不同沙障材料对固沙林地面节肢动物群落结构分布的影响
2022-08-04冯永宏苟琪刘任涛罗雅曦常海涛张静蒋嘉瑜
冯永宏, 苟琪, 刘任涛,*, 罗雅曦, 常海涛, 张静, 蒋嘉瑜
不同沙障材料对固沙林地面节肢动物群落结构分布的影响
冯永宏1, 苟琪2,3, 刘任涛2,3,*, 罗雅曦2, 常海涛2, 张静1, 蒋嘉瑜2,3
1. 宁夏大学农学院, 银川 750021 2. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021 3. 宁夏大学生态环境学院, 银川 750021
为了明确沙障材料差异性对固沙林地面节肢动物群落结构的影响规律, 以腾格里沙漠东南缘2013年布设的麦草、尼龙网两种沙障模式下的固沙林地为研究样地, 于2018年春、夏、秋季, 研究了沙丘底部和顶部不同沙障模式下的固沙林地地面节肢动物群落组成及多样性分布特征。结果发现: (1)沙丘底部, 2种沙障林地地面节肢动物优势类群除了均包括蠼螋科、步甲科、叶甲科外, 麦草方格林地还包括拟步甲科和鞘翅目幼虫2个类群, 而尼龙网方格林地还包括鳞翅目幼虫1个类群; 但在沙丘顶部, 2种沙障林地地面节肢动物优势类群均为蠼螋科、步甲科、叶甲科和拟步甲科。(2)沙丘底部和顶部, 优势类群步甲科、叶甲科个体数在2种沙障林地间均无显著差异; 但优势类群拟步甲科个体数表现为麦草方格林地显著高于尼龙网方格。在沙丘底部, 蠼螋科个体数表现为2种沙障林地间无显著差异; 而在顶部, 蠼螋科个体数表现为麦草方格林地显著高于尼龙网方格。(3)沙丘底部和顶部, 地面节肢动物个体数、类群数均表现为麦草方格林地显著高于尼龙网方格, 但Shannon指数、优势度指数2种沙障林地间均无显著差异。(4)RDA和偏RDA分析结果表明, 花棒冠幅与高度、油蒿冠幅是不同微生境中地面节肢动物个体数分布产生差异的主要因素。(5)研究表明, 沙障材料对地面节肢动物群落组成、个体数和类群数分布均产生显著影响, 而且地面节肢动物群落组成分布亦受到沙丘微地形的影响, 但沙障材料对地面节肢动物多样性分布的影响较小。
腾格里沙漠; 沙障材料; 沙丘微地形; 地面节肢动物; 群落多样性
0 前言
沙漠化是全球性严重的环境问题之一, 全球现有约4560万km2的沙漠化土地, 并且以每年5—7万km2的速度增长[1]。沙漠化不仅造成严重的生态系统失衡, 而且给我国农业生产和人民生活带来严重影响[2]。因此, 需要采取合理的人工沙漠化防治措施来改善生态环境。在腾格里沙漠东南缘, 进行固沙林营造并扎设草方格, 可以有效改善土壤理化性质, 改良土壤结构, 促进退化土壤修复[3–5], 已被认为是成功的生态恢复措施而得到了大面积推广。然而, 不同沙障材料由于其本身特性及价格成本的差异性, 往往会影响沙漠固沙造林工程的实施效果和固沙造林后植被恢复过程[6]。并且, 地面节肢动物及其多样性是固沙林地生态系统中生物多样性的重要组成部分, 在物质转换、凋落物分解、土壤发育和成熟等生态功能过程中扮演着重要的角色[7], 能够直接反映固沙林地生态恢复过程和进程。因此, 研究麦草、尼龙网等不同沙障模式下的固沙林地地面节肢动物群落组成及多样性分布特征, 对于干旱风沙区生物多样性保育、人工植被建设以及沙漠化防治均具有重要的理论与实践意义。
目前, 学者们已从不同角度对不同材料沙障的固沙效应做了较多研究[8–11]。孙涛等[11]在民勤发现尼龙网沙障随时间的延长仍具有较好的固沙功能, 而麦草沙障的固沙功能随时间的增加逐渐降低。王雨浩[12]在腾格里沙漠的东南缘发现在沙障规格相同的情况下, 麦草沙障的防风固沙效益要优于聚乳酸纤维沙障。研究表明, 不同材料草方格固沙效应的差异性与沙障本身特性及应用成本密切相关。例如, 尼龙网沙障作为一种新型的防护材料, 具有造价相对低廉, 抗老化, 抗风沙, 特别是在一些风沙灾害比较严重的地区, 尼龙网防护能快速达到立竿见影的效果[3]; 但尼龙网方格更是一种物理性防沙技术, 且不易分解而造成塑料污染。而草方格沙障采用的麦草为生物材料, 既可以作为枯落物, 其分解可以为沙漠微生物提供养分来源, 又具有保水性能[5]; 但缺点是分解较快, 且分解之后的固沙效果降低。因此, 不同材料沙障在固沙效应方面均存在自己的优势和劣势, 将会对固沙林地的土壤-植被系统恢复进程产生不同的影响。但是, 不同沙障材料对固沙林地地面节肢动物群落结构分布的影响规律, 尚不清楚。并且, 以往研究多针对地形条件相对平坦的沙地中某一种固沙材料的研究[13]。例如, 屈建军等[8]对半隐蔽格状沙障的综合防护效益研究中, 发现同一规格的格状沙障往往由于设置的地形部位不同,其防护作用可能产生很大的差异。已有研究表明, 微地形变化可以对沙质荒漠拟步甲科昆虫分布产生深刻影响[13]。但是, 关于草方格固沙林地中地面节肢动物多样性对较大起伏地形的适应性是否存在差异研究, 报道较少。
