朗德鹅TLR4基因的生物信息学分析
2022-08-09王权锜张会新刘玉芬
王权锜,张会新,程 敏,刘玉芬
(哈尔滨师范大学 生命科学与技术学院,黑龙江 哈尔滨 150025)
Toll样受体(TLRs)属于一种模式识别受体(PRPs),在动物免疫应答反应中起重要作用,其通过识别病原相关分子模式(PAMPs)来启动天然免疫应答,达到清除病原微生物及相关代谢产物的目的。目前已经在禽类中发现10种TLRs(TLR1La/b、TLR2a/b、TLR3/4/5/7/15/21),其中TLR3/4/5/7与哺乳动物中发现的TLRs是直系同源物[1-2]。而TLR4作为唯一一种介导MyD88和TRIF两种信号通路的TLRs家族成员,可以引发一系列级联反应导致TNFα、IL-6、IL-8和IL-12等细胞因子的释放[3-4]。Wang等[5]首次克隆出朗德鹅TLR4基因,全长2641 bp,预测编码843个氨基酸,并检测出在过度喂养条件下,TLR4在皮下脂肪和肠中的表达显著增加,但在心脏中表达下降。朱道仙等[6]研究证实患痛风病的雏鹅肠道内菌群发生变化可激活TLR4/MyD88/NF-κB通路,进而诱导炎症反应使肾脏受损。上述研究提示TLR4在鹅免疫系统中发挥着重要作用。本研究通过生物信息学的方法分析鹅TLR4蛋白特征,为进一步探索TLR4基因在鹅免疫系统中的功能提供理论基础。
