基于高通量测序技术的宁夏贺兰山东麓产区酿酒葡萄微生物多样性的研究
2022-08-04薛蓓于佳俊杨帆卢灏泽石俊张晓蒙马文瑞张晶晶陈璐王妍凌薛洁
薛蓓,于佳俊,杨帆,,卢灏泽,,石俊,张晓蒙,马文瑞,张晶晶,3,陈璐,3,王妍凌,3,薛洁*
1(西藏农牧学院 食品科学学院,西藏 林芝,860000)2(中国食品发酵工业研究院,北京,100015)3(新疆农业大学 食品科学与药学学院,新疆 乌鲁木齐,830052)
宁夏贺兰山东麓葡萄酒产区是我国最适宜葡萄种植的产区之一,具有发展葡萄产业得天独厚的资源禀赋和区位优势,自古以来是中国主要的酿酒葡萄及葡萄酒产地,近两年由于葡萄产业化及龙头企业的发展,宁夏葡萄酒产量逐年提高,工业水平有了明显的发展,葡萄酒已经成为自治区经济转型和支柱性优势特色产业。
葡萄酒的酿造是由多种微生物共同作用完成的。NICHOLAS等[1]首次提出葡萄酒的“微生物风土”概念,即葡萄酒微生物的群落结构特征可以表现出产区特异性的特点。通过对不同产区、不同葡萄品种、不同年份葡萄酒微生物多样性的检测,以及相对应的葡萄酒风格之间的比较,越来越多的研究认为微生物是葡萄酒产区质量和感官风格的决定性因素之一。
目前分析酿酒葡萄中微生物多样性的方法主要有PCR技术,其中包括限制片段长度多态性(restriction fragment length polymorphism,RFLP)和随机扩增多态性DNA分析(random amplified polymorphic DNA,RAPD),但是这些方法不仅耗时,而且结果也不太准确。高通量测序技术的出现完全避开了这些缺点,该技术不仅测序周期短,测出的数据量大,而且结果准确率高、成本低,所以该方法是目前最适合分析微生物多样性的技术。张二豪等[2]利用高通量测序技术发现西藏芒康和林芝地区葡萄果皮上优势菌属分别为有孢汉逊酵母属(Hanseniaspora)和枝孢属(Cladosporium)。张俊杰等[3]利用高通量测序技术在市售夏黑无核葡萄果皮上检测出致病菌志贺氏菌属(ShigellaCastellani),对市场售卖果蔬前的检疫工作有一定的借鉴意义。BOKULICH等[4]利用高通量技术发现美国加利福尼亚两个不同产区的霞多丽中的葡糖杆菌属(Gluconobacter),在葡萄酒发酵过程中可以促进合成脂类物质,从而提升葡萄酒的品质。
本文利用高通量测序技术分析贺兰山东麓产区中西夏区、永宁县、青铜峡,石嘴山4个葡萄园区中酿酒葡萄果皮上的微生物组成,揭示产区微生物的多样性,为筛选产区优质的微生物以及酿酒微生物菌库的建立提供了理论基础。
1 材料与方法
1.1 材料与试剂
样品来自中国宁夏贺兰山东麓葡萄酒产区中西夏区(XXQ)、永宁县(YNX)、青铜峡(QTX)和石嘴山(SZS)4个产区中的葡萄,各产区地址如图1所示,于2019年9月采样,样品如表1 所示。
琼脂糖,西班牙Biowest Agarose公司;NGS-ITS1 copy1 Barcode1-70、NGS-16s V4 copy1 Barcode1-70,生工生物工程(上海)股份有限公司;Gold View I核酸染料,北京中生瑞泰科技有限公司;DL 2000 Marker,美国Axygen公司;所有提取用无机、有机溶剂均为国产分析纯。