鉴于此, 在腾格里沙漠东南缘选取麦草、尼龙网沙障模式下的人工固沙林地为研究对象, 在沙丘底部和顶部生境中布设调查样点, 采用陷阱诱捕法对地面节肢动物群落组成、多样性分布特征进行调查, 结合环境分析, 旨在阐明不同沙障材料对固沙林地地面节肢动物群落结构分布的影响规律, 丰富沙漠生态系统土壤动物生态学的研究内容, 为人工林建设、采取有效的防沙治沙措施及退化生态系统恢复提供依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏回族自治区中卫市腾格里沙漠东南缘(37°25′N, 104°45′E), 海拔1350—1450 m, 属于典型的温带荒漠化草原向草原化荒漠的过渡地区。年平均气温10.0 ℃, 最低温-25.1 ℃, 最高温38.1 ℃。年平均降水量186.2 mm, 且分布不均匀, 降水主要集中于7—9月, 全年日照时间3264 h。全年主要西北风居多, 年均风速2.9 m·s–1。土壤以细沙粒为主, 以风沙土及灰棕荒漠土分布最广。该区地形起伏变化较大, 景观主要是高大且密集的网格状沙丘, 沙丘从西北向东南倾斜, 且呈阶梯状分布。
在世界银行贷款项目“宁夏黄河东岸防沙治沙林业生态工程”中卫人工固沙植被项目的资助下, 于2013年开始新一轮草方格人工植被固沙工程建设。麦草沙障是用长条状麦草捆设置成1 m×1 m的网格状, 地面以上15 cm, 地面埋入沙内深度15 cm; 尼龙网沙障是用尼龙网材料设置成1 m×1 m的网格状, 地面以上15cm, 地面埋入沙内深度15cm。种植规格为: 1 m×3 m和1 m×1 m(株距×行距)分别种植花棒和柠条, 并在沙障内撒播油蒿种子。经过近5年的固沙恢复, 沙丘底部麦草方格内花棒、柠条和油蒿密度分别为0.14株·m–2、0.14株·m–2、11.6株·m–2, 尼龙网方格内花棒、柠条和油蒿密度分别为0.19株·m–2、0.19株·m–2、7.4株·m–2; 沙丘顶部麦草方格内花棒、柠条和油蒿密度分别为0.11株·m–2、0.08株·m–2、12.4株·m–2, 尼龙网方格内花棒、柠条和油蒿密度分别为0.61株·m–2、0.06株·m–2、2.8株·m–2。灌丛植物种包括: 柠条()、油蒿()、花棒()、沙拐枣()和沙木蓼()等旱生灌木。
1.2 试验设计与取样调查
1.2.1 试验设计
于2018年春季, 在腾格里沙漠东南缘2013年开始建设的人工灌丛固沙林地中, 分别选择麦草、尼龙网两种沙障模式下的林地为研究样地, 每种沙障模式下的林地设置5个重复样区, 每个样地面积均为900 m2, 间距为50 m左右。在每种样地, 选择丘顶和丘底分别布设调查样点4个, 共布设2沙丘微生境×2沙障×5重复×4个样点=80样点。
1.2.2 地面节肢动物取样调查
分别于2018年春季(4月)、夏季(7月)、秋季(9月), 在每个调查样点, 采用国际通用的陷阱诱捕法采集地面节肢动物[14]。具体方法为: 在每个调查样点的中心位置将塑料杯(上下直径分别为14 cm和7 cm, 高度为10 cm)埋入土中, 杯口略低于地面, 同时在其内加入少量甘油和体积分数为3%的福尔马林溶液, 以增加诱捕的有效性。每次持续时间均为12 d(每隔3 d检查收集1次, 避免采集标本的损失)。试验结束后将收集到的地面节肢动物标本带回实验室进行分类鉴定。
地面节肢动物标本分类鉴定主要依据《宁夏贺兰山昆虫》[15]、《中国土壤动物检索图鉴》[16]和《昆虫分类》[17]等。根据不同地面节肢动物类群在群落总个体数中所占比例来划分等级, 其划分标准为优势类群(占群落总个体数的10%以上)、常见类群(1%—10%)和稀有类群(<1%)[18]。
1.2.3 植被调查、土壤样品采集与分析
在每个调查样点布设5 m×5 m的样方, 测定柠条和花棒的高度(cm)、冠幅(m2)指标。同时, 在5 m× 5 m的样方内, 按照“M”型设置5个1 m×1 m的草方格, 调査沙障内油蒿的高度(cm)、冠幅(m2)指标。由于灌丛植被同一年中不同季节变化不大, 故只调查1次。
在每个调查样点内, 利用5点取样法采集混合土样装入自封袋(取样深度为0—10 cm), 放置在保温箱中, 土壤样品带回实验室, 取混合土样中的1/4用于土壤含水量的测定, 剩余3/4土样经自然风干后, 过2 mm土壤筛, 以去除树根、叶片等杂物, 以供土壤理化性质测定使用。具体测定指标包括: 土壤温度、容重、含水量、pH、电导率、粒径组成、有机碳和全氮共8个指标。
