理纬度和环境温度下的昆虫种群发育历期存在差异,低纬度种群发育历期常短于高纬度种群,高温度栖息种群发育历期常高于低温度栖息种群(涂小云和陈元生, 2012; 潘飞, 2014)。例如亚洲玉米螟Ostriniafurnacalis在20~31℃栖息纬度下,卵期、蛹期与栖息纬度存在正相关关系(涂小云等, 2015);花斑花
绒寄甲Dastarcushelophoroides各虫态发育历期随栖息纬度的升高而延长(陈元生等, 2018);扶桑绵粉蚧Phenacoccussolenopsis、石蒜绵粉蚧Phenacoccussolani和康氏粉蚧Pseudococcuscomstocki等蚧虫种群发育历期均随栖息环境的温度升高而缩短(徐盼等, 2012; 王飞飞等, 2014; 李思怡等, 2018)。本研究表明锡兰玻壳蚧在滇西保山均温18.4~21.6℃栖息环境中1年发生2代,种群世代历期为83.15±4.95 d(表6),明显长于该虫在马来西亚乌鲁冷岳杨桃果园内的世代历期49.9±1.06 d(Ibrahim, 1994);两地栖息种群除寄主植物不同外,滇西保山的种群栖息年均温低于乌鲁冷岳近10℃,地理纬度高出乌鲁冷岳及该虫发源地斯里兰卡近20°。可见研究结果与相关文献报道基本吻合,锡兰玻壳蚧种群在高纬度、低温度地区世代发育历期有所延长,但该虫种群发育有效积温和种群消长演替等还需进一步深入研究。
目前世界上记载玻壳蚧属仅有4种,锡兰玻壳蚧是我国近年来新记录的一种,仅在广州南沙区白楸植物上采集过该虫体样本(仇玲, 2020),本研究发现的寄主余甘子为国内新记述,入侵来源、发生历史及天敌种类尚未查清。在国外该虫寄主范围很广,尤其喜食热带水果植物,被我国进出口检疫列为输华水果植物高风险的有害生物之一(焦懿等, 2010)。然而,我国滇西正处热带、亚热带水果植物适生栽培的主产区,笔者通过对余甘子及周边热带农业水果产区调查发现,锡兰玻壳蚧在余甘子林内的种群密度、产卵繁殖量远高于已报道的堆蜡粉蚧Nipaecoccusvastator和埃及吹绵蚧Iceryaaegyptiaca(蒋华等, 2021b),发生严重的14.66 hm2余甘子果园年扩散范围达17.28%;毗邻的石榴Punicagranatum果园已有少量植株枝条被寄生,周边的芒果Mangiferaindica、荔枝Litchichinensis、龙眼Dimocarpuslongan和柑橘Citrusreticulata等果园虽尚未发现危害,但威胁性很大,需引起农林部门重视,避免远距离、多寄主间传播扩散。下一步我们还将对该害虫的防治措施,特别是理化诱控、天敌利用、生物制剂施用等措施开展深入研究。
本研究首次报道了锡兰玻壳蚧在我国滇西地区不同虫态的形态特征及生物学特性,其非常适应该地区热带、亚热带干热河谷的气候环境,该虫已定殖为害余甘子植株,是影响余甘子开发利用及产业振兴的重要害虫种类,生产上应充分结合其相关生物学特性开展科学防控。加强天敌利用,合理间伐密植,剪除烧毁虫枝,严格检疫除害,科学监测预报,抓准1-2龄若虫分散薄弱期精准施药,多措并举,未雨绸缪,防患于未然。
猜你喜欢
杂志排行

昆虫学报的其它文章
- Morphological differentiation in the Asian honey bees (Apis cerana) in China
- 贝氏小奥林螨连续饲养的草栖钝绥螨生命表参数和捕食能力
- 施药后不同时间玉米田中常用农药残留对玉米螟赤眼蜂的影响
- Inhibition of glucose oxidase gene decreases the resistance of Mythimna separata (Lepidoptera: Noctuidae) larvae to Bacillus thuringiensis infection
- 枣食芽象甲化学感受蛋白PyasCSP4的基因克隆、表达及配体结合特征
- RNAi介导的GdHsp60和GdHsp70基因沉默对沙葱萤叶甲幼虫抗寒性的影响