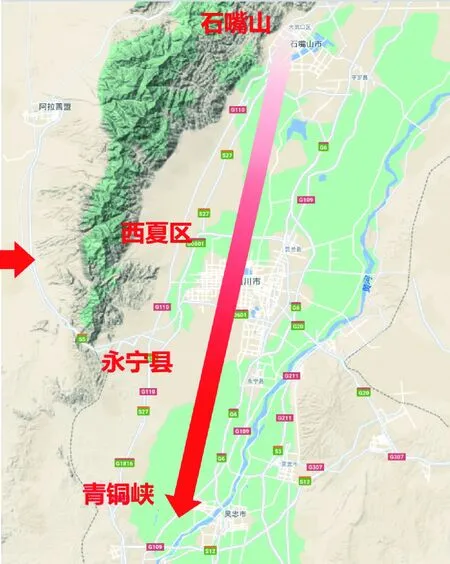
图1 产区地图Fig.1 Production area map
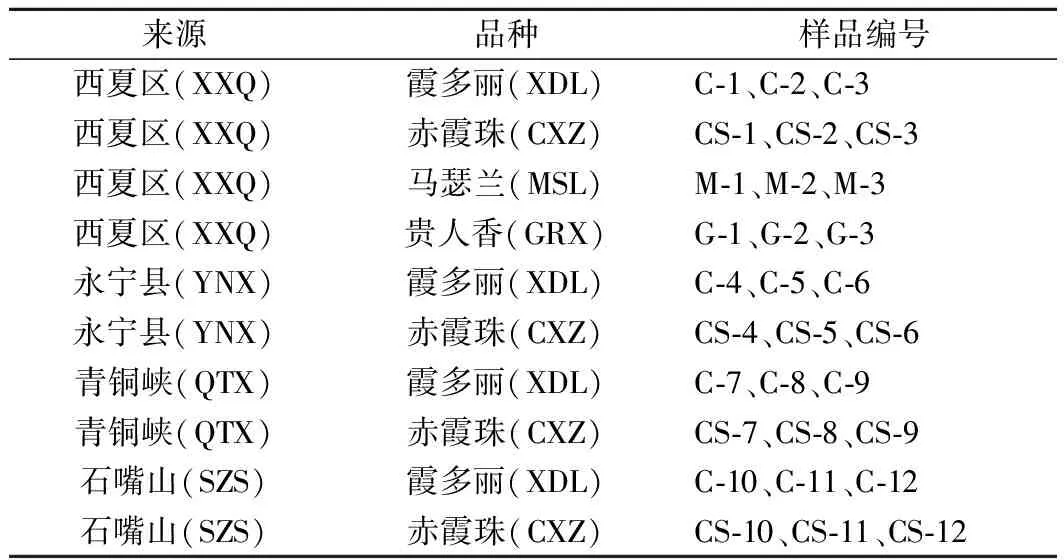
表1 实验样品Table 1 Experimental samples
1.2 仪器与设备
Illumina Misq测序平台,美国Illumina公司;5810R离心机,德国 Eppendorf 公司;PCR仪,美国伯乐公司;立式电热压力蒸汽灭菌锅,上海申安医疗器械厂。
1.3 采样方法
选择各葡萄园中有代表性的葡萄树,用消毒后的剪刀从葡萄树的上、中、下3个地方采集果实,采集后放入无菌采样袋中,再将采样袋放入装有液氮的采样箱中带回实验室,-20 ℃低温保藏。将同产区同品种样品混合制成聚合样本后用于后续试验。
1.4 葡萄细菌和真菌的分离[5-6]
(1)取适量葡萄,在无菌条件进行果皮分离,分离后在液氮条件下研磨果皮,称取4 g果皮粉放入无菌的50 mL离心管A1中。
(2)加入6 mL TE Buffer缓冲溶液,配平后5 000×g离心6 min,然后将上清液转移到另一个无菌的50 mL离心管A2中。
(3)重复步骤2两次。
(4)向A1管中加入6 mL PBS Buffer 缓冲溶液,振荡混匀后进行离心,5 000×g离心6 min,将上清液在无菌条件下加入到A2管中。
(5)配平后进行离心,9 000×g离心10 min后舍弃上清液,沉淀物放于-20 ℃进行保存。
1.5 DNA的提取
采用Fast DNA SPIN Kit for Soil试剂盒(MP)进行提取。
1.6 样品分析
细菌PCR所用的引物为16S rDNA V4:16S 515F (5′-GTGCCAGCMGCCGCGGTAA-3′);16S 806R(5′-GGACTACHVGGGTWTCTAAT-3′)。真菌PCR所用的引物为引物ITS1:ITS 1F(5′-CTTGGTCATTTAGAGGAGTAA-3′);ITS 1R(5′-GCTGCGTTCTTCATCGATGC-3′)。PCR反应体系为:dNTP Mixture 4 μL,10×PCR Buffer(Mg2+plus)5 μL,正反向引物各1 μL,样品DNA 5 μL,ExTaq酶0.25 μL,ddH2O补足至50 μL,充分混匀。真菌PCR扩增程序:98 ℃预变性3 min;98 ℃变性45 s,53 ℃退火30 s,72 ℃延伸45 s,反应 35 个循环;72 ℃延伸8 min。-20 ℃保存。细菌PCR扩增程序:95 ℃预变性3 min;95 ℃变性45 s, 50 ℃ 退火30 s,72 ℃延伸1 min,反应35个循环;72 ℃延伸5 min。-20 ℃保存[7]。
