外源水杨酸对高温胁迫下小麦籽粒灌浆特性的影响
2022-08-09樊永惠李宇星武倩倩秦博雅杨文俊黄正来
樊永惠,葛 婷,杨 咏,李宇星,武倩倩,秦博雅,杨文俊,黄正来
(1.安徽农业大学农学院/农业部黄淮南部小麦生物学与遗传育种重点实验室,安徽合肥 230036;2.太湖县江塘乡农业综合服务中心,安徽太湖 246400)
小麦作为世界三大作物之一,在全球栽培总面积最大,分布范围最广,总产量最高,对保障粮食安全起到重要作用。全球气候变暖使得小麦频繁遭受高温损害,特别是灌浆期高温胁迫严重影响到小麦产量和品质。研究表明,相较籽粒灌浆的适宜温度(即18.0~22.0 ℃)每升高1 ℃,小麦减产5%~10%,当小麦生育后期最高温度超过30 ℃,损失更大甚至绝收。
水杨酸(SA)是一种普遍存在于高等植物细胞内的小分子酚类物质,其在诸多植物内发挥提升生物及非生物胁迫耐受性能的作用。前人研究大多集中在SA在抗寒、抗盐碱以及抗病等方面的作用机理,对于SA抗热的研究较少。粒重是决定小麦产量的重要因素之一。周秋峰等研究发现,高温胁迫造成小麦的过氧化物酶(POD)和超氧化物歧化酶(SOD)活性降低,小麦光合能力降低、植株衰老加速,供给籽粒合成的光合产物降低,导致小麦粒重和粒数减少,产量降低。而向妙莲等研究发现,外源水杨酸可提高小麦超氧化物歧化酶(SOD)活性,减缓叶片脂质过氧化作用,延缓叶片衰老,利于灌桨速率和粒重的提高。高温胁迫下,外施SA可以提高小麦植株的耐热性,并显著影响植株内源激素的变化。内源激素是植物体内产生的一类在低浓度即可对植物生长发育及环境应答等过程产生重要调控作用的代谢产物。薛建平等研究发现,在高温胁迫下,内源激素ABA、IAA和GA3对半夏倒苗起着重要的调控作用,外源喷施SA可延缓高温胁迫下半夏的倒苗,提高产量。但外源SA对高温胁迫下小麦内源激素的影响迄今未见报道,且有关SA缓解小麦高温胁迫的生理机制以及SA对灌浆期高温胁迫下小麦籽粒灌浆特性的影响尚未完全明确。本研究选择在抽穗期、开花期、花后5 d、花后10 d四个不同时期喷施SA(每个时期连续喷施4 d),在花后15-19 d通过被动式增温棚模拟高温环境,分析不同处理对产量、籽粒灌浆特性的效应,以期为小麦耐高温机理和抗高温栽培技术提供理论依据。