1 材料与方法
1.1 试验材料
从NCBI中检索不同物种TLR4基因编码蛋白质序列,朗德鹅(AEC32857.1)、马岗鹅(AKF02412.1)、绿头鸭(NP_001297342.1)、芦花鸡(AFK08533.1)、灰山鹑(AGO86772.1)、黑尾原鸡(ACR26280.1)、非洲鸵鸟(AKP92940.1)、斑胸草雀(ACN58233.1)、密河鳄(KYO23184.1)、虎斑响尾蛇(XP_039178821.1)、斑马鱼(XP_009292463.1)、鲤鱼(XP_018980331.2)。
1.2 试验方法
通过生物信息学软件对朗德鹅TLR4基因编码产物理化性质、结构、功能以及进化性进行分析(表1)。

表1 生物信息学软件及网址
2 结果
2.1 鹅TLR4基本特征
通过ExPASy-ProtParam、ExPASy-ProtScale以及UniProt数据库对朗德鹅TLR4基本理化性质、亲/疏水性以及亚细胞定位进行预测,结果显示TLR4基因共编码843个氨基酸残基,分子式为C4364H6867N1135O1239S32,理论等电点7.55,亲水系数0.019(>0)。疏水性最强残基为亮氨酸(3.611),亲水性最强氨基酸残基为组氨酸(-3.311),疏水性氨基酸残基相对较多(图1)。UniProt数据库预测可以看出鹅TLR4大部分定位于细胞膜(图2)。