取PCR产物10 μL用2%琼脂糖凝胶电泳进行检测,将检测合格样品送至北京诺禾致源科技股份有限公司进行进行真菌ITS ITS1、细菌16S rDNA V4区域文库构建,最后用Illumina Misq 进行测序分析[8]。
1.7 数据预处理
将测序完成后的数据进行预处理,过滤去除引物和接头序列、非特异性扩增序列及嵌合体,使得最后用于操作分类单元(operational taxonomic units,OTU)分析的数据有效可靠。
1.8 数据分析
1.8.1 OTU聚类分析
OTU是指通过一定的距离度量方法计算两两不同序列之间的距离度量或相似性,继而设置特定的分类阈值,获得同一阈值下的距离矩阵,进行聚类操作,形成不同的分类单元。用Qiime软件对4个不同产区的酿酒葡萄进行OTU分析,将相似度≥97%的聚为一类,采用贝叶斯算法获得每个OTU的分类学信息,利用R软件绘制韦恩图后进行分析。
1.8.2 Alpha分析
Alpha分析能够反映微生物群落的丰度和多样性,主要包括 Chao1、多样性指数Shannon 和 Coverage。可以通过Coverage值检测出测序结果有无代表性,其数值越高,结果越具代表性。
1.8.3 微生物多样性分析
通过对OTU数据进行分析,利用柱状图对样品中的细菌和真菌进行统计分析并确定优势菌属;利用主坐标分析(principal coordinate analysis,PCoA)和非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)聚类树分析样本间相似度;利用韦恩图来观察4个不同产区中酿酒葡萄样品中所具有的相同的菌属,通过上述分析方法来综合分析样品中微生物的多样性。
2 结果与分析
2.1 微生物群落多样性分析
微生物的Alpha多样性指数有Shannon和Simpson两种。Shannon值越大,群落多样性越高;Coverage指各样本文库的覆盖率,其数值越高,则样本中序列被测出的概率越高,而没有被测出的概率越低。从表2中得出,在细菌方面,4个不同产区微生物多样性不仅受酿酒葡萄品种的影响,而且也受小产区气候、土壤条件的影响,从细菌多样性来讲,4个产区多样性指数从高到低依次为SZS、YNX、XXQ、QTX;品种间细菌多样性指数从高到低依次为霞多丽、贵人香、马瑟兰和赤霞珠。
从图2-b可以看出,在真菌属水平上,XXQ产区的样品中真菌最为丰富的菌属为拟点霉属(Phomopsis)、未鉴定出的菌属(unidentified),较为丰富的菌属为Rhodosporidiobolus;在YNX产区的样品中最为丰富的菌属为曲霉菌属(Aspergillus)、链格孢属(Alternaria),其次较为丰富的是短梗霉属(Aureobasidium)和Naganishia;在QTX产区的样品中最为丰富的菌属为Papiliotrema、有孢汉逊酵母属(Hanseniaspora)、Curvibasidium,其次较为丰富的是线黑粉酵母属(Filobasidium);在SZS产区的样品中最为丰富的菌属赤霉菌属(Gibberella)、附球霉属(Epicoccum),其次较为丰富的是Cladosporium和亚隔孢壳属(Didymella)。