1 材料与方法
1.1 试验设计
本试验于2018-2020年在安徽农业大学校内试验基地农萃园(31.83°N、117.24°E)进行。土壤类型为黄棕壤土,2018-2019年度小麦播种前0~20 cm土层土壤pH=6.5,土壤有机质 13.9 g·kg、速效氮86.2 mg·kg、速效磷13.4 mg·kg、速效钾71.5 mg·kg。 2019-2020年度小麦播种前0~20 cm土层有机质15.2 g·kg、速效氮90.6 mg·kg、速效磷16.5 mg·kg、速效钾79.6 mg·kg。供试小麦品种为扬麦18(本地区大面积推广品种)。采取大田盆栽方式,盆内直径25 cm,深度为30 cm,每盆装7.5 kg筛土。种植时每盆施用复合肥6 g,复合肥中各养分的比例为N∶P∶K= 17∶17∶17。其中,氮肥分两次施入,基追比为 5∶5,追肥于拔节期施入。播种日期为2018年11月7日和2019年11月4日。土壤经水沉实后播种,每盆播种20粒,在三叶一心时,间苗至每盆8株。每个处理21盆,每个处理均设置三个重复。试验盆周围大田种植相同品种小麦,按照高产田块要求进行田间管理。
水杨酸处理浓度为2016-2017年度预试验中筛选出的喷施浓度0.1 mmol·L,在抽穗期(SAHG)、开花期(SAAG)、花后5 d(SA5G)、花后10 d(SA10G)分别对小麦进行叶面喷雾,每个时期连续喷施4 d,每天1次,每次喷施1 L·m。喷施至叶片正反两面均湿润为止(有一层小水珠欲落)。对照组(NN)喷施等量的蒸馏水。采用随机区组试验设计,所有喷施溶液均用1 mmol·LKOH调整pH值为6.0 ± 0.1,每100 mL溶液含有0.02 mL Tween-20。
于花后15 d通过被动式增温棚进行高温处理,连续处理5 d。为防止棚内增温幅度过高,每天正午将塑料薄膜上卷30 cm便于棚内通风。用Elitech(RC-4)智能温湿度记录仪每10 min测1次,连续记录小麦冠层空气温、湿度。处理结束后将盆移至自然环境下。2019年、2020年灌浆期高温胁迫分别平均增温5.7 ℃、6.0 ℃(图1)。
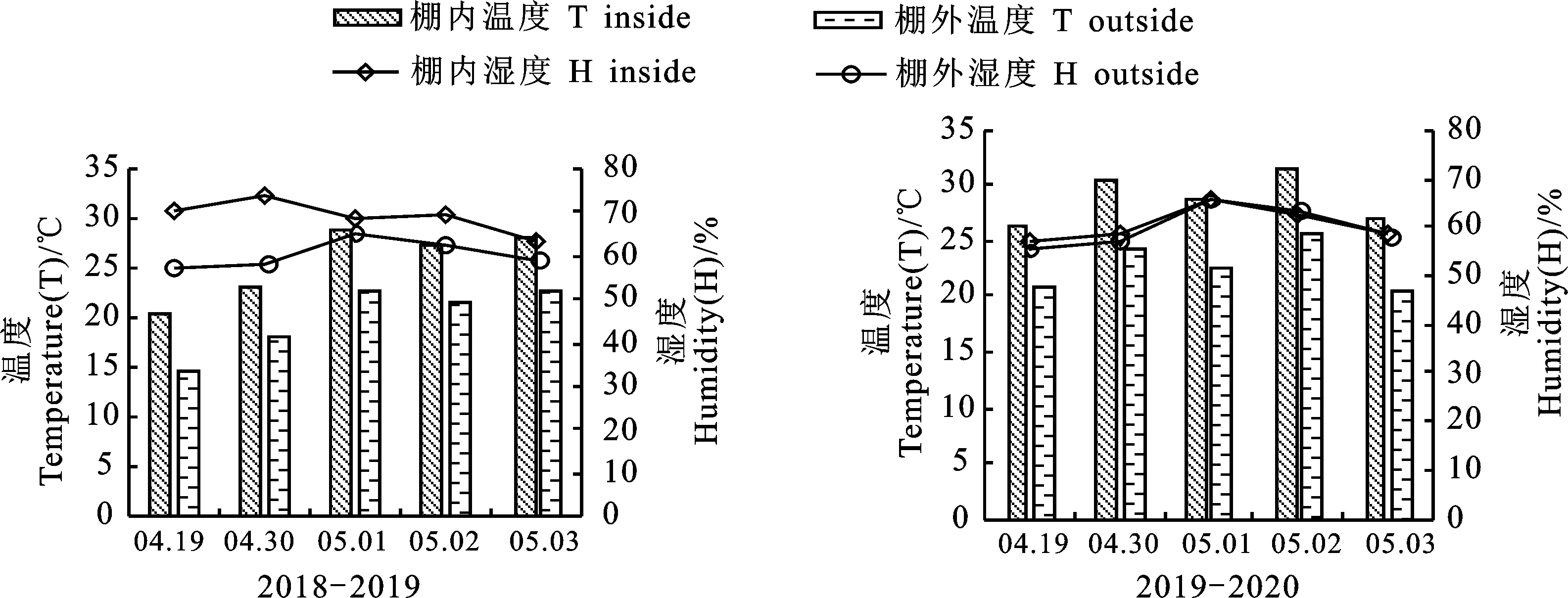