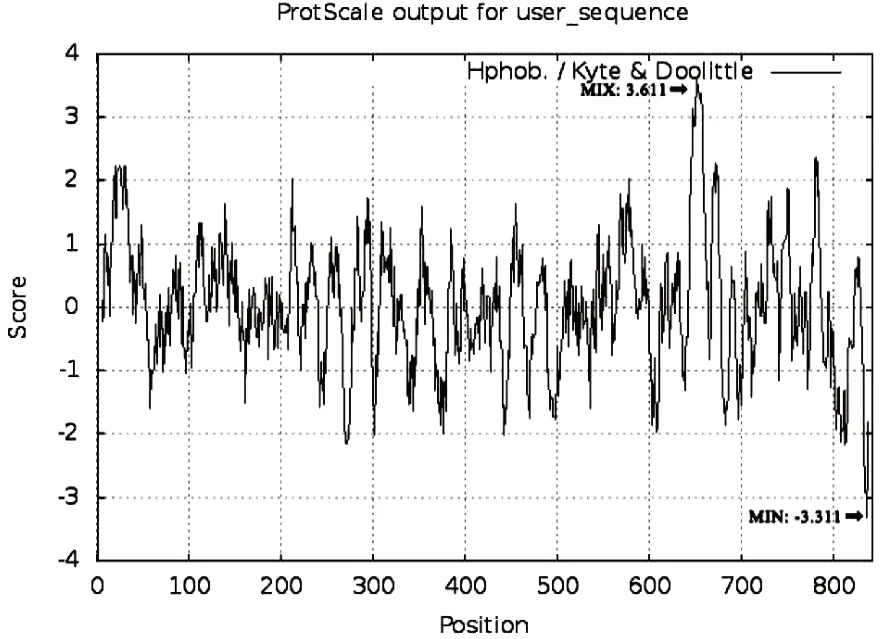
图1 鹅TLR4亲/疏水性预测

图2 鹅TLR4亚细胞定位
2.2 鹅TLR4基础结构预测
通过SignalP-5.0、TMHMM-2.0以及SMART对朗德鹅TLR4进行信号肽、跨膜结构域以及结构域预测,结果显示鹅TLR4具有信号肽的可能性为0.8497,剪切位点位于第28和29位残基之间(图3),与SMART预测结果相似。TMHMM-2.0预测出2个跨膜结构域(图4),第一个跨膜结构域位于第12至第34位残基之间;第二个跨膜结构域位于第640至662位残基之间。SMART预测结果显示,TLR4胞外区具有8个亮氨酸重复序列(Leucine-rich repeat,LRR)、1个C端亮氨酸富集区(Leucine-rich repeat C-terminal damain,LRR_CT)以及胞内区的TIR结构域(图5)。
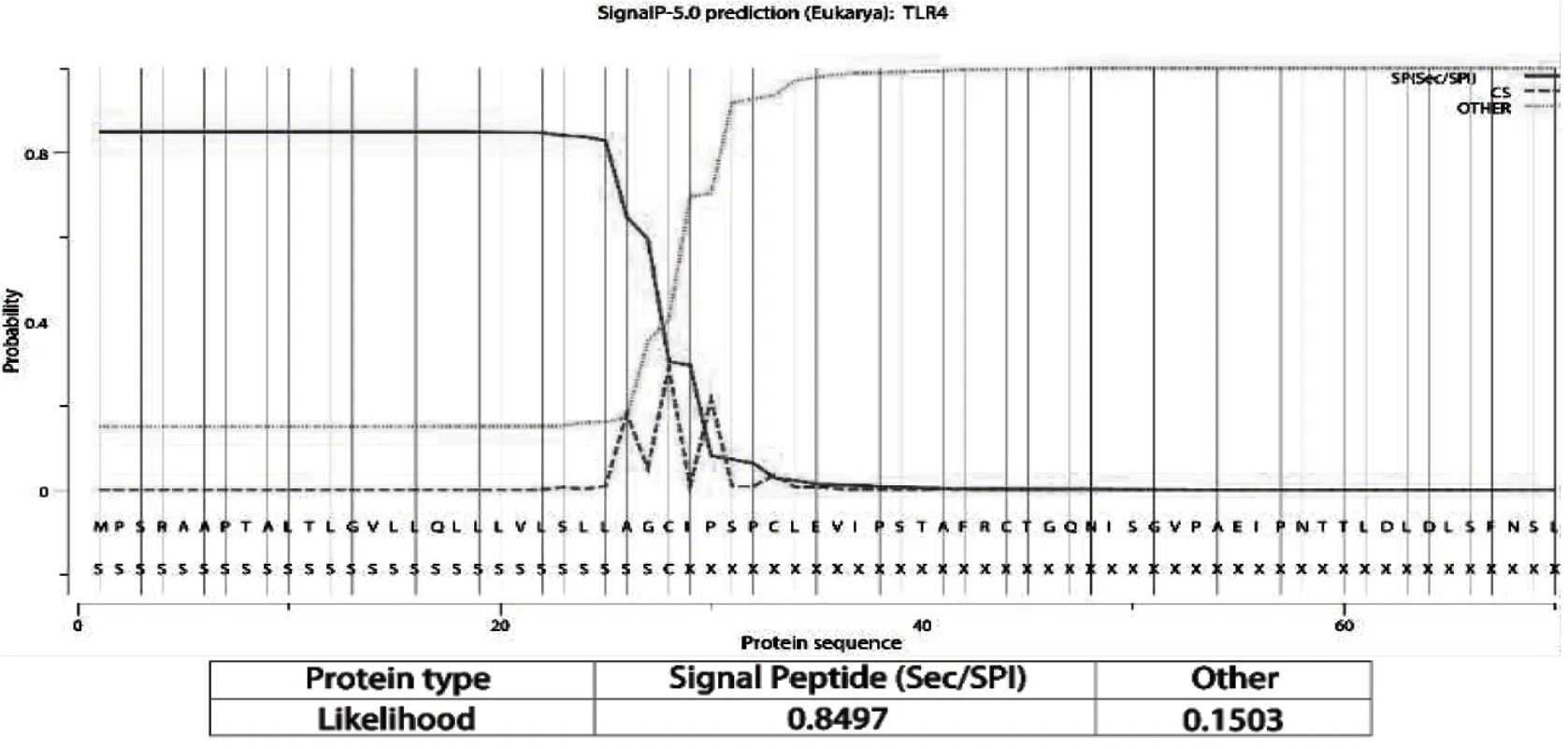
图3 鹅TLR4信号肽预测
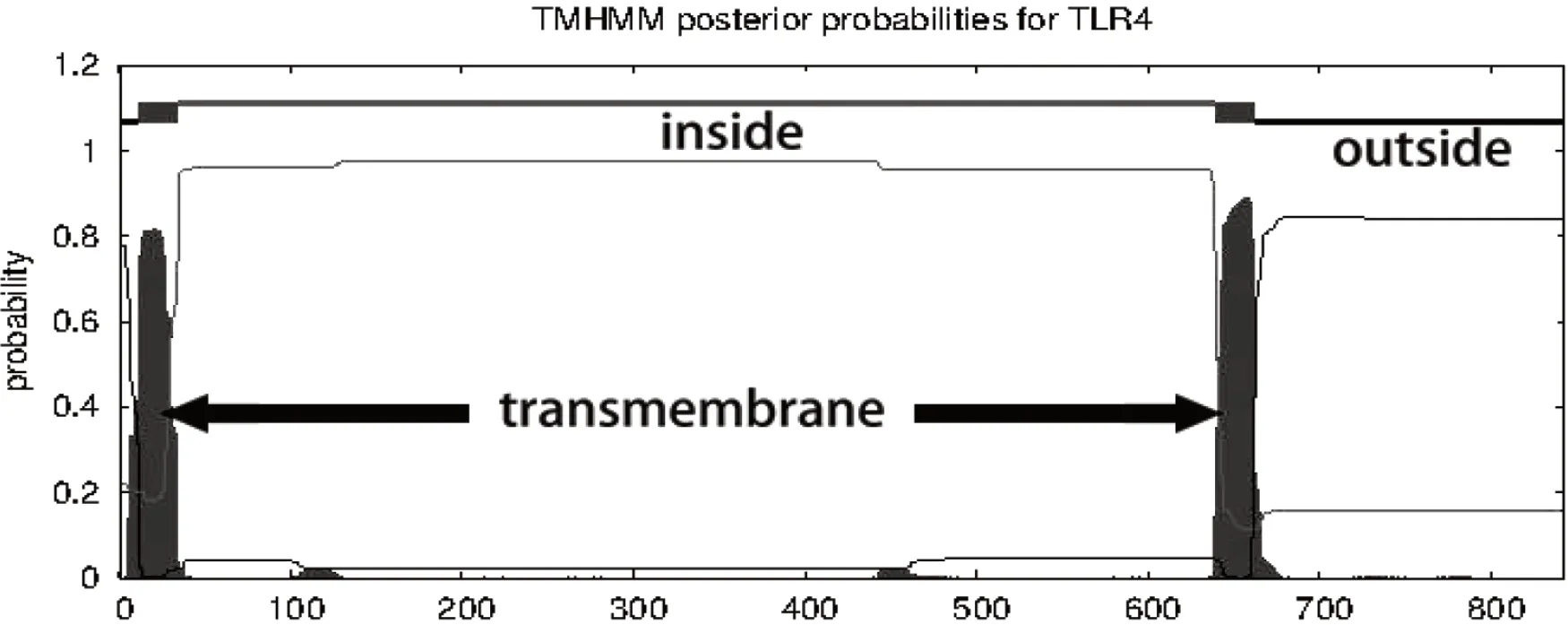
图4 鹅TLR4跨膜预测

图5 鹅TLR4结构域预测
2.3 不同物种间TLR4氨基酸序列的进化性