因此,在属水平上,无论是细菌多样性,还是真菌多样性,4个产区间都存在明显差异,这或许就是不同产区葡萄酒风格差异的原因所在。
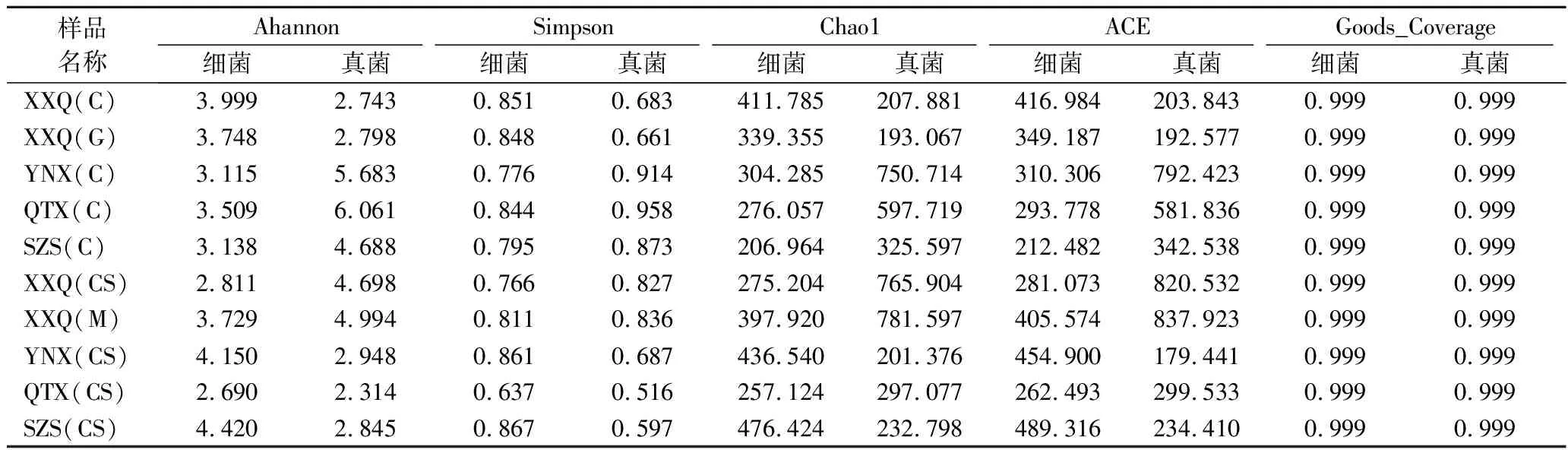
表2 样品Alpha多样性指数表Table 2 Sample Alpha diversity indexes table
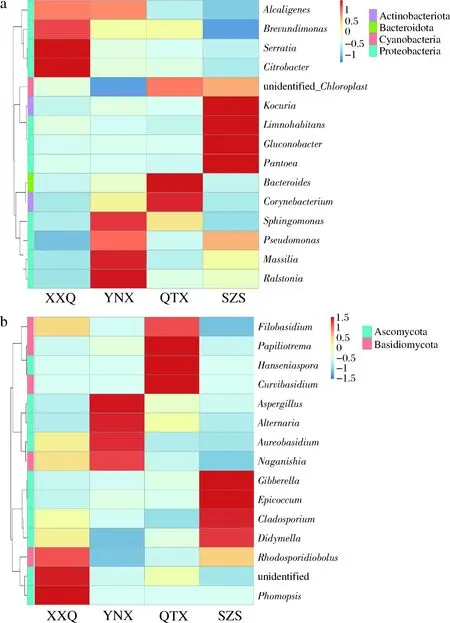
a-样品细菌相对丰度热图;b-样品真菌相对丰度热图图2 样品相对丰度热度图Fig.2 Heat map of samples relative abundance
2.2 微生物丰度分析
由于检测出样品中含有多种微生物,有些微生物含量很少,与丰度高的微生物在同一图中不能明显的突出,所以把丰度>1%的物种作为划分依据,选择排名前15的优势菌属,以相对丰度为纵坐标,绘制柱形图。
2.2.1 不同产区赤霞珠样品真菌丰度分析
从图3可以看出,在属水平上,赤霞珠样品中最为丰富的菌属为Alternaria、Cladosporium、镰刀菌属(Fusarium)、毛壳菌属(Chaetomium)、Didymella。不同产区微生物多样性有差异,但也有相似之处。XXQ和SZS产区虽然丰度最高的是Cladosporium,但是在SZS产区,丰度较高的Chaetomium、Fusarium和Alternaria所占比较非常接近,而XXQ产区Alternaria微生物含量相对较多,而Chaetomium、Fusarium微生物含量甚微。YNX和QTX产区微生物丰度比较相似,丰度最高的Alternaria,其次是Cladosporium。
从图4可以看出,XXQ、QTX、SZS、YNX葡萄园中赤霞珠样品所含的真菌OTU分别为280个、218个、208个、388个。XXQ和YNX产区拥有219种相同的微生物种类,这与两个产区在贺兰山葡萄酒产区的地理位置相近有关;石嘴山产区由于位于整个产区的最北端,因此这或许是产区赤霞珠果皮上所含的真菌OUT值最小的原因所在;在4个小产区中,QTX产区由于位于贺兰山东麓产区的南端,距离SZS产区较远,两个产区的微生物的相似度较低,这也说明了不同产区微生物的多样性受地理条件的影响。
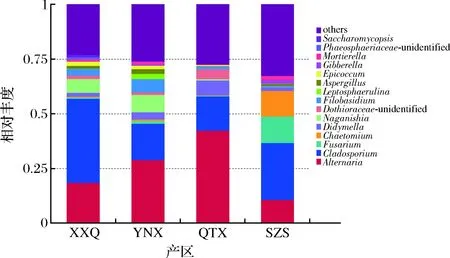
图3 真菌属水平相对丰度柱状图Fig.3 Histogram of relative abundance of fungi
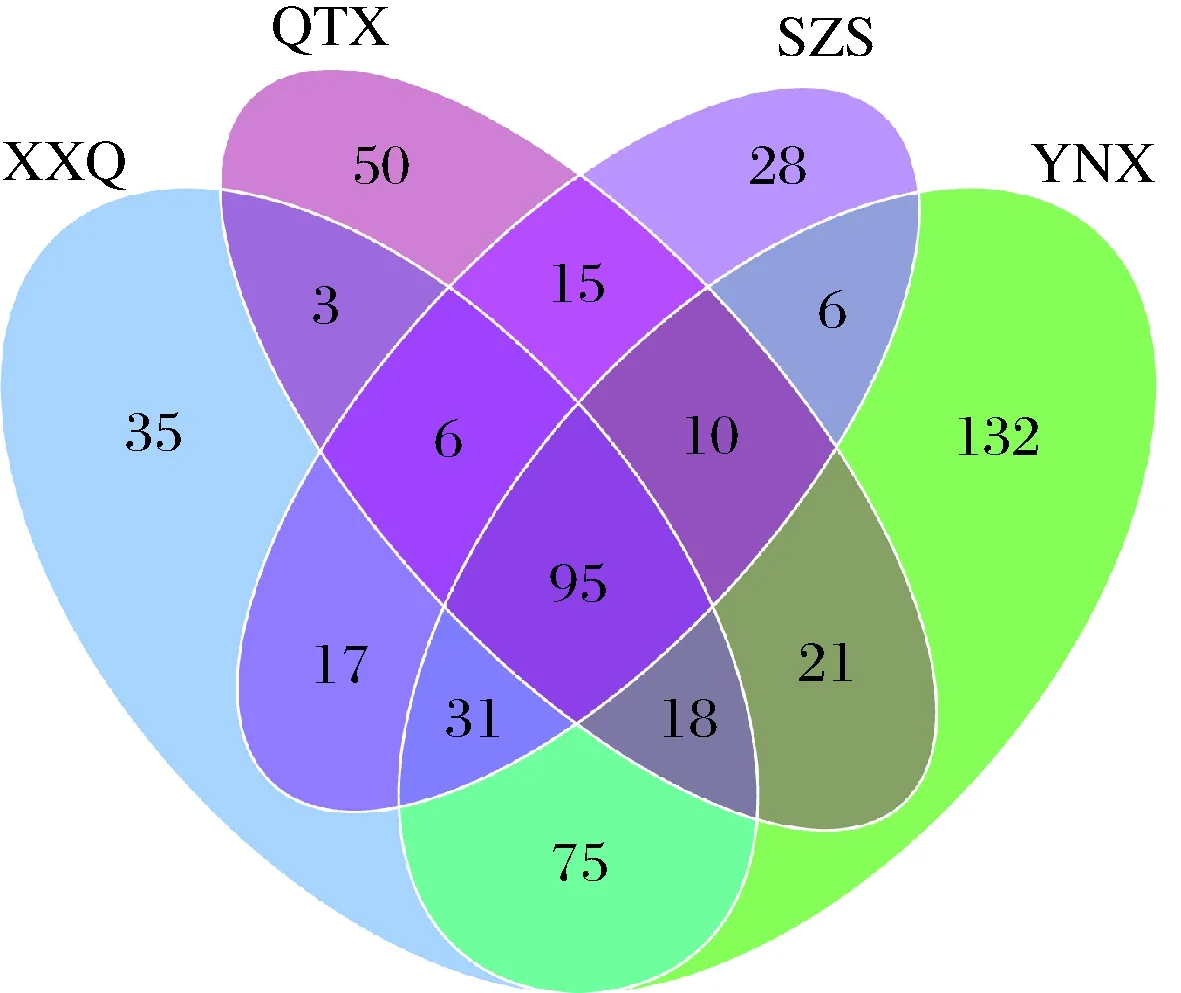
图4 真菌群落维恩图Fig.4 Venn diagram of fungal community
2.2.2 不同产区赤霞珠样品细菌丰度分析