图1 试验地冠层温湿度变化(2018-2020年)
1.2 测定项目与方法
1.2.1 产量测定
成熟期每小区收获6盆,自然风干(小麦籽粒含水率达到13%左右)后测籽粒产量,记录每盆穗数、穗粒数。各处理全部收获、风干后(含水率约13%)测千粒重,三次重复。
1.2.2 强、弱势粒粒重测定
成熟期每处理取10穗参考姜 东等方法将麦穗平均分为三部分,即上、中和下部小穗,每部分小穗基部第1、第2位籽粒计为强势粒,第 3~5位籽粒计为弱势粒。三次重复。
1.2.3 籽粒蔗糖、可溶性糖含量测定
取不同处理花后10、15、20、25、30、35 d籽粒,蔗糖含量采用间苯二酚法测定;可溶性糖含量采用蒽酮比色法测定。
1.2.4 籽粒蔗糖合成酶(SS)活性测定
取花后5、10、15、25 d籽粒,SS活性测定参考Rufty等的方法并略加改进。200 μL SS提取液中加 50 μL pH 7.5的 HEPES-NaOH 缓冲液、50 μL 10 mmol·LMgCl、100 μL 50 mmol·LUDPG、100 μL 50 mmol·L6-磷酸果糖,30 ℃反应30 min;加 200 μL 40% NaOH 溶液混匀,100 ℃保温 10 min;加 2.0 mL30%盐酸混匀,80 ℃保温10 min;加1 mL 1%间苯二酚,混匀后80 ℃保温10 min;冷却后于480 nm下比色测生成磷酸蔗糖含量。用蔗糖生成量表示酶活性。
1.2.5 籽粒内源激素含量测定
采用酶联免疫法(ELISA)略作修改测定籽粒中吲哚乙酸(IAA)、脱落酸(ABA)和细胞分裂素(CTK)含量,具体步骤如下:
(1)样品中激素的提取:取0.5 g花后5、10、15、25 d籽粒,加2 mL提取液[80%(v/v)甲醇,内含1 mmol·L二叔丁基对甲苯酚(BHT)],在冰浴下研磨成匀浆,转入10 mL离心管,再用3 mL提取液分次将研钵冲洗干净,一并转入试管中,摇匀。4℃下过夜提取,之后3 500 r·min离心8 min,取上清液,残渣弃去。
(2)过柱:上清液过C-18固相萃取柱。具体步骤:80%甲醇(1 mL)平衡柱→上样→收集样品→移开样品后用100%甲醇(5 mL)洗柱→100%乙醚(5 mL)洗柱→100%甲醇(5 mL)洗柱→循环。