土壤温度(℃)采用地温仪测定。土壤容重(采用环刀法在每个调查样点取原状土样1个; g·cm–3)和土壤含水量(%)均采用烘干称重法测定。土壤pH和土壤电导率(μS·m–1)均采用1:5土水比浸提后, 分别使用P4多功能仪(Muiti-line P4 Universal Meter, WTW公司, Germany)和便携式电导率仪进行测定[19]。土壤粒径组成采用Mastersizer3000激光衍射粒度分析仪进行测定。根据美国农业部(USDA)制土壤质地分级标准将本试验土壤样品质地划分为: 细沙粒(100—250 μm)、极细沙粒(50—100 μm)和粉粒(2—50 μm)3个等级[1]。土壤有机碳(%)和全氮(%)分别采用重铬酸钾氧化外加热法和凯氏定氮法(意大利DK6, UDK140分析仪)测定[20]。
1.3 数据处理
分别将春、夏、秋季每个样地采样点数据进行整理, 用于统计地面节肢动物个体数(只·陷阱–1)、类群数以及计算群落多样性, 包括Shannon指数和优势度指数。然后, 利用平均值法将3个季的数据进行平均。
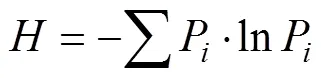
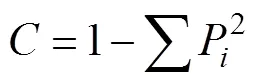
式中,P表示类群的个体数在总个体中的比例, 即P=/。
所有数据采用SPSS 25.0软件进行统计分析。通过独立样本检验分析同一位置不同沙障材料调查样点间地面节肢动物个体数、类群数、多样性指数的差异性。统计在<0.05水平上存在显著差异。应用Origin 2018进行作图。
利用已建立的4种不同生境地面节肢动物个体数据矩阵, 采用DCA分析(Detrended correspondence analysis, DCA)确定排序轴的梯度长度(Lengths of gradient, LGA)。依据LGA值大小选择适宜的排序分析方法, 其基本准则是: 当LGA<4时, 宜采用线性模型排序; 当LGA>4时, 宜采用非线性模型排序[21]。由于本文地面节肢动物群落DCA计算出的LGA=1.779<4, 故采用线性模型的冗余分析(Redundancy analysis, RDA)来确定地面节肢动物个体数分布与环境因子间的关系。同时, 采用偏RDA分析(Partial RDA)和蒙特卡洛置换检验(Monte Carlo permutation test), 定量评价每个环境因子对地面节肢动物群落分布变化的贡献率(即独立解释量)。在偏RDA分析的基础上, 选择对对地面节肢动物群落分布有较高影响的解释变量, 绘制地面节肢动物类群分布与解释变量关系的二维排序图(Biplot)。应用CANOCO4.5进行分析运算。为了保证试验数据满足正态分布及减小异常值对分析结果的影响, 对动物个体数量及环境因子数据进行平方根转换, 对样方进行中心化和标准化(Center and standardize)并对物种数据进行误差方差标准化(Standardize by error variance)处理。
2 结果与分析
2.1 环境特征
由表1可以看出, 无论是在沙丘底部还是顶部, 土壤有机碳、pH、含水量、电导率、容重、土壤粉粒和温度均表现为麦草方格和尼龙网方格间无显著差异(>0.05)。但是, 在沙丘底部, 仅土壤极细沙粒含量表现为麦草方格显著高于尼龙网方格(<0.05); 在沙丘顶部, 仅土壤全氮和细沙粒含量表现为麦草方格显著低于尼龙网方格(<0.05)。
由表2可知, 无论沙丘底部还是顶部, 柠条高度表现为麦草方格显著低于尼龙网方格(<0.05); 而油蒿高度表现为麦草方格和尼龙网方格间无显著差异(>0.05)。在沙丘底部, 花棒冠幅表现为麦草方格显著低于尼龙网方格(<0.05), 而柠条冠幅、花棒高度、油蒿冠幅均表现为麦草方格和尼龙网方格间无显著差异(>0.05); 在沙丘顶部, 柠条冠幅、油蒿冠幅均表现为麦草方格显著低于尼龙网方格(<0.05), 花棒高度表现为麦草方格显著高于尼龙网方格(<0.05), 而花棒冠幅表现为麦草方格和尼龙网方格间无显著差异(>0.05)。
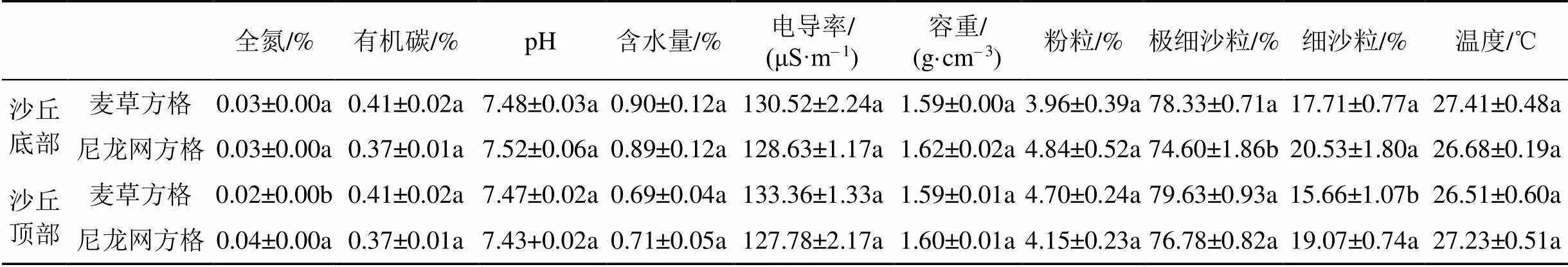
表1 不同材料沙障沙丘底部和顶部土壤理化性质(均值±标准误)
注: 同列不同字母表示同一沙丘位置不同沙障材料处理间差异显著(<0.05)。

表2 不同材料沙障沙丘底部和顶部地表植被指标(均值±标准误)
注: 同列不同字母表示同一沙丘位置不同沙障材料处理间差异显著(<0.05)。
2.2 地面节肢动物群落组成与数量特征