在减少腐败存量、遏制腐败增量的努力中,我国多年来经历了“运动反腐”、“权力反腐”、“制度反腐”、“体系反腐”等多种模式,[1]却陷入了腐败存量与增量此起彼伏的怪圈。我们要承认腐败存在的历史必然性及腐败难以彻底消失的事实、最大限度提高“减存遏增”的效率,要在保证公共治理的必要权力和维持经济社会发展前进的前提下,寻找腐败存量与增量的平衡点,将腐败存量与增量控制在符合经济和社会可接受的最低水平。为达到这一目的,本文试图探索一种有效控制腐败存量与增量的平衡机制,以期为腐败存量与增量问题的解决提供方向。
从图5可以看出,在属水平上,赤霞珠样品中的优势菌属为罗尔斯通氏菌属(Ralstonia)、unidentified_Chloroplast、Massilia、Gluconobacter、鞘氨醇单胞菌属(Sphingomonas)。在XXQ、YNX、SZS样品里丰度最高的菌属是Ralstonia,其次都是unidentified_Chloroplast;Ralstonia是重要的植物致病菌,可在营养缺乏的环境中生存。而在QXT样品里丰度最高的菌属是unidentified_Chloroplast,其次是Ralstonia。Massilia在XXQ、QTX、SZS样品里含量很低甚至没有检测出来;Gluconobacter在XXQ、YNX、SZS样品里含量很低甚至没有检测出来;Sphingomonas只在YNX中存在,在其他3个样品中含量很少甚至没有检测出来,Sphingomonas对芳香化合物有极为广泛的代谢能力,并且该菌属某些菌种能够合成有价值的胞外生物高聚物,可带给葡萄酒特殊的风味。
4个不同产区在属水平上优势菌属不仅存在差异,而且微生物种类间也存在差异,图6结果显示,XXQ、QTX、SZS、YNX葡萄园中赤霞珠样品所含的细菌OTU分别为138个、311个、225个、148个。但是与真菌不同的是,虽然QTX和SZS相聚较远,但两个产区拥有相同的细菌种类却最多,达到了82种,说明与真菌相比,赤霞珠品种中细菌的分布受地理位置的影响较小。
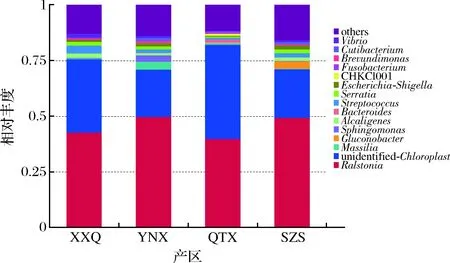
图5 细菌属水平相对丰度柱状图Fig.5 Histogram of relative abundance of bacteria
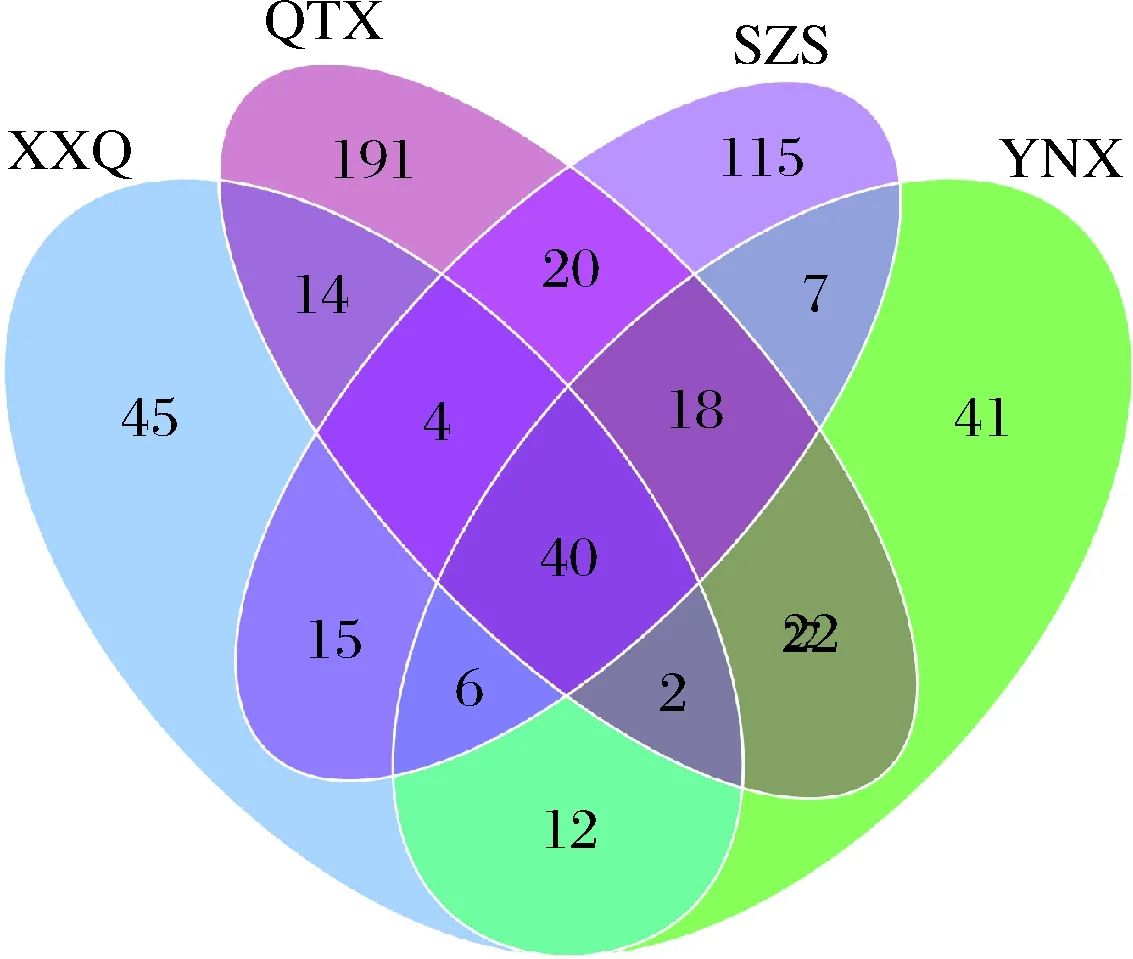
图6 细菌群落维恩图Fig.6 Venn diagram of bacterial community
2.2.3 不同产区霞多丽样品真菌丰度分析
与赤霞珠品种相比,霞多丽品种真菌的分布既有差异,也有相同之处。图7结果显示,在属水平上,霞多丽样品中最为丰富的菌属为Alternaria、Cladosporium、Naganshia、Hanseniaspora、Aspergillus、Filobasidium和Didymella。在XXQ、YNX、SZS样品里丰度最高的菌属是Aspergillus,其次都是Cladosporium;在SZS样品里丰度最高的菌属是Cladosporium,其次是Aspergillus。Hanseniaspora在QYX样品里丰富度较高,但是在其他3个样品里含量很少甚至没有检测出来,Hanseniaspora对葡萄采后病害具有防治作用,可抑制灰霉病、炭疽病等病害[9];而Didymella出现在所有样品中,但在YNX中丰度较小,Didymella中有许多植物致病菌,所以推测QYX产区在本年份病害会比较轻。