(3)干燥:将过柱后的样品转入25 mL小烧杯中,真空浓缩干燥,除去提取液中的甲醇,用样品稀释液(100 mL pH 7.5 磷酸缓冲液中加0.1 mL Tween-20,0.1 g明胶)定容。
(4)测定:采用酶联免疫试剂盒进行激素抗体反应,具体步骤参照试剂盒说明书进行,试剂盒购自中国农业大学,激素含量使用xMark微孔板吸光度分光光度计(Bio-Rad,USA)测定。CTK含量为玉米素+玉米素核苷(Z+ZR)含量和异戊烯基腺嘌呤+异戊烯基腺嘌呤核苷(iP+iPR)含量的总和。
1.3 数据处理
采用Excel 2010进行数据处理并作图,采用DPS对试验数据进行显著性分析。
2 结果与分析
2.1 外源SA对灌浆期高温胁迫小麦产量和千粒重的影响
由图2(A和B)可知,与NN相比,灌浆期高温胁迫(NG)显著降低了小麦的产量。不同阶段喷施SA处理(SAHG、SAAG、SA5G和SA10G)的小麦产量均高于NG,这表明预喷施SA可降低灌浆期高温胁迫造成的小麦产量损失,缓解效果以SAAG处理最好,其次依次为SA5G、SA10G、SAHG。由图2(C和D)可知,与NN相比,灌浆期高温胁迫显著降低了小麦千粒重;SAHG、SAAG、SA5G和SA10G处理的小麦千粒重均高于NG,这表明灌浆期温度过高会导致籽粒千粒重下降,预喷施SA可降低高温胁迫导致的小麦千粒重降低程度。

NN:对照;NG:灌浆期高温胁迫;SAHG:抽穗期喷施外源SA +灌浆期高温胁迫;SAAG:开花期喷施外源SA +灌浆期高温胁迫;SA5G:花后5 d喷施外源SA +灌浆期高温胁迫;SA10G:花后10 d喷施外源SA +灌浆期高温胁迫。图柱上不同字母表示处理间差异显著(P<0.05)。
2.2 外源SA对灌浆期高温胁迫小麦成熟期强弱势粒粒重的影响
由表1可知,与NN相比,NG显著降低了不同穗位的粒重,弱势粒的下降幅度大于强势粒,不同穗位粒重下降幅度表现为上部小穗>中部小穗>下部小穗。SAHG、SAAG、SA5G和SA10G处理下,小麦不同穗位强、弱势粒粒重均高于NG,这表明预喷施SA可缓解高温胁迫下小麦不同穗位强、弱势粒粒重的损失。缓解效果以SAAG最好,其次依次为SA5G、SA10G、SAHG;在本试验条件下,外源SA对高温胁迫导致的上部小穗及弱势粒粒重的降低缓解效果较好。