本次调查共计获得地面节肢动物659只。鉴定隶属于10目25科27个类群(表3)。其中, 优势类群为步甲科、拟步甲科、蠼螋科和叶甲科, 其个体数分别占总个体数的30.20%、18.51%、16.24%和13.20%。常见类群为鞘翅目幼虫、鳞翅目幼虫、蚱蜢科、金龟甲科和平腹蛛科, 其个体数分别占总个体数的6.53%、3.19%、2.28%、2.12%和1.67%。其余18个类群为稀有类群, 其个体数仅占总个体数的6.07%。
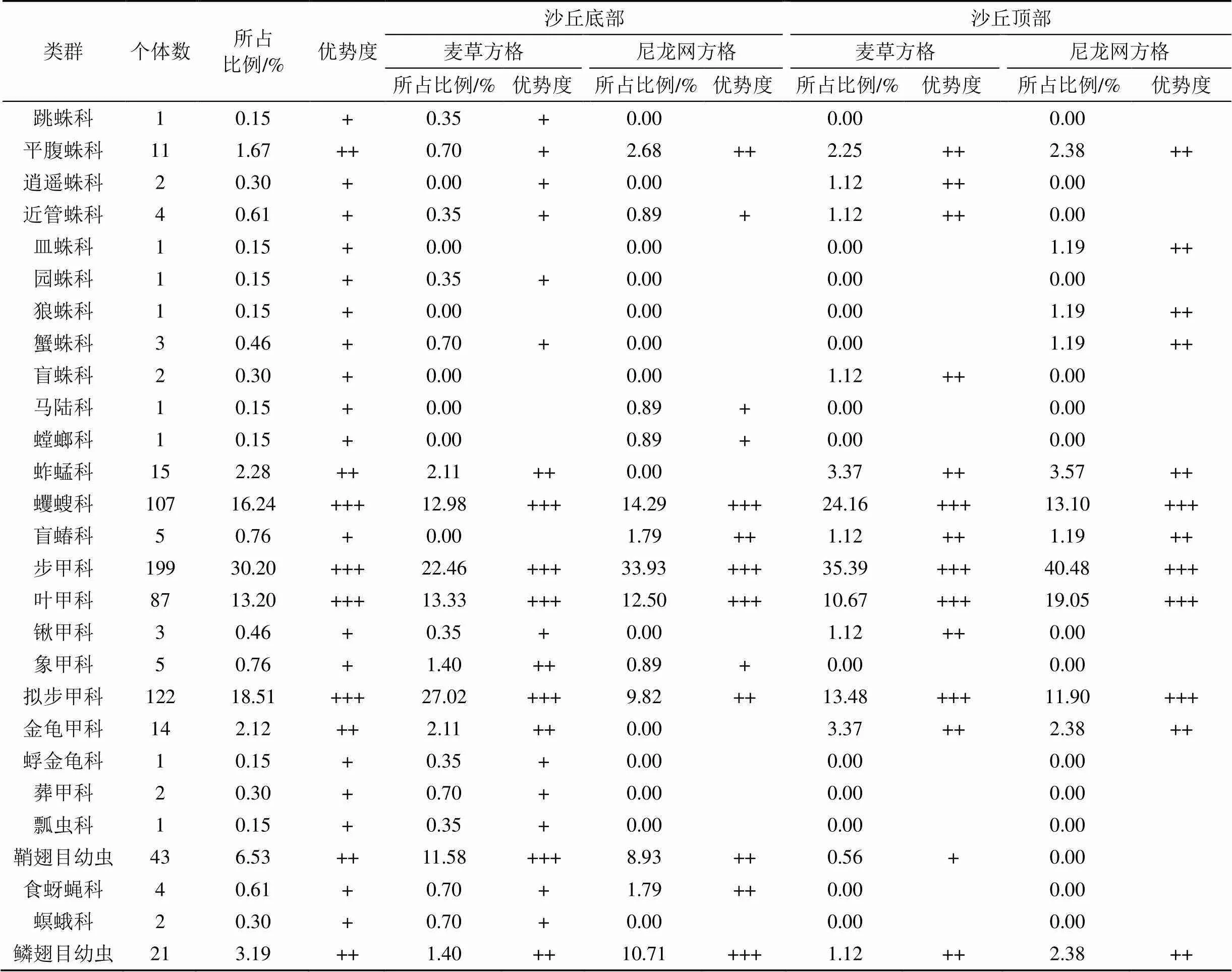
表3 地面节肢动物群落组成及多度分布
注: 个体数占全部捕获量的10%以上为优势类群, 用+++表示; 介于1%—10%为常见类群, 用++表示; 介于0.1%—1%为稀有类群, 用+表示。
在沙丘底部, 麦草方格固沙林地内共捕获地面节肢动物285只、18科、20类群。其中, 优势类群包括拟步甲科、步甲科、叶甲科、蠼螋科和鞘翅目幼虫5个类群, 个体数分别占总个体数的27.02%、22、46%、13.33%、12.98%和11.58%; 常见类群包括蚱蜢科、金龟甲科、象甲科和鳞翅目幼虫4个类群, 个体数分别占总个体数的2.11%、2.11%、1.4%和1.4%; 其余11个类群为稀有类群, 其个体数仅占总个体数的5.61%。尼龙网方格固沙林地内共捕获地面节肢动物112只、10科、12类群。其中, 优势类群包括步甲科、蠼螋科、叶甲科和鳞翅目幼虫4个类群, 个体数分别占总个体数的33.93%、14.29%、12.5%和10.71%; 常见类群包括拟步甲科、鞘翅目幼虫、平腹蛛科、盲蝽科和食蚜蝇科5个类群, 个体数分别占总个体数的9.82%、8.93%、2.68%、1.79%和1.79%; 其余3个类群为稀有类群, 其个体数仅占总个体数的3.57%。
在沙丘顶部, 麦草方格固沙林地内共捕获地面节肢动物178只、12科、14类群, 其中, 优势类群包括步甲科、蠼螋科、拟步甲科和叶甲科4个类群, 个体数分别占总个体数的35.39%、24.16%、13.48%和10.67%; 常见类群包括蚱蜢科、金龟甲科、平腹蛛科、锹甲科、逍遥蛛科、近管蛛科、盲蛛科、鳞翅目幼虫和盲蝽科9个类群, 个体数分别占总个体数的3.37%、3.37%、2.25%、1.12%、1.12%、1.12%、1.12%、1.12%和1.12%; 其余1科为稀有类群, 其个体数仅占总个体数的0.56%。尼龙网方格固沙林地内共捕获地面节肢动物84只、11科、12类群, 其中, 优势类群包括步甲科、叶甲科、蠼螋科和拟步甲科4个类群, 个体数分别占总个体数的40.48%、19.05%、13.1%和11.9%; 常见类群包括蚱蜢科、金龟甲科、平腹蛛科、鳞翅目幼虫、皿蛛科、狼蛛科、蟹蛛科和盲蝽科8个类群, 个体数分别占总个体数的3.57%、2.38%、2.38%、2.38%、1.19%、1.19%、1.19%和1.19%; 无稀有类群。
从图1可以看出, 无论在沙丘底部还是顶部, 步甲科、叶甲科个体数均表现为麦草方格和尼龙网方格间无显著差异(>0.05), 而拟步甲科个体数均表现为麦草方格显著高于尼龙网方格(<0.05)。在沙丘底部, 蠼螋科个体数表现为麦草方格和尼龙网方格间无显著差异(>0.05); 但在沙丘顶部, 蠼螋科个体数表现为麦草方格显著高于尼龙网方格(<0.05)。
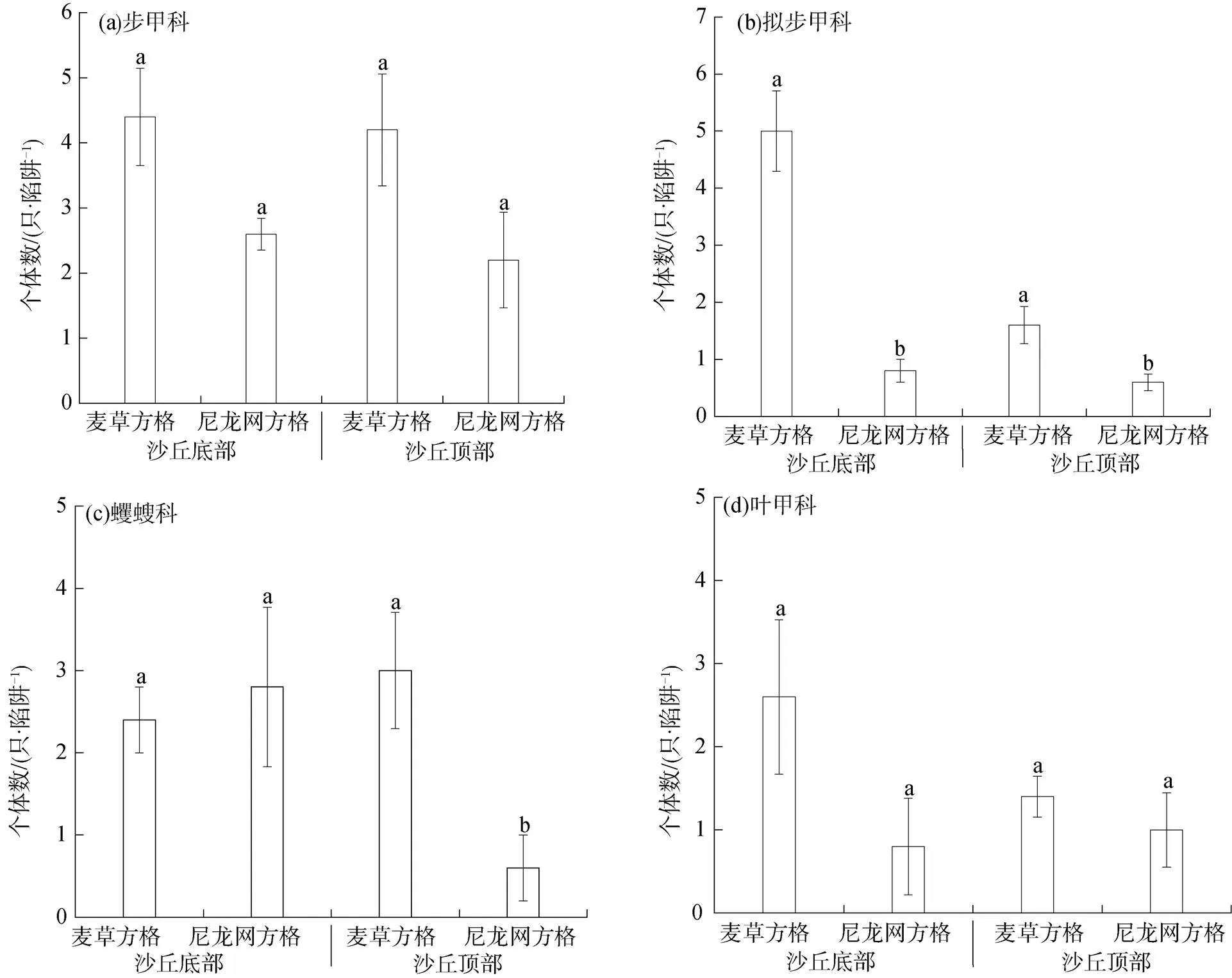
注: 不同字母表示同一沙丘位置不同沙障材料处理间差异显著(P<0.05)。
Figure 1 The abundance of dominant arthropod groups at dune bottom and top within checkerboard of different materials
2.3 地面节肢动物群落多样性
从图2可以看出, 无论在沙丘底部还是顶部, 地面节肢动物个体数、类群数均表现为麦草方格显著高于尼龙网方格(<0.05); 但地面节肢动物Shannon指数、优势度指数均表现为麦草方格和尼龙网方格间无显著差异(>0.05)。
2.4 地面节肢动物个体数分布与环境因子间的相关分析
由表4可知, 步甲科、拟步甲科个体数均与柠条高度和冠幅呈负相关关系(<0.05), 而与土壤总氮、极细沙粒、细沙粒、花棒高度、冠幅、油蒿冠幅之间均无相关性(0.05)。蠼螋科个体数与柠条高度和油蒿冠幅呈负相关关系(<0.05), 而与土壤总氮、极细沙粒、细沙粒、花棒高度、冠幅、柠条冠幅之间均无相关性(0.05)。叶甲科个体数与土壤总氮、极细沙粒、细沙粒、花棒高度、冠幅、柠条高度、冠幅、油蒿冠幅之间均无相关性(0.05)。