从图8可以看出,XXQ、QTX、SZS、YNX葡萄园中霞多丽样品所含的真菌OTU分别为267、152、98、325个。其中XXQ和YNX产区距离最近,拥有相同的种类也最多,达到了113种;QTX和SZS产区相聚较远,相同的真菌种类也较小,为47种;YNX产区的霞多丽果皮上所含的真菌OTU值最大,而SZS产区的霞多丽果皮上所含的真菌OUT值最小,该结果与赤霞珠品种分析结果完全一致。
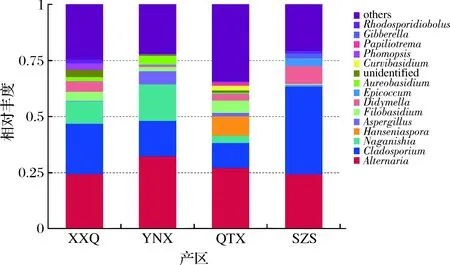
图7 真菌属水平相对丰度柱状图Fig.7 Histogram of relative abundance of fungi
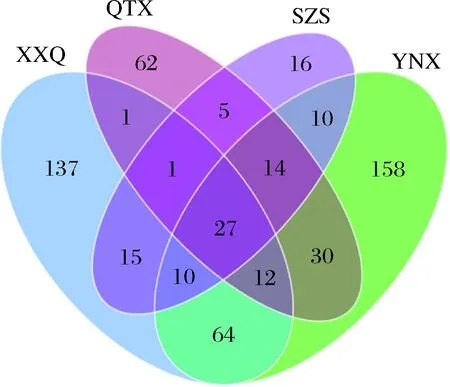
图8 真菌群落维恩图Fig.8 Venn diagram of fungal community
2.2.4 不同产区霞多丽样品细菌丰度分析
从图9可以看出,在属水平上,霞多丽样品中最为丰富的菌属为Serratia、Ralstonia、Pantoea、unidentified_Chloroplast、假单胞菌属(Pseudomonas)。所有产区中都表现出较高的微生物多样性。XXQ中丰度最高的菌属是Serratia;YNX中丰度最高的菌属是Ralstonia;QYX中丰度最高的菌属是Ralstonia;SZS中丰度最高的菌属是unidentified_Chloroplast。虽然最为丰富的5种菌属在所有样品中都存在,但是在每个产区中的丰度却存在差异。
4个产区不仅细菌丰度存在差异,而且微生物的种类也不同,图10结果显示,XXQ、QTX、SZS、YNX葡萄园中霞多丽样品所含的细菌OTU分别为700、472、600、892个,产区间差异非常显著,YNX产区的霞多丽果皮上所含的细菌OTU值最大,而SZS产区的霞多丽果皮上所含的细菌OUT值最小。而且与真菌种类分布具有相同的规律,产区距离越近,相同的微生物种类也最多,XXQ和YNX产区有相同种类516种,而QTX和SZS产区有相同种类只有209种。
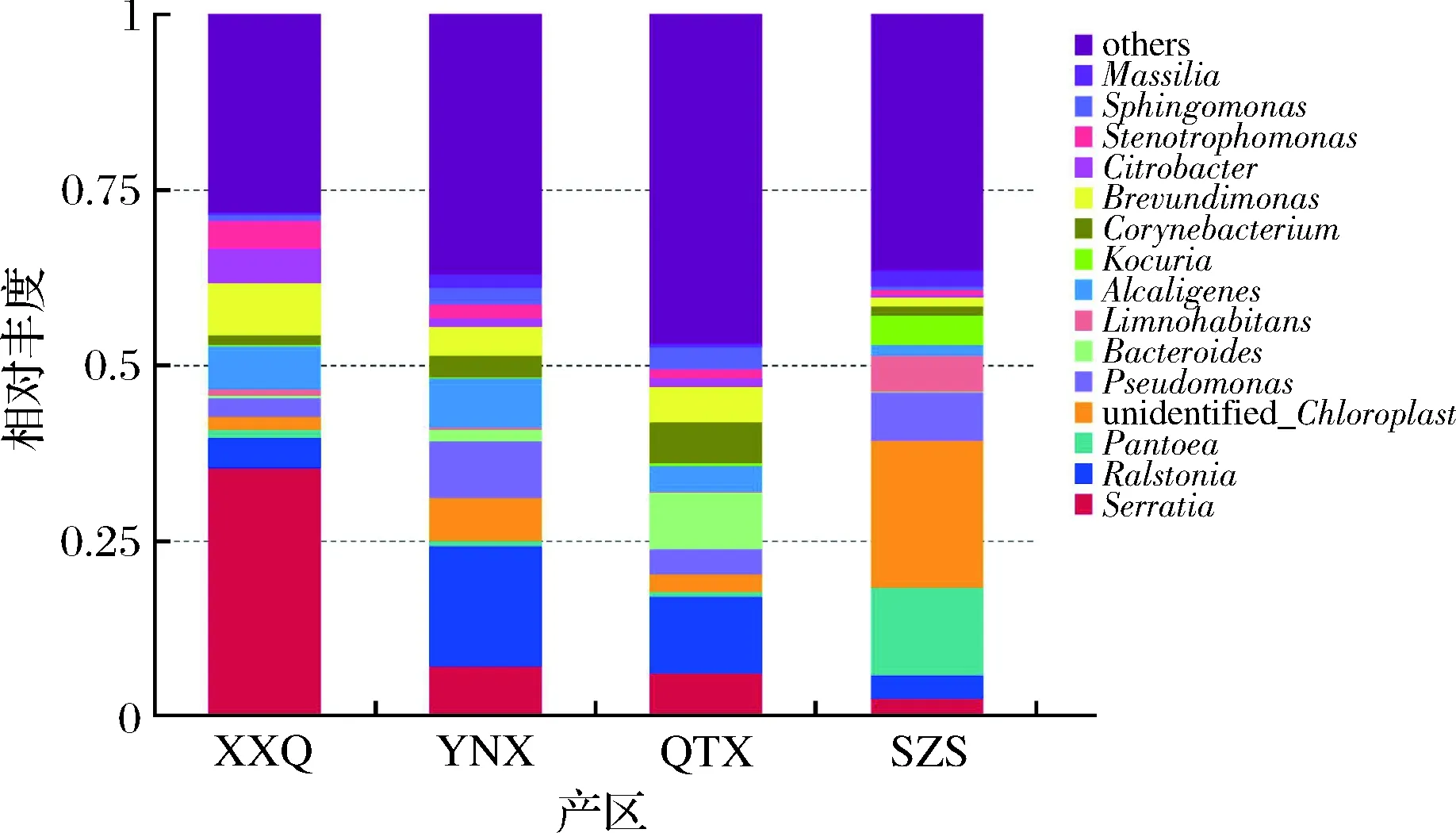
图9 细菌属水平相对丰度柱状图Fig.9 Histogram of relative abundance of bacteria
2.2.5 同产区不同酿酒葡萄品种真菌丰度分析
为了比较同产区不同酿酒葡萄品种微生物多样性的差异,本研究比较了西夏区4个品种间微生物多样性差异,结果如图11、图13所示。
从图11可以看出,4个不同品种,由于采收期存在差异,所以在属水平上的真菌丰度也不同。霞多丽品种最为丰富的真菌是Cladosporium、Alternaria、Naganishia、Didymella和Filobasidium,而贵人香品种最为丰富的真菌是Alternaria、Cladosporium、unidentified、Megnaporthe和Didymella;赤霞珠和马瑟兰品种优势菌属一致,为Cladosporium、Alternaria,但除优势菌属外,赤霞珠中占比较高的是Naganishia、Didymella和Epicoccum,马瑟兰中占比较高的是unidentified、Epicoccum和Rhodosporidiobolus。