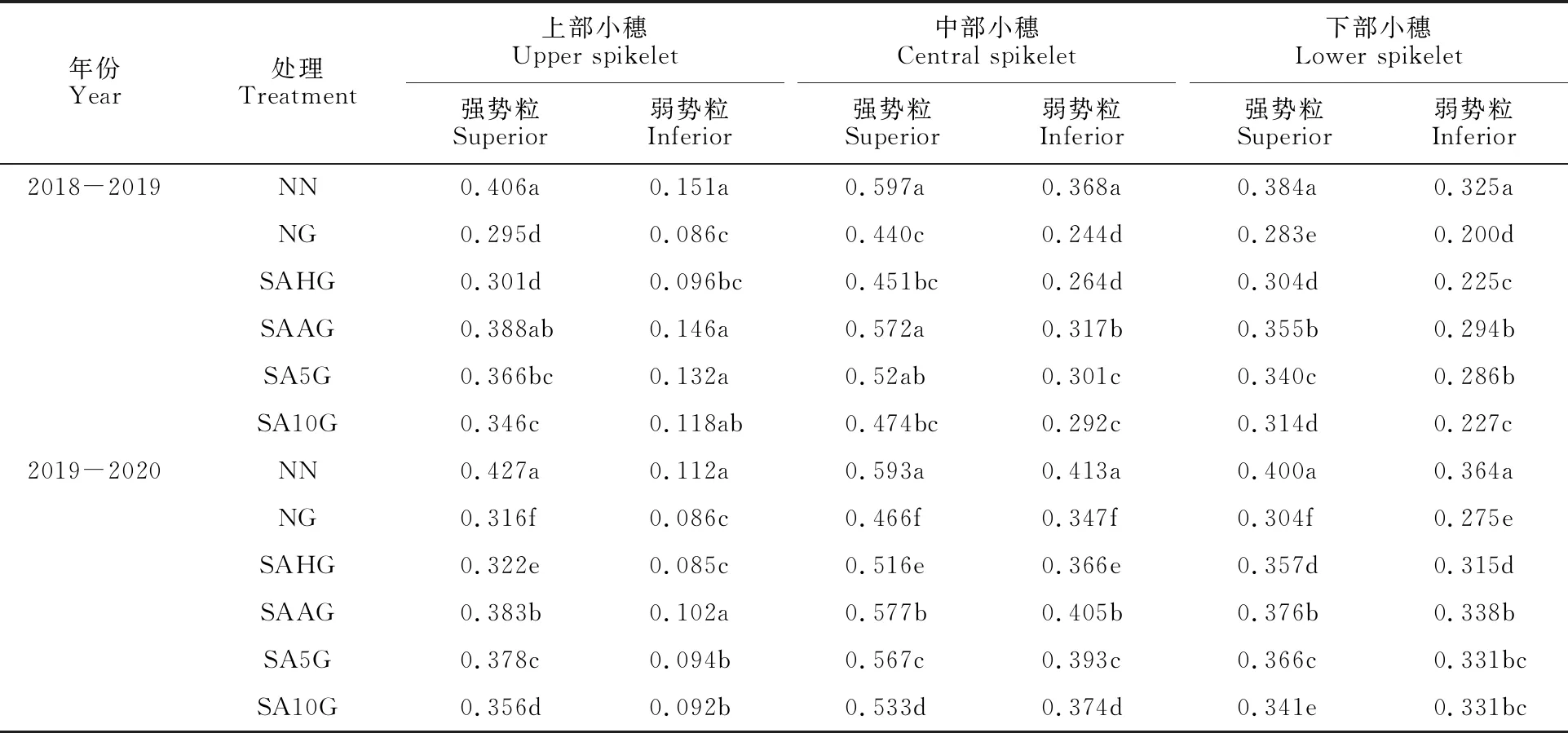
表1 不同处理下的小麦成熟期强弱势粒粒重
2.3 外源SA对灌浆期高温胁迫下小麦籽粒灌浆特性的影响
2.3.1 对粒重的影响
灌浆期高温胁迫前,与NN相比,各处理条件下(SAHG、SAAG、和SA5G)的小麦籽粒干物质积累量均显著高于NN。由图3可知,灌浆期高温胁迫后,与NN相比,NG显著降低了粒重,降低幅度在花后28 d时大于花后21 d。预喷施SA处理(SAHG、SAAG、SA5G和SA10G)的小麦粒重均显著高于NG,这表明预喷施SA缓解了高温胁迫下小麦粒重的减少,不同时期预喷施SA对灌浆期高温胁迫小麦粒重的缓解效果表现为SAAG>SA5G>SA10G>SAHG。

相同时间图柱上不同字母表示处理间在 0.05水平上差异显著。下同。
2.3.2 对籽粒蔗糖含量和可溶性糖含量的影响
由图4可知,灌浆期高温胁迫前,各处理下(SAHG、SAAG、SA5G和SA10G)的小麦籽粒蔗糖含量和可溶性糖含量均高于NN处理。灌浆期高温胁迫后,NG较NN相比显著降低了小麦籽粒蔗糖含量和可溶性糖含量,预喷施SA处理(SAHG、SAAG、SA5G和SA10G)的小麦籽粒蔗糖含量和可溶性糖含量均高于NG,这表明预喷施SA对高温胁迫下小麦籽粒蔗糖含量和可溶性糖含量的降低有所缓解,缓解效果以SAAG最好,其次依次为SA5G、SA10G、SAHG,即开花期预喷施SA对高温胁迫的缓解效果最佳。
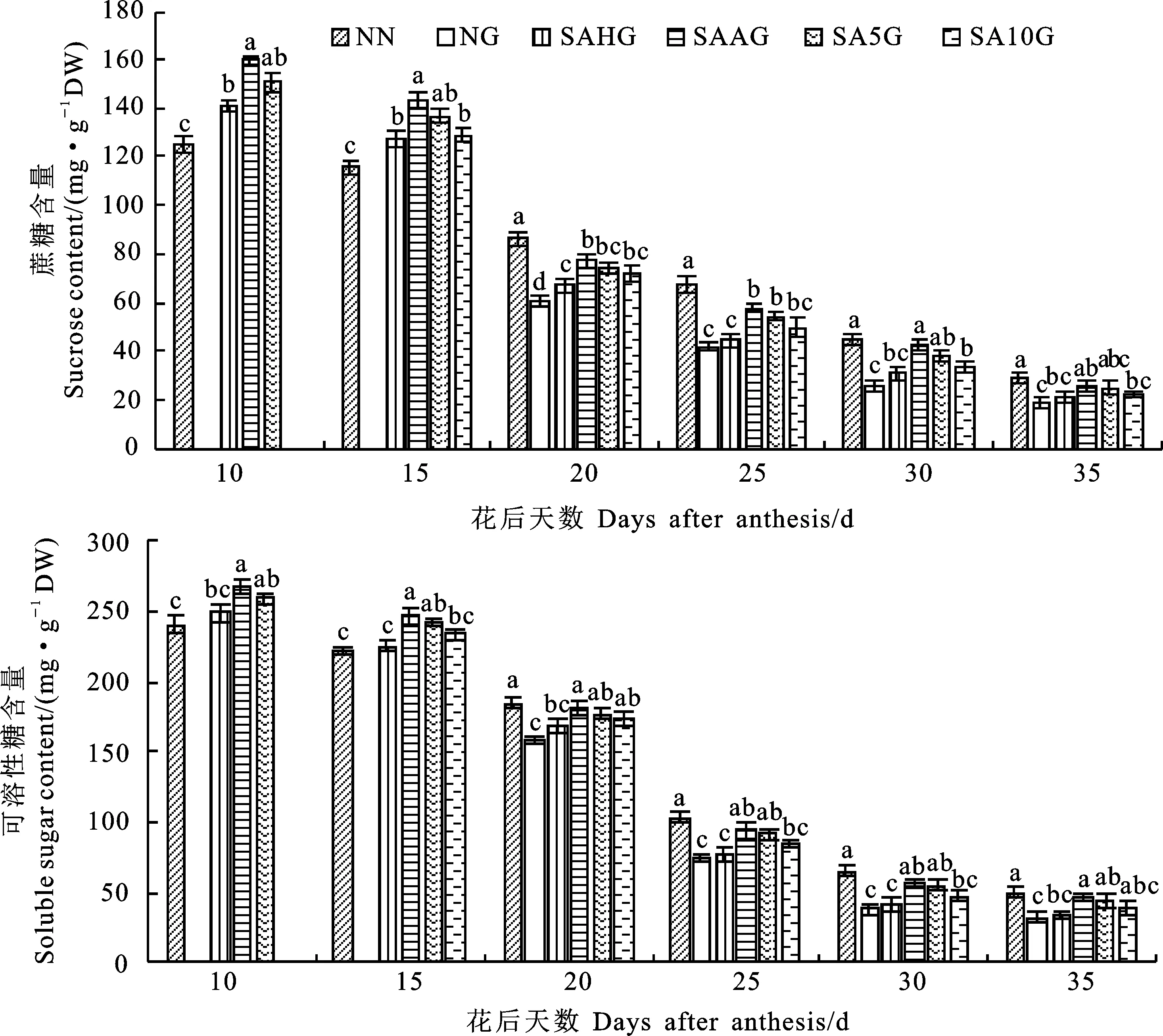
图4 外源SA对小麦籽粒蔗糖含量和可溶性糖含量的影响
2.3.3 对籽粒蔗糖合成酶(SS)活性的影响
由图5可知,小麦籽粒SS活性随着生育进程的推进呈先升后降趋势。灌浆期高温胁迫前,与NN相比,SA处理下(SAHG、SAAG、SA5G和SA10G)的小麦籽粒SS活性均高于NN。与NN相比,NG显著降低了小麦籽粒SS活性;在花后25d(SAAG、SA5G和SA10G)处理的小麦籽粒SS活性均显著高于NG,SAHG处理与NG差异不显著,这表明预喷施SA对高温胁迫导致的小麦籽粒SS活性的降低有所缓解,缓解效果以SAAG效果最好,其次依次为SA5G、SA10G、SAHG,即开花期预喷施SA对高温胁迫的缓解效果较佳。