通过MEGA7分析在NCBI中检索的不同物种TLR4氨基酸序列的进化性。结果显示,朗德鹅与马岗鹅、绿头鸭、芦花鸡、灰山鹑、黑尾原鸡亲缘关系最近,与非洲鸵鸟、斑胸草雀的亲缘关系较近,而与爬行类和鱼类的亲缘关系较远(图6)。
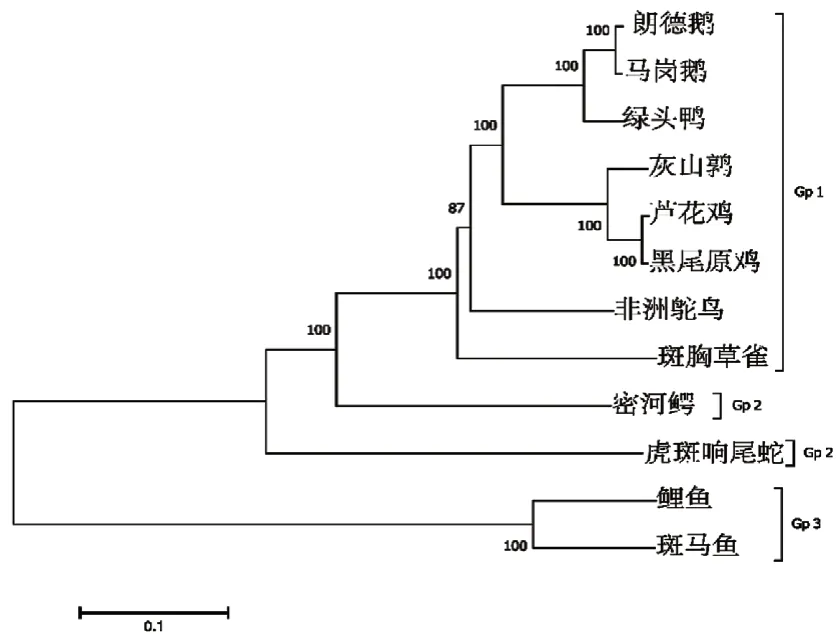
图6 朗德鹅TLR4氨基酸序列进化树
2.4 鹅TLR4修饰位点预测
通过NetNGlyc-1.0和NetPhos-3.1预测鹅TLR4修饰位点。其中N-糖基化位点存在12个,但位于第626和640位残基的修饰可信度得分较低,分别是0.3883和0.3824(图7)。磷酸化位点预测显示鹅TLR4存在89个磷酸化位点,其中丝氨酸(S)61个、苏氨酸(T)22个、酪氨酸(Y)6个。
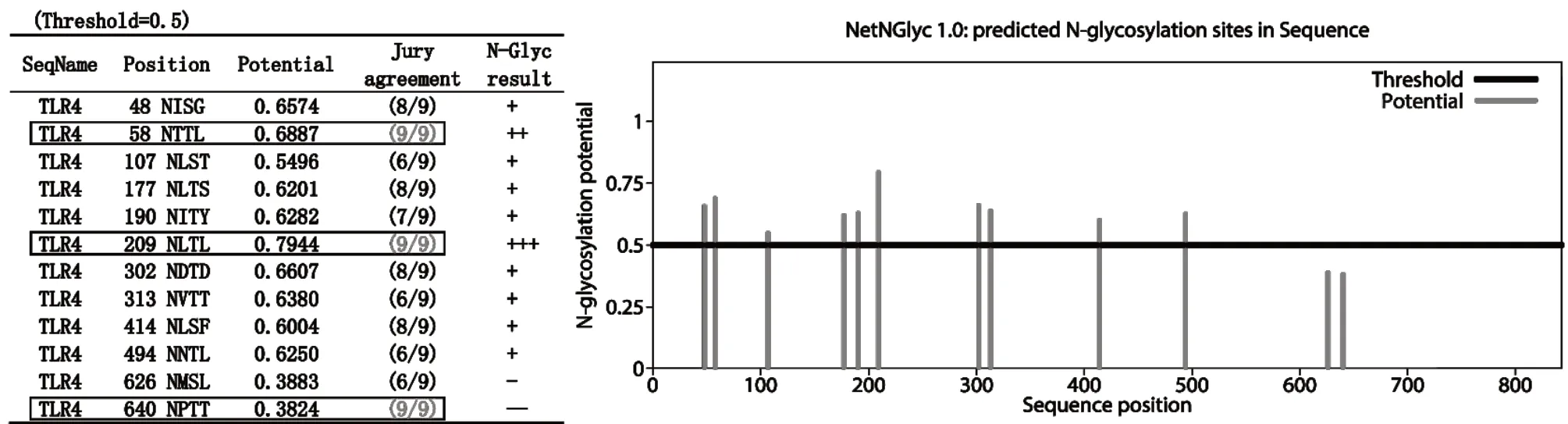
图7 鹅TLR4 N-糖基化位点
2.5 鹅TLR4高级结构预测
通过SOPMA和I-TASSER预测鹅TLR4二级、三级结构,结果显示鹅TLR4蛋白由α-螺旋(54.69%)、延展链(12.34%)、β-转角(2.58%)以及无规则卷曲(30.13%)四种结构组成(图8)。三级结构预测结果可以看出,胞外区是TLR家族典型马蹄形结构,纯黑色部分为亮氨酸(Leu),大部分位于胞外区,与SMART预测的LRR结构域存在于胞外区结果相吻合,结合二级结构预测可判断跨膜方式属于α-螺旋跨膜(图9)。