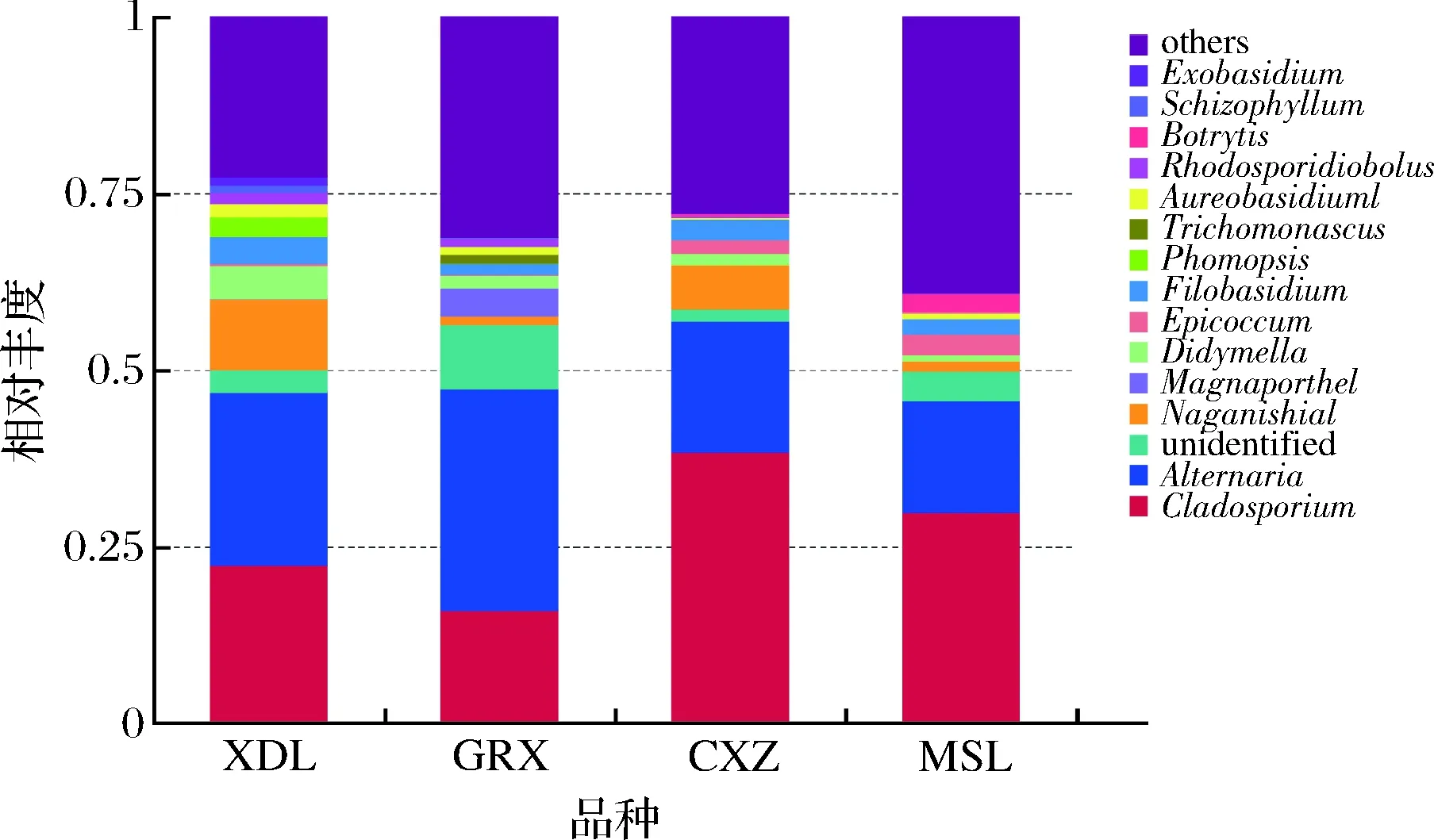
图11 真菌属水平相对丰度柱状图Fig.11 Histogram of relative abundance of fungi
对比分析4个不同酿酒葡萄品种真菌的种类,结果如图12所示,酿酒葡萄采收期越晚,葡萄果皮上的微生物种类也越多,GRX、XDL、CXZ、MSL 4个品种中所含的真菌OTU分别为210、267、280、366个。XDL和GRX有相同种类91种,CXZ和MSL有相同种类210种,而红葡萄品种和白葡萄品种间拥有相同类别的真菌种类也较小,XDL和CXZ、XDL和MSL有相同种类均为56种。
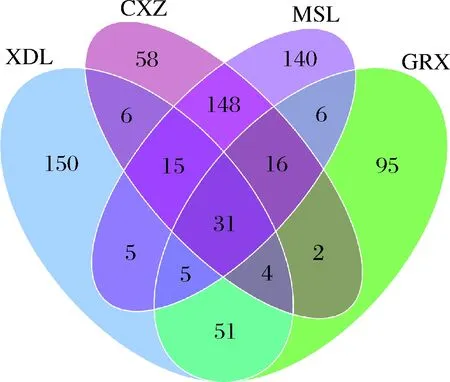
图12 真菌群落维恩图Fig.12 Venn diagram of fungal community
细菌的丰度分布4个品种间差异较大,如图13所示。在属水平上,霞多丽中最丰富的是Serratia,贵人香中最丰富的是Bacteroides,而赤霞珠和马瑟兰中最为丰富的都是罗尔斯通氏菌属(Ralstonia)。白葡萄品种间差异较大,除优势菌种外,在霞多丽中占主要比例的还有产碱菌属 (Alcaligenes)、Brevundimonas、Citrobacter和Stenotrophomonas。而在贵人香中占主要比例为罗尔斯通氏菌属(Ralstonia)、Alcaligenes、unidentified_Chloroplast和Acinetobacter。红葡萄品种中微生物的多样性差异相对较小,除了优势菌种外,在两种葡萄中占主要比例的还有unidentified_Chloroplast、Acinetobacter和Pseudomonas。
图14结果表明,随着采收期的推迟,葡萄果皮上细菌的种类逐渐减少,XDL、GRX、MSL、CXZ样品中所含的细菌OTU分别为700、567、201、138个,因此细菌的分布虽然与地理位置关系较小,但是与葡萄采收期却密切相关,这也是白葡萄果皮上所含的细菌OTU值比红葡萄品种大的原因所在。
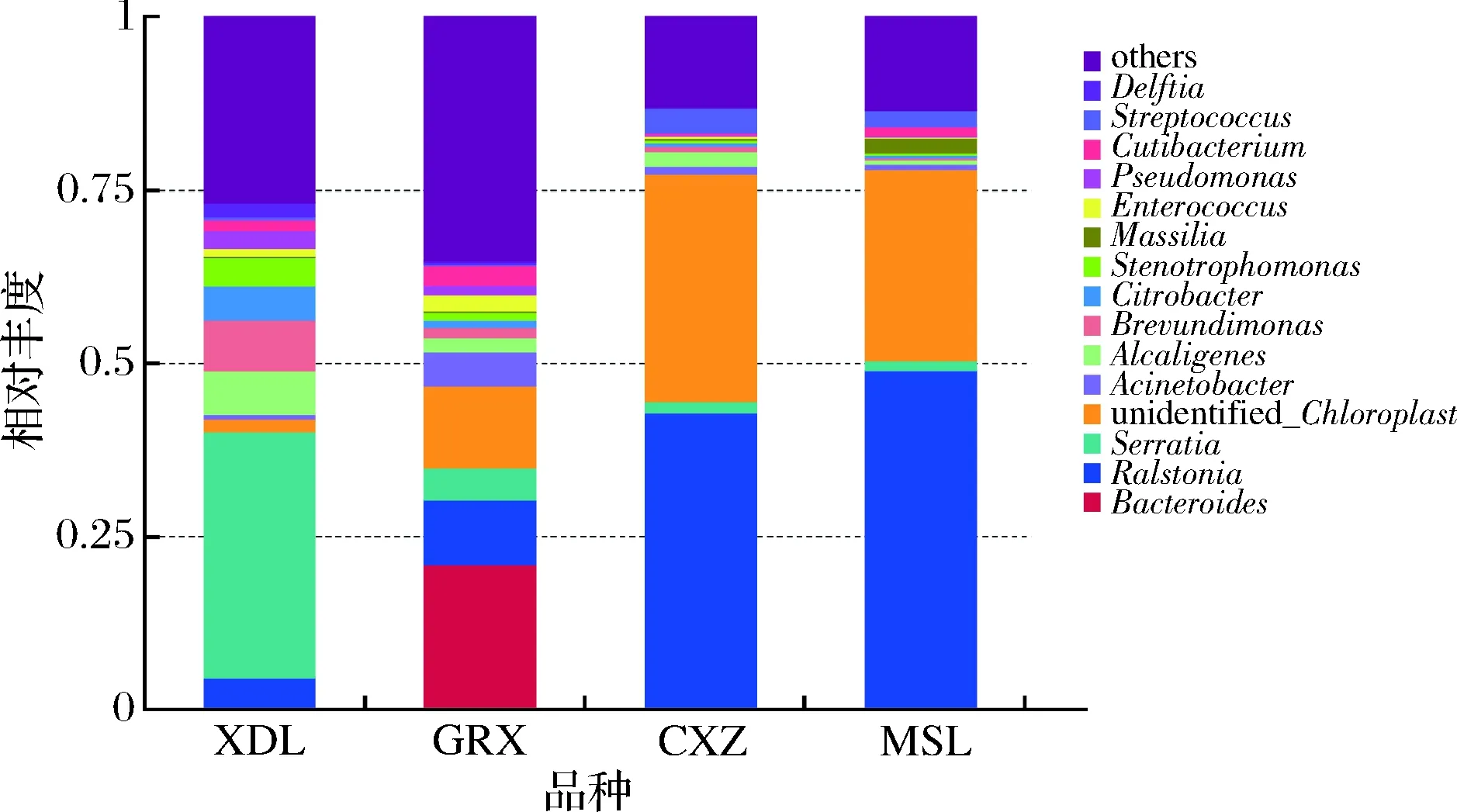
图13 细菌属水平相对丰度柱状图Fig.13 Histogram of relative abundance of bacteria
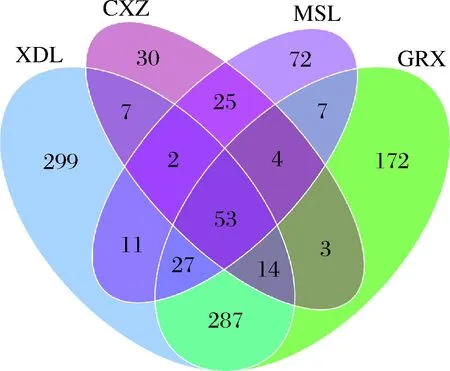
图14 细菌群落维恩图Fig.14 Venn diagram of bacterial community
2.3 微生物群落结构相似性分析