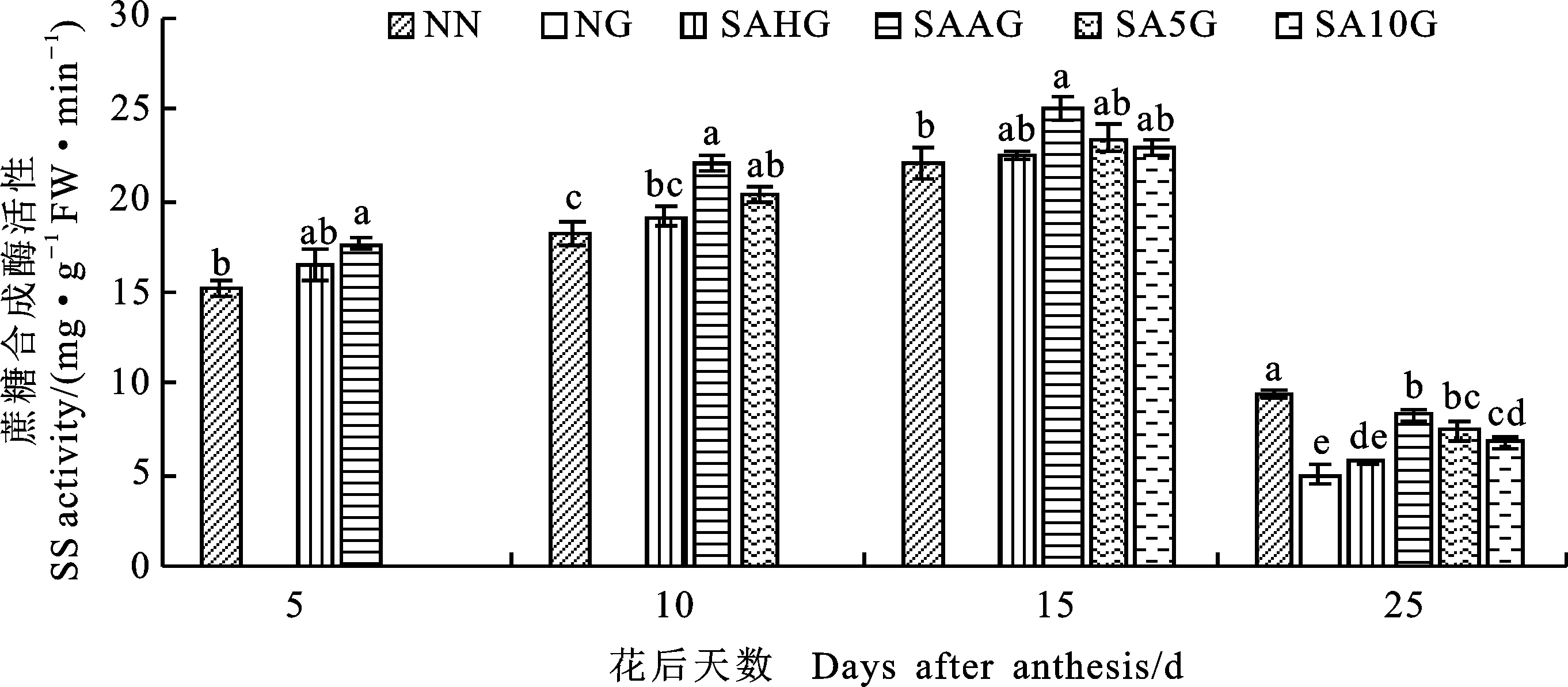
图5 外源SA对小麦籽粒蔗糖合成酶活性的影响
2.3.4 对籽粒吲哚乙酸含量的影响
由图6可知,灌浆期高温胁迫前,SAHG、SAAG、SA5G和SA10G处理的小麦籽粒IAA含量均高于NN。与NN相比,NG显著降低了小麦籽粒IAA含量,SAHG、SAAG、SA5G和SA10G处理的小麦籽粒IAA含量均显著高于NG,这表明预喷施SA缓解了高温胁迫导致的小麦籽粒IAA含量的降低,缓解效果以开花期喷施SA效果较佳。说明外源SA可提高籽粒IAA含量,缓解高温胁迫造成的IAA含量降低。

图6 外源SA对小麦籽粒吲哚乙酸含量的影响
2.3.5 对籽粒脱落酸(ABA)含量的影响
由图7可知,小麦籽粒ABA含量随着生育进程的推进呈先升后降的趋势。灌浆期高温胁迫前,与NN相比,SA处理下(SAHG、SAAG、SA5G和SA10G)的小麦籽粒ABA含量均高于NN。与NN相比,NG显著降低了小麦籽粒ABA含量;SA处理下(SAHG、SAAG、SA5G和SA10G)的ABA含量均显著高于NG,这表明预喷施SA对高温胁迫导致的小麦籽粒ABA含量的降低有所缓解,缓解效果以开花期预喷施较佳。

图7 外源SA小麦籽粒脱落酸含量的影响
2.3.6 对籽粒细胞分裂素(CTK)含量的影响
由图8可知,从花后5 d至花后25 d,小麦籽粒CTK含量呈先下降后上升趋势,在花后15 d时CTK含量最大。灌浆期高温胁迫前,SA处理下(SAHG、SAAG、SA5G和SA10G)的小麦籽粒CTK含量均高于NN。与NN相比,NG显著降低了小麦籽粒CTK含量;不同SA处理下(SAAG、SA5G和SA10G)的小麦籽粒CTK含量均显著高于NG,SAHG与NG间差异不显著,这表明预喷施SA缓解了高温胁迫下小麦籽粒CTK含量的降低, 缓解效果以开花期喷施SA效果较佳。

图8 外源SA对小麦籽粒细胞分裂素含量的影响
3 讨论与结论