对地面节肢动物个体数与环境因子间关系的RDA排序分析表明, 第一典型轴(=2.442,=0.014)和所有典型轴(=1.647,=0.002)在统计学上达到显著水平, 说明排序分析能够较好地反映地面节肢动物个体数与环境因子的关系。并且, 前两个排序轴累积解释了29.5%(第一排序轴解释了18.3%和第二排序轴解释了11.2%)的地面节肢动物群落变异。从图3中可以看出, 沙丘底部麦草方格和尼龙网方格主要在第二、三象限, 而顶部麦草方格与尼龙网方格主要在第一、四象限。
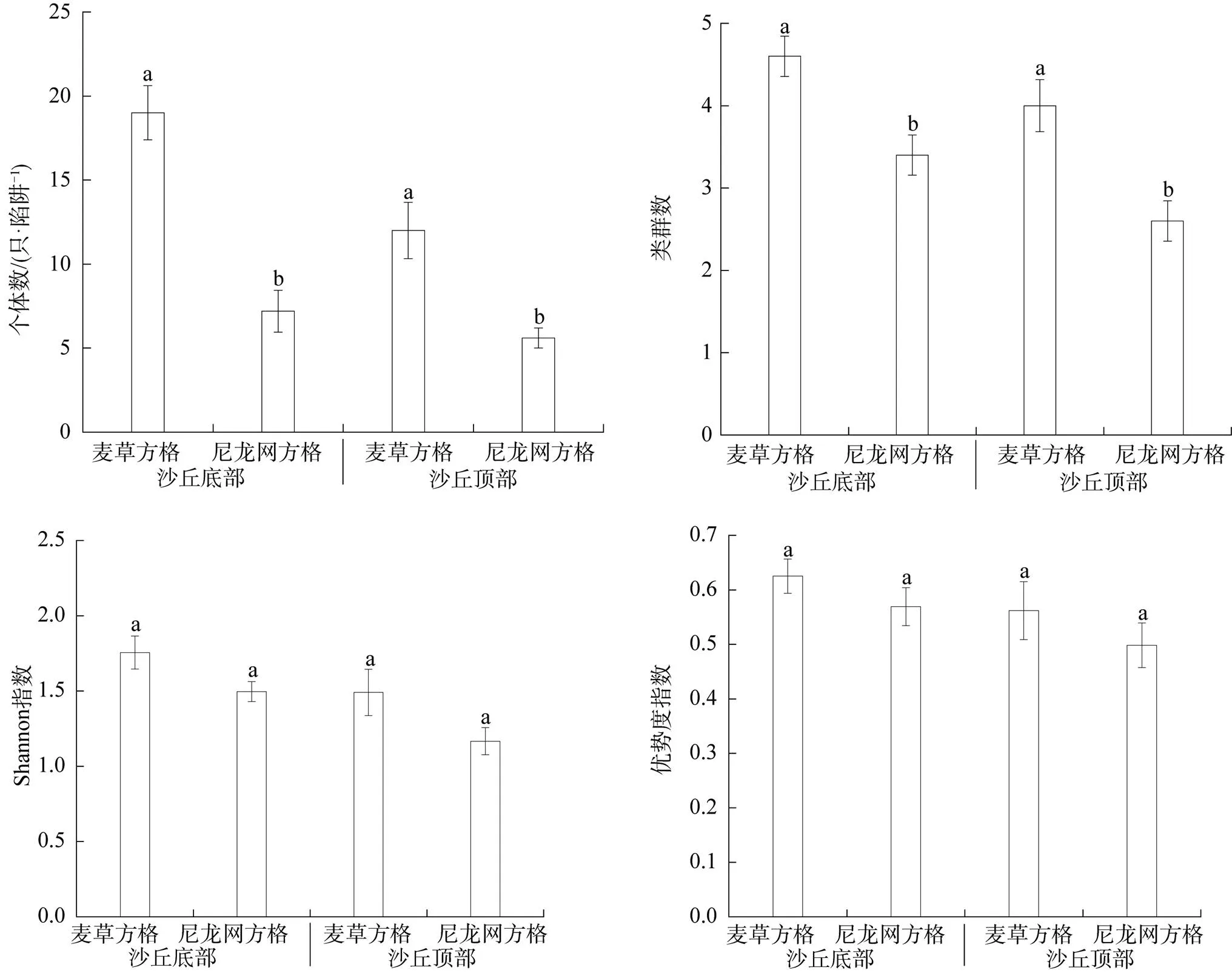
注: 不同字母表示同一沙丘位置不同沙障材料处理间差异显著(P<0.05)。
Figure 2 The arthropod community index at dune bottom and top within checkerboard of different materials
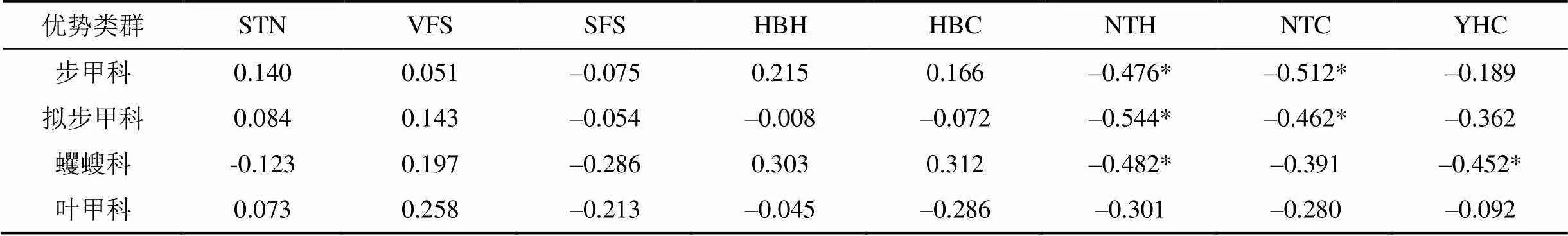
表4 优势类群个体数与环境因子间的Pearson相关系数
注: STN: 土壤总氮; VFS: 土壤极细沙粒; SFS: 土壤细沙粒; HBH: 花棒高度; HBC: 花棒冠幅; NTH: 柠条高度; NTC: 柠条冠幅; YHC: 油蒿冠幅。*表示在0.05水平(双侧)显著相关。
偏RDA分析表明(表5), 花棒冠幅与高度、油蒿冠幅对地面节肢动物个体数分布产生显著影响(<0.05), 贡献率分别为9%、9%和8%, 而其余环境因子对地面节肢动物个体数影响不显著(>0.05)。
3 讨论
3.1 不同沙障材料对土壤与植被分布的影响
本研究中, 在沙丘顶部, 土壤全氮表现为麦草方格显著低于尼龙网方格。其原因可能是沙丘顶部尼龙网方格中冠幅较大的柠条根系含有大量的根瘤菌, 可以固定空气中的氮素, 进而有利于其土壤全氮的积累[22]。土壤细沙粒表现为麦草方格显著低于尼龙网方格。其原因可能是较大的柠条冠幅和油蒿冠幅可以降低风速[19], 从而导致质量较重的土壤细沙粒沉积在沙丘顶部。土壤极细沙粒含量表现为麦草方格和尼龙网方格间无显著性差异。原因可能是沙丘顶部风速大, 导致顶部麦草方格和尼龙网方格内极细沙粒均随风的运移沉积在沙丘其他部位[6], 故2种沙障间极细沙粒无显著差异。但是在沙丘底部, 土壤极细沙粒含量表现为麦草方格显著高于尼龙网方格。分析原因可能是在麦草沙障林地中, 由于油蒿密度较大, 覆盖度较高, 而且油蒿冠层结构致密低矮便于细粒物质的捕获保存[23], 同时油蒿根系发达并且主要在浅层土壤连续活跃活动[24], 因而其根系分泌物对土壤颗粒也有一定改善作用; 而尼龙网沙障样地则以花棒和柠条居多, 由于近地层分枝较少且稀疏, 使其易受风蚀影响, 沙层表面较细颗粒相对不易积累[25], 同时, 花棒和柠条为深根系植物[26], 因此对浅层土壤颗粒的影响相对较小。无论在沙丘底部还是顶部, 土壤有机碳、pH、含水量、电导率、容重、粉粒和温度均表现为麦草方格与尼龙网方格间无显著差异, 这与罗雅曦[19]在腾格里沙漠的研究结果相似。可能原因是麦草容易腐烂, 经过5年时间导致麦草所起的保持水分和提高养分的作用随之减弱[27]。因此, 不同沙障材料对土壤有机碳、pH、含水量、电导率、容重、粉粒和温度的影响较小。
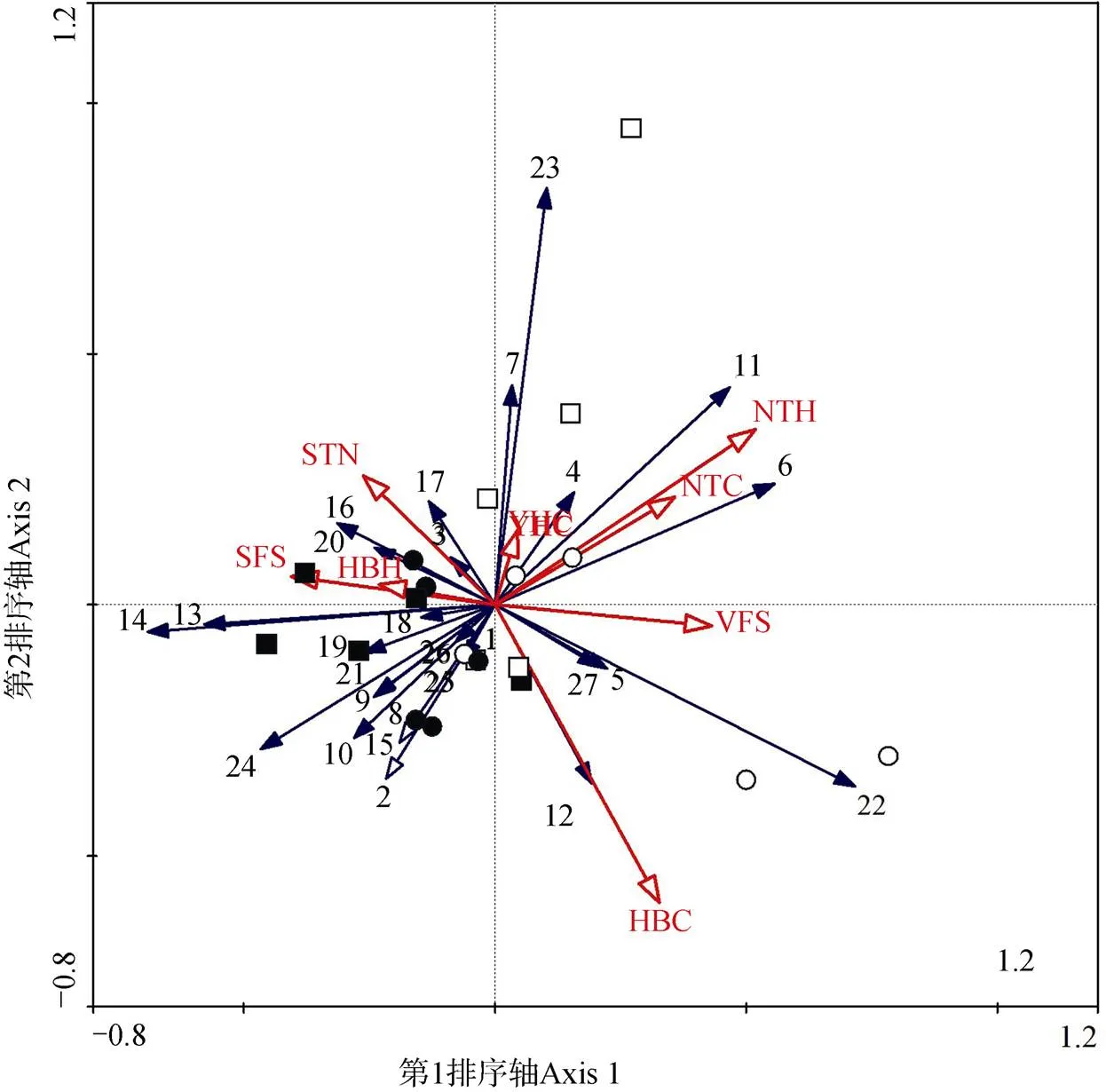
注: HBC: 花棒冠幅; HBH: 花棒高度; NTH: 柠条高度; SFS: 土壤细沙粒; VFS: 土壤极细沙粒; STN: 土壤总氮; NTC: 柠条冠幅; YHC: 油蒿冠幅; 1: 步甲科; 2: 拟步甲科; 3: 螟蛾科; 4: 叶甲科; 5: 金龟甲科; 6: 蚱蜢科; 7: 盲蝽科; 8: 瓢虫科; 9: 跳蛛科; 10: 象甲科; 11: 平腹蛛科; 12: 锹甲科; 13: 食蚜蝇科; 14: 鞘翅目幼虫; 15: 蠼螋科; 16: 近管蛛科; 17: 皿蛛科; 18: 园蛛科; 19: 蟹蛛科; 20: 蜉金龟科; 21: 葬甲科; 22: 逍遥蛛科; 23: 狼蛛科; 24: 鳞翅目幼虫; 25: 螳螂科; 26: 马陆科; 27: 盲蛛科; ●=底部麦草方格; ■=底部尼龙网方格; ○=顶部麦草方格; □=顶部尼龙网方格。
Figure 3 RDA two-dimensional diagram of the relationship between ground-active arthropods community distribution and environmental factors
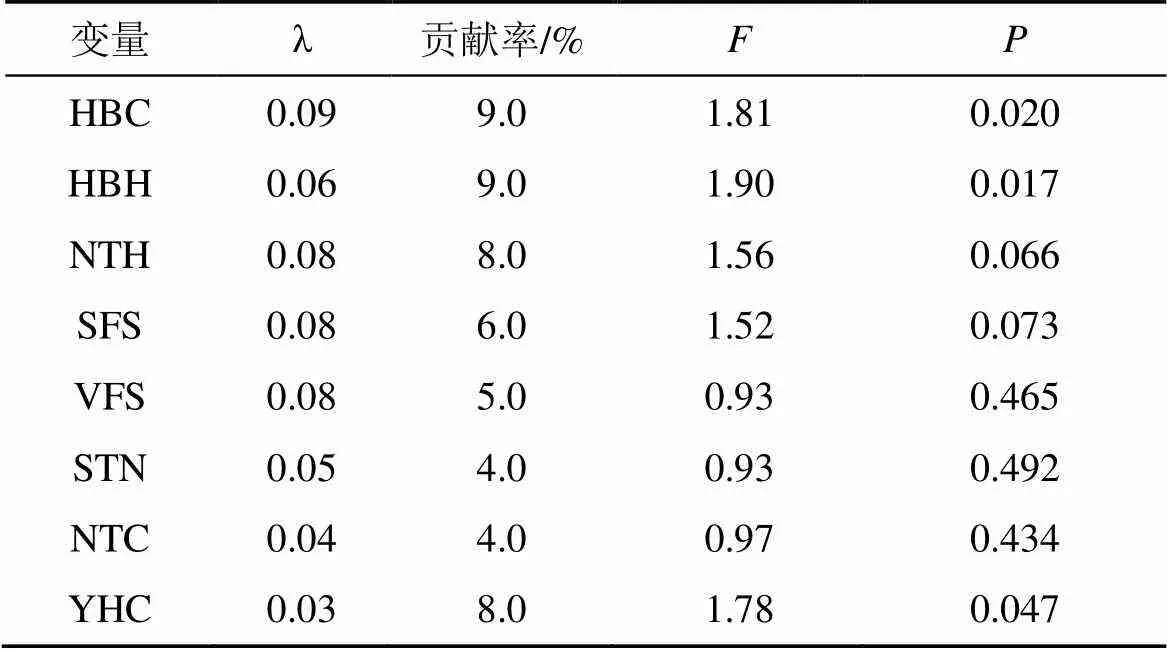
表5 环境因子对地面节肢动物个体数变化的相对贡献偏RDA分析
注: λ: 边际效应; HBC: 花棒冠幅; HBH: 花棒高度; NTH: 柠条高度; SFS: 土壤细沙粒; VFS: 土壤极细沙粒; STN: 土壤总氮; NTC: 柠条冠幅; YHC: 油蒿冠幅。