图8 鹅TLR4二级结构
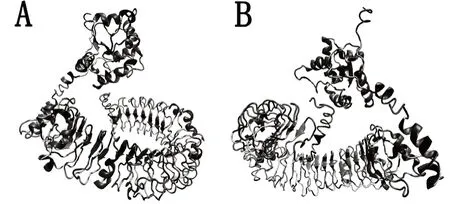
图9 鹅TLR4三级结构
2.6 鹅TLR4相互作用预测
通过STRING数据库预测与鹅TLR4相互作用的蛋白质,并构建相互作用网络图(图10)。结果显示鹅TLR4与TIRAP、LY96、MYD88、HMGB1以及TRAF6等10种蛋白质有相互作用,相关评分见表2。

表2 TLR4与10种蛋白质相关度评分
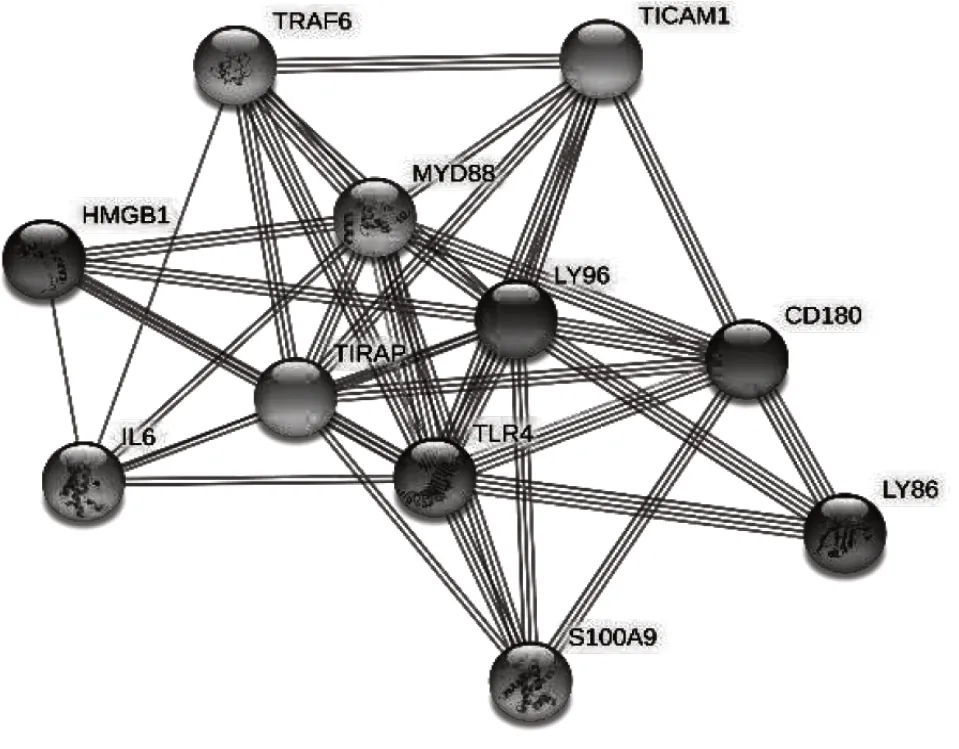
图10 鹅TLR4相互作用预测
3 讨论与结论
在朗德鹅TLR4蛋白基本理化性质分析中可以看出在600~700位残基之间存在疏水峰值,推测该位置可能为疏水的跨膜结构域,之后的跨膜结构域以及结构域预测则证实了上述猜测,这也与猪、牛等哺乳动物跨膜结构域预测相吻合[7-8]。在TMHMM-2.0预测出来的两个跨膜结构域中,第一个跨膜结构域与SignalP-5.0预测的信号肽区域重合,可以判断该位置并没有跨膜结构域,这符合SMART的预测结构。
TLR4胞外区具有保守性的结构域LRR,但在三花鹅TLR4胞外区前321位残基中有6个LRR[9],而朗德鹅相同位置的LRR的数量只有4个,可能是因为106位和205位残基发生错义突变所导致,而在哺乳动物海南地方猪中已经证实G7209T或G7781A位点的突变可导致TLR4蛋白质分子结构发生改变,并可能影响LRR的数量[10]。