3.1 外源SA对小麦产量的影响
随着全球范围内气候变暖,高温逆境频繁出现,对我国小麦产量造成严重威胁。灌浆期对小麦产量的形成至关重要,此时遭受高温胁迫会使小麦产量减少;千粒重可以作为筛选小麦抗热性的简易指标。本试验结果表明,灌浆期高温胁迫显著降低了小麦的穗粒数和千粒重,导致产量显著下降;不同阶段预喷施SA缓解了高温导致的千粒重降低,从而小麦产量的降低也得到了缓解,以开花期喷施SA 效果最佳。
对小麦不同穗位强、弱势粒粒重进行分析发现,高温胁迫下,强势粒相较于弱势粒的降低幅度小,上部小穗粒重受高温胁迫的影响最大。说明灌浆期高温胁迫主要影响小麦上部小穗和弱势粒的生长发育。中部小穗开花以及灌浆时间比下部和上部小穗更早,本试验是在灌浆中期对小麦进行高温胁迫,这可能试验中高温胁迫对中部小穗生长发育影响较小的主要原因。这与赵俊晔等强势粒开花早,灌浆启动更快,速度更快,弱势粒与其相反,同时灌浆时间晚于强势粒的观点一致。本试验结果表明,外源SA可以有效缓解高温胁迫对小麦弱势粒和上部小穗的生长发育负效应,从而缓解小麦产量的降低,开花期预喷施SA缓解效果最好。
3.2 外源SA对小麦籽粒灌浆特性的影响
灌浆特性包括灌浆时间与灌浆速率。李永庚等与Bhullar等认为,灌浆前期高温胁迫致使穗粒数显著减少,灌浆中期与后期高温胁迫使粒重降低,最终导致减产。Shah等与Plaut等认为,开花后高温胁迫使叶片光合速率下降,从而致使干物质累积速度降低。这与本试验结果一致。外源SA缓解了灌浆期高温胁迫导致的小麦籽粒干物质积累量的降低,以开花期喷施SA缓解效果最好。
非结构性碳水化合物(如可溶性糖和淀粉)是植物新陈代谢的重要物质,也是光合作用的直接产物,其在植物体内的代谢直接影响植物生长和对外界环境的响应。小麦的光合产物主要以蔗糖的形式向外输送。有研究表明,高温胁迫会抑制植物器官内光合产物向外输出,导致籽粒汁液和韧皮部中蔗糖含量下降,致使灌浆强度以及粒重急速降低。本试验结果表明,外源SA增加了籽粒中蔗糖和可溶性糖含量。蔗糖是光合产物运输到籽粒中的最初形式,SS是小麦籽粒催化淀粉合成反应的第一步关键酶,其活性反映了籽粒降解利用蔗糖的能力,在花后主要调节籽粒灌浆和淀粉积累,被认为是谷类作物“库强”的重要指标之一。籽粒蔗糖含量与旗叶蔗糖含量极显著正相关,说明旗叶光合产物(蔗糖)的供应对籽粒的蔗糖含量有显著影响。本试验结果显示,外源SA提高了籽粒SS活性,说明在SA预处理下籽粒降解蔗糖的能力增强,防止蔗糖在籽粒中过多积累,从而保证库中糖代谢的进行。缓解效果为开花期预喷施SA最好。