无论在沙丘底部还是顶部, 柠条高度均表现为麦草方格显著低于尼龙网方格, 表明不同沙障材料对柠条高度存在显著影响。分析原因可能是麦草寿命短, 容易腐烂, 扎设一定时间后, 某一部位麦草被风蚀裸露后可能会加速邻近区域麦草的裸露, 麦草所起的保持水分和提高养分的作用随之减弱[27]。因此, 对于需水量相对较大的柠条难以维持其自身长时间的生长发育及其繁衍, 从而导致柠条高度呈现出麦草方格显著低于尼龙网方格。在沙丘底部, 柠条和油蒿冠幅均表现为麦草方格和尼龙网方格间无显著差异。原因可能是沙丘底部土壤含水量均较高[27], 故2种沙障间柠条和油蒿冠幅无显著差异。但在沙丘顶部, 柠条和油蒿冠幅表现为麦草方格显著低于尼龙网方格。分析原因可能是麦草沙障抗风沙侵蚀能力减弱[6], 从而导致柠条和油蒿冠幅生长较慢。在沙丘底部, 花棒高度表现为麦草方格与尼龙网方格间无显著差异, 而在沙丘顶部表现为麦草方格显著高于尼龙网方格。其原因可能是沙丘底部麦草方格和尼龙网方格内土壤含水量均较高[28–29], 故花棒高度无显著差异; 而在沙丘顶部, 麦草沙障可以作为枯落物, 其分解可以为沙漠微生物提供养分来源[5], 故麦草沙障对花棒高度影响较尼龙网沙障大。
3.2 不同沙障材料对地面节肢动物群落组成与多样性分布的影响
在腾格里沙漠生态系统中, 随沙障材料和沙丘微地形变化、土壤理化性质和植被改变的同时, 地面节肢动物群落组成与结构分布也随之发生显著变化。通过比较不同沙障材料林地间地面节肢动物优势类群的组成, 发现在沙丘底部, 优势类群除了共有步甲科、蠼螋科、叶甲科3个类群外, 麦草方格固沙林地还包括拟步甲科和鞘翅目幼虫2个类群, 而尼龙网方格固沙林地内优势类群则还包括鳞翅目幼虫1个类群。说明麦草方格固沙林地更适合拟步甲科和鞘翅目幼虫生存, 而尼龙网方格固沙林地更适合鳞翅目幼虫生活。分析原因可能是沙丘底部麦草方格固沙林地内柠条高度显著低于尼龙网方格, 而柠条、花棒密度较低, 导致地面阻力较小, 有利于拟步甲科个体的移动和活动[13], 进而导致拟步甲科个体数分布较多, 故在沙丘底部拟步甲科个体数表现为麦草方格显著高于尼龙网方格, 这与娄巧哲等[30]在古尔班通沙漠的研究结果类似。同时, 麦草沙障可以作为枯落物, 其分解可以为沙漠鞘翅目幼虫提供食物来源[5], 故麦草方格固沙林地有利于拟步甲科和鞘翅目幼虫生存。但与此相反的是, 尼龙网方格林地柠条高度较大, 为鳞翅目幼虫取食提供了有利的食物资源。鳞翅目幼虫常取食叶片包括卷叶、缀叶、结鞘、吐丝结网或钻入植物组织取食[31], 故尼龙网方格固沙林地更适合鳞翅目幼虫生存。在沙丘顶部, 拟步甲科则是2种沙障材料林地的共同优势类群, 说明拟步甲科个体数更适应沙丘顶部这种地面阻力较小的特殊环境[13], 进而导致拟步甲科成为沙丘顶部2种沙障材料林地的共有优势类群。由于沙丘顶部麦草方格内油蒿冠幅显著低于尼龙网方格, 且麦草方格内油蒿生长稀疏, 从而导致地面阻力较小, 有利于拟步甲科个体的移动和活动[13]。故拟步甲科个体数表现为麦草方格显著高于尼龙网方格。
本研究发现, 无论在沙丘底部还是顶部, 步甲科、叶甲科个体数均表现为麦草方格和尼龙网方格间无显著差异, 这与2种沙障材料林地间土壤含水量、电导率、容重、pH、粉粒、地温无显著差异有关。分析原因可能是地面节肢动物的个体数分布特征受到土壤含水量、电导率、容重、pH、粉粒、地温等环境因子的影响较大, 而受到土壤养分含量的影响较小[32]。在沙丘底部, 蠼螋科个体数表现为麦草方格和尼龙网方格间无显著差异, 但在沙丘顶部, 表现为麦草方格显著高于尼龙网方格。前者原因可能是沙丘底部土壤相对湿润[24], 蠼螋偏喜于在土壤水分条件较好的沙地环境中生存, 故蠼螋科个体数分布较多而成为优势类群, 且在2种沙障材料林地间无显著差异。后者原因可能是沙丘顶部麦草沙障可以作为枯落物, 为植食性动物提供食物来源的同时, 亦间接为捕食性蠼螋科个体提供了潜在的食物来源[5], 从而导致麦草方格林地蠼螋科个体数显著高于尼龙网方格。
本研究还发现, 无论是在沙丘底部还是顶部, 麦草方格内地面节肢动物个体数、类群数均显著高于尼龙网方格。分析原因主要是麦草沙障作为枯落物可以为地面节肢动物提供食物来源, 同时又具有保水性能, 微生境的改善可以为地面节肢动物活动提供生存条件[5], 故麦草沙障较尼龙网沙障更有利于地面节肢动物的生存。但地面节肢动物群落多样性表现为无论是在沙丘底部还是顶部, 地面节肢动物Shannon指数、优势度指数均表现为麦草方格和尼龙网方格间无显著差异。分析原因可能是经过5年的草方格固沙, 某一部位麦草被风蚀裸露后可能会加速邻近区域麦草的裸露, 麦草所起的保持水分和提高养分的作用随之减弱[19,27], 从而导致不同沙障林地地面节肢动物多样性分布无显著差异性, 同时也说明不同材料沙障对地面节肢动物多样性分布的影响有限。