可以看出TLR4基因序列具有多态性,具体功能是否受到影响还有待进一步探究。但以往研究已经证明在鸭中免疫机能会受到TLR4第3外显子中基因多样性的影响[11]。
TLR4氨基酸序列进化性分析显示,TLR4在不同物种间存在一定差异性,与三黄鸡TLR4进化性分析相符[12]。其中朗德鹅与马岗鹅亲缘关系最近,而与同为禽类的非洲鸵鸟、斑胸草雀亲缘关系较远,可能是因地缘关系所导致,在鱼类中与斑马鱼亲缘关系较远,这可能是斑马鱼和禽类的TLR4相比不仅缺失了识别LPS的能力,同时也缺少了信号肽区[13-14]。String蛋白互作预测结果显示,TIRAP、LY96、MYD88评分达到了0.999,LY96是协同TLR4识别其激动剂LPS的重要蛋白[15],TIRAP是含TIR域的接受子蛋白,其与MYD88、TRIF、TRAM等在TIR下游信号传导中起着重要作用,MYD88是一种胞质衔接蛋白,在实验中是衡量TLR4表达量的重要参照。
通过生物信息学分析朗德鹅TLR4蛋白基本特性、修饰位点、进化性以及高级结构特征等,结构符合TLRs家族典型特征,修饰位点多处于胞外功能区,进化性分析说明不同种生物间TLR4存在着种属差异性。本分析为研究TLR4的免疫功能以及鹅养殖中抗病育种提供了理论基础。