3.3 外源SA对小麦籽粒内源激素含量的影响
内源激素含量及其平衡是调控植物生长发育的重要因素。有研究认为,植物激素对小麦产量构成因素的形成有调节作用。不同类型小麦品种籽粒发育过程中内源激素含量及其比例变化存在差异。一般认为,内源激素的含量与胚乳细胞的分裂和物质的运输有密切关系。研究表明,SA影响植物生长、诱导形态发生与其影响激素代谢之间有一定关系,可能是通过干扰一种或两种主要激素如生长素实现的。刘 新等研究表明,水杨酸可以提高蚕豆幼苗中内源细胞分裂素和生长素的含量。本研究结果显示,正常生长条件下,小麦籽粒维持较高生理活性的时间比高温胁迫条件下长,而外源SA提高了小麦籽粒ABA含量。唐 瑭等的研究指出,ABA对SS活性有重要的调节作用,籽粒灌浆进程中内源激素通过调节SS活性变化来影响库器官中蔗糖的代谢,从而调节水稻的灌浆充实。因此,经过SA预处理的小麦可能是通过ABA调节籽粒中蔗糖淀粉代谢的关键酶(SS)活性,增大库强,从而促进籽粒的灌浆。Berleth等的研究指出,IAA在一定浓度范围内能有效调节籽粒中H-ATP酶的活性,表明IAA调控着源器官中蔗糖向库器官(籽粒胚乳细胞)的输送。Yang等的研究指出,淀粉分支酶和异淀粉酶的活性受CTK调节,CTK、IAA可通过调节细胞发育相关酶的活性来调控籽粒胚乳细胞数目、体积及内容充实物。在本研究中,外源SA提高了小麦籽粒IAA含量和CTK含量,这表明外源SA预处理有利于提高IAA和CTK对源库间糖代谢平衡的调控能力,增强籽粒蔗糖的卸载和转化能力及淀粉的合成,从而影响产量的形成。一般认为, 植物开花至成熟期间,CTK和脱落酸ABA对营养器官的衰老及其细胞内含物的转移、同化物的运输分配、籽粒贮存性碳水化合物和蛋白质的形成和积累均具有重要的调控作用。不同时期预喷施SA提高了小麦IAA、CTK和ABA的含量,其对源库间糖代谢平衡的调控能力有所增强,从而提高了籽粒的灌浆充实度。