3.3 地面节肢动物群落与环境因子的关系
本研究RDA和偏RDA分析结果表明, 花棒冠幅、花棒高度和柠条高度均对地面节肢动物个体数分布产生显著影响, 这与董炜华等[29]的研究结果相似。但是地面节肢动物个体数分布受到的土壤环境因子较小, 这与已有的研究结果不一致[30,32]。许多研究结果已经证实, 荒漠水分和全氮含量是影响荒漠节肢动物生长和分布的重要环境因子[33–34]。在景观及更大尺度上, 气候条件如温度和降水是影响地面节肢动物分布的环境因子[35–36]。刘任涛等[37]在干旱区不同林龄灌丛下研究表明, 土壤容重、pH和电导率对地面节肢动物群落结构影响显著。综合分析表明, 本研究所调查的土壤因子并不能反映环境条件对地面节肢动物群落分布的影响, 并且相关环境因子(包括植被和土壤因子)对地面节肢动物个体数分布的总贡献率仅为29.5%, 说明仍有其他重要因素如土壤全P、全K、有机质等因子对地面节肢动物个体数分布产生影响[37–38], 今后可能需要对土壤全K、全P和有机质含量等其他因素进行调查。
4 结论
(1)地面节肢动物群落优势类群包括步甲科、拟步甲科、蠼螋科和叶甲科, 反应了地面节肢动物对沙漠特殊生境的选择性和适应性。
(2)沙障材料对地面节肢动物群落组成、个体数和类群数分布均产生显著影响, 而且地面节肢动物群落组成分布亦受到沙丘微地形的影响。
(3)麦草方格较尼龙网方格可以更有效丰富地面节肢动物个体数, 但沙障材料对地面节肢动物多样性分布的影响较小。
(4)灌丛营造+草方格措施可以对固沙林地地面节肢动物个体数分布产生直接影响, 特别是花棒冠幅、花棒高度和柠条高度成为主要限制因素。
[1] 常海涛, 刘任涛, 刘佳楠, 等. 草方格造林固沙过程中土壤性质变化及分形特征—以腾格里沙漠东南缘为例[J]. 水土保持学报, 2018, 32(3): 58–65.
[2] LI Jinchang, LIU Haixia, SU Zhizhu, et al. Changes in wind activity from 1957 to 2011 and their possible influence on aeolian desertification in northern China[J]. Journal of Arid Land, 2015, 7(6): 755–764.
[3] 屈建军, 刘贤万, 雷加强, 等. 尼龙网栅栏防沙效应的风洞模拟实验[J]. 中国沙漠, 2001(3): 62–66.
[4] LI Xinrong, CHENG Yingwu, SU Yangui, et al. Effects of biological soil crust on desert insect diversity: evidence from the Tengger Desert of northern China[J]. Arid Land Research and Management, 2006, 20: 1–18.
[5] 胡英娣, 周洁. 用于草方格沙障的麦草方格保存研究[R]. 沙坡头沙漠研究试验站编, 1991.
[6] 马学喜, 王海峰, 李生宇, 等. 两种固沙方格沙障的防护效益及地形适应性对比[J]. 水土保持通报. 2015, 35(3): 344–349.
[7] 张立欣, 段玉玺, 王博, 等. 库布齐沙漠不同人工固沙灌木林土壤微生物量与土壤养分特征[J]. 应用生态学报, 2017, 28(12): 3871–3880.
[8] 屈建军, 凌裕泉, 俎瑞平, 等. 半隐蔽格状沙障的综合防护效益观测研究[J]. 中国沙漠, 2005, 25(3): 329–335.
[9] 董治宝, FRYREAR DONALD W, 高尚玉. 直立植物防沙措施粗糙特征的模拟实验[J]. 中国沙漠, 2000, 20(3): 260–263.
[10] 李瑞军. 棉秆沙障防风固沙效益比较[D]. 兰州: 甘肃农业大学, 2009: 47.
[11] 孙涛, 刘虎俊, 朱国庆, 等. 3种机械沙障防风固沙功能的时效性[J]. 水土保持学报, 2012, 26(4): 12–16.
[12] 王雨浩. 麦草、PLA及其混合沙障防风固沙效益研究[D]. 呼和浩特: 内蒙古师范大学, 2019: 78.
[13] 刘继亮, 赵文智, 李锋瑞, 等. 微地形变化对沙质荒漠拟步甲科昆虫分布的影响[J]. 干旱区研究, 2017, 34(6): 1388–1394.
[14] 刘任涛, 赵哈林, 赵学勇. 科尔沁沙地不同造林类型对土壤动物多样性的影响[J]. 应用生态学报, 2012, 23(4): 1104-1110.
[15] 王新谱, 杨贵军. 宁夏贺兰山昆虫[M]. 北京: 科学出版社, 2010.
[16] 尹文英. 中国土壤动物检索图鉴[M]. 北京: 科学出版社, 1998.
[17] 郑乐怡, 归鸿. 昆虫分类[M]. 南京: 南京师范大学出版社, 1999.
[18] 刘任涛, 郗伟华, 刘佳楠, 等. 沙地柠条(Caragana)灌丛微生境节肢动物群落特征[J]. 中国沙漠, 2018, 38(1): 117–125.
[19] 罗雅曦. 不同材料沙障固沙林地土壤—植被系统变化及其生态效应评价[D]. 银川: 宁夏大学, 2019: 40.
[20] 鲍士旦. 土壤农化分析3版[M]. 北京: 中国农业出版社, 2000.
[21] 赵娟,刘任涛,刘佳楠, 等.北方农牧交错带退耕还林与还草对地面节肢动物群落结构的影响[J].生态学报, 2019, 39(5): 1653–1663.
[22] 苏永中, 赵哈林, 张铜会. 几种灌木、半灌木对沙地土壤肥力影响机制的研究[J]. 应用生态学报, 2002, 13(7): 802–806.
[23] 鲍乾, 杨瑞, 李万红, 等. 喀斯特高原峡谷区不同恢复模式的土壤生态效应[J]. 水土保持学报, 2017, 31(3): 154–161+168.
[24] 高国雄. 毛乌素沙地东南缘人工植被结构与生态功能研究[D]. 北京: 北京林业大学, 2007: 181.
[25] 尹辉. 我国西南典型喀斯特峰丛洼地土壤理化特征研究[D]. 北京: 中国地质科学院, 2012: 143.
[26] 莫保儒, 蔡国军, 杨磊, 等. 半干旱黄土区成熟柠条林地土壤水分利用及平衡特征[J]. 生态学报, 2013, 33(13): 4011–4020.
[27] 王逸敏. 固沙沙障对植被、土壤和生物土壤结皮的影响[D]. 西安: 西北大学, 2018: 68.
[28] 张乐, 刘志民. 丘间低地生态过程研究进展[J]. 生态学杂志, 2007(7): 1101–1106.
[29] 董炜华, 李金霞, 刘继亮. 沙丘和丘间低地地面节肢动物群落比较研究[J]. 干旱区资源与环境, 2016, 30(1): 157–161.
[30] 娄巧哲, 徐养诚, 马吉宏, 等. 古尔班通古特沙漠南缘地表甲虫物种多样性及其与环境的关系[J]. 生物多样性.2011, 19(4): 441–452.
[31] 刘淑蓉. 天津八仙山鳞翅目幼虫多样性初步研究[D]. 天津: 南开大学, 2011: 90.
[32] 常海涛,刘佳楠,赵娟,等. 乌拉特荒漠草原红砂灌丛林地地面节肢动物群落季节分布特征[J]. 生态学报, 2020 (9): 1–10.
[33] 赵文智, 刘鹄. 干旱、半干旱环境降水脉动对生态系统的影响[J]. 应用生态学报, 2011, 22(1): 243–249.
[34] 赵哈林, 刘任涛, 赵学勇, 等. 旱作农田改为水浇地对沙质土壤节肢动物群落的影响[J]. 干旱区资源与环境, 2014, 28(1): 9–14.
[35] LI Fengrui, LIU Jiliang, LIU Changan, et al. Shrubs and species identity effects on the distribution and diversity of ground - dwelling arthropods in a Gobidesert[J]. Journal of Insect Conservation, 2013: 1–13.
[36] ZHAO Halin, LIU Rentao. The " bug island" effect of shrubs and its formation mechanism in Horqin Sand Land, Inner Mongolia[J]. Catena, 2013, 105: 69–74.
[37] 刘任涛, 朱凡, 柴永青. 干旱区不同年龄灌丛斑块地面节肢动物的聚集效应[J]. 应用生态学报, 2014, 25(1): 228–236.
[38] 刘继亮,李锋瑞,牛瑞雪,等.黑河中游干旱绿洲土壤盐渍化对大型土壤动物群落的影响[J]. 应用生态学报. 2012, 23(6): 1551–1561.
Effects of shrub revegetation within different sand barrier materials on the community structure of ground-active arthropods in desert ecosystems
FENG Yonghong1, GOU Qi2,3, LIU Rentao2,3,*, LUO Yaxi2, CHANG Haitao2, ZHANG Jing1, JIANG Jiayu2,3
1. College of Agriculture, Ningxia University, Yinchuan 750021, China 2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education, Ningxia University, Yinchuan 750021, China 3. College of Ecology and Environment, Ningxia University, Yinchuan 750021, China
The aim of this study was to probe into the effects of different sand barrier materials on arthropod community structure in afforested shrub plantations in desert ecosystems. The composition and diversity distribution of ground-active arthropod communities were investigated at the bottom and top of sand dunes within checkerboards of wheat straw and nylonin spring, summer and autumn of 2018 in the southeast edge of Tengger Desert. The results showed that: (1) At dune bottom, there were three dominant groups including Labiduridae, Carabidae and Chrysomelidae observed in the microhabitats within checkboards of two materials. In addition, there were two groups including Tenebrionidae and Coleoptera larvae in the microhabitats within wheat-straw checkerboard, and one group including Lepidoptera larvae in the microhabitats within nylon checkerboard. However, at dune top, there were similar dominant groups in the microhabitats between the two materials including Labiduridae, Carabidae, Chrysomelidae, and Tenebrionidae. (2) There was no significant difference in the abundance of Carabidae and Chrysomelidae between the microhabitats at dune bottom and top. At dune bottom, the abundance of Tenebrionidae was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard, and the abundance of Labiduridae was found to indicate no significant difference between the dune microhabitats within two materials; at dune top, there was no significant difference in the abundance of Tenebrionidae between the microhabitats within two materials, and the abundance of Labiduridae was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard. (3) At dune bottom and top, the total abundance and group richness of ground-active arthropods was significantly higher in the microhabitats within wheat-straw checkerboard than within nylon checkerboard, but there was no significant difference in Shannon index and dominance index in the microhabitats between the two materials. (4) The results of RDA and partial RDA analysis showed that the height and crown width of, and crown width ofwere the main factors driving the differences of abundance distribution of ground-active arthropods between different microhabitats. It was concluded that sand barrier materials indicated significant effects on the composition, abundance and group richness distribution of ground-active arthropod community, and their composition and abundance distribution were also affected by the dune microhabitats, but there was little effect of sand barrier materials on the ground-active arthropod diversity distribution in Tengger Desert ecosystem.
Tengger Desert; sand barrier material; dune microhabitats; ground-active arthropod; community structure
冯永宏, 苟琪, 刘任涛, 等. 不同沙障材料对固沙林地面节肢动物群落结构分布的影响[J]. 生态科学, 2022, 41(5): 35–45.
FENG Yonghong, GOU Qi, LIU Rentao, et al. Effects of shrub revegetation within different sand barrier materials on the community structure of ground-active arthropods in desert ecosystems[J]. Ecological Science, 2022, 41(5): 35–45.
10.14108/j.cnki.1008-8873.2022.05.005
958.15+5
A
1008-8873(2022)05-035-11
2020-08-19;
2020-09-27
宁夏自然科学基金(2020AAC0214, 2021AAC03047); 宁夏重点研发计划项目(2021BEG03007); 国家自然科学基金(41867005)
冯永宏(1996—), 男, 陕西榆林人, 硕士研究生; 主要从事水土保持与荒漠化防治研究, E-mail: 13772928515@163.com
刘任涛, 男, 博士, 研究员, 主要从事防沙治沙与生态恢复相关研究, E-mail: nxuliu2012@126.com