从图15-a可以看出,当PC1为41.07%,PC2为18.4%时,XXQ马瑟兰与XXQ赤霞珠样品在PCA图中的位置集中,距离紧密,表示这两种样品间微生物相似度较高。SZS赤霞珠样品在PCA图中比较分散,距离较远,表示样品间微生物相似度较低,其原因可能是SZS地区的气候夏天热而短促、天气经常变化导致样品间真菌微生物的不同。从图15-b中可以看出在0.68时SZS霞多丽样品出现分支,说明此葡萄样品间的微生物差异性较小;SZS赤霞珠在0.04时出现分支,说明此葡萄样品间的微生物差异性较大。
从图16-a可以看出,当PC1为46.27%,PC2为18.85%时,XXQ霞多丽和QTX霞多丽样品在PCA图中的位置集中,距离紧密,表示这两种样品间微生物相似度较高,其他样品在PCA图中比较分散,距离较远,表示样品间微生物相似度较低。从图16-b 中可以看出在0.32时XXQ和SZS产区霞多丽样品出现分支,说明此葡萄样品间的微生物差异性较小。样品间微生物相似度较低,可能与产区的空间距离、气候特征有直接关系。
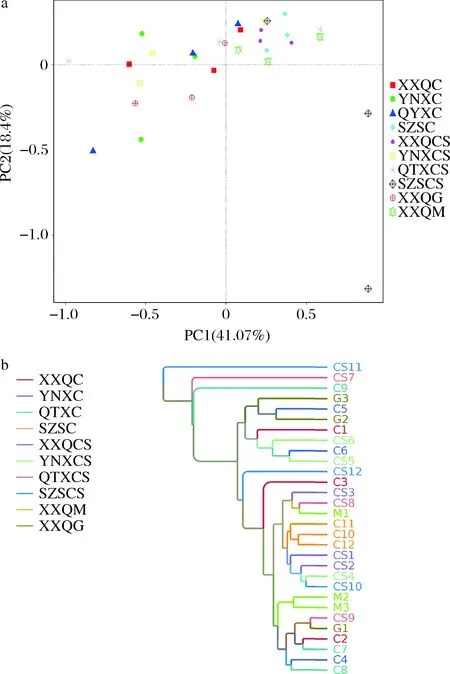
a-样品真菌群落PCoA;b-UPGMA聚类分析图图15 样品真菌β多样性分析Fig.15 Fungal β diversity analysis of samples注:XXQC(西夏区霞多丽)、YNXC(永宁县霞多丽)、QTXC(青铜峡霞多丽)、SZSC(石嘴山霞多丽)、XXQCS(西夏区赤霞珠)、YNXCS(永宁县赤霞珠)、QTXCS(青铜峡赤霞珠)、SZSCS(石嘴山赤霞珠)、XXQG(西夏区贵人香)、XXQM(西夏区马瑟兰)(下同)
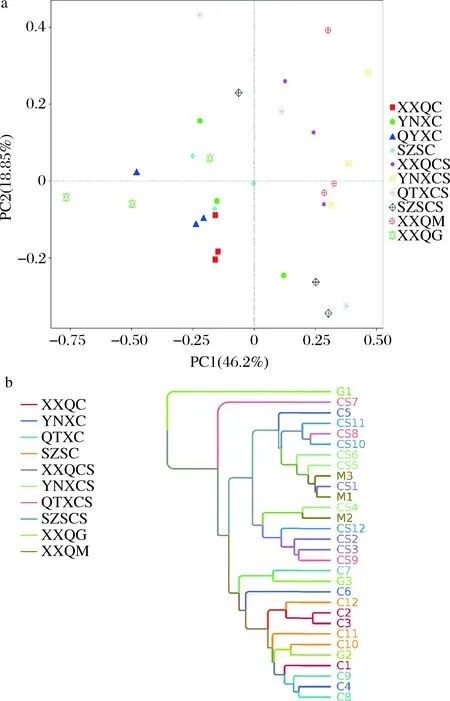
a-样品细菌群落PCoA;b-UPGMA聚类分析图图16 样品细菌β多样性分析Fig.16 Bacterial β diversity analysis of samples
3 讨论
葡萄酒酿造是一个由不同微生物参与,相互作用的复杂过程。目前产区所面临的问题主要是产品同质化严重,缺少产区风格,葡萄酒酿造多依赖进口的商业菌株,所以研究葡萄中微生物的多样性对提高葡萄酒质量有举足轻重的作用。张世伟等[10]研究发现,不同酿酒葡萄果皮上的优势真菌菌属有Alternaria、Cladosporium、Phoma、Fusarium等。赵昱等[11]研究发现不同地区酿酒葡萄果皮上的优势真菌为Phenylobacterium、Ralstonia、Aureobasidium、Rhodotorula。本研究利用高通量测序分析了XXQ、YNX、QTX、SZS产区赤霞珠和霞多丽果皮上微生物多样性,检测到的优势真菌属有Alternaria、Cladosporium、Fusarium、Chaetomium、Didymella、Naganishia,Hanseniaspora、Aspergillus、Filobasidium。通过文献对比发现,葡萄果皮的优势菌属既有相同之处也有区别,结果造成差别的原因可能是因为品种、产区以及环境的不同。其中链格孢属大多数是有害菌,会引起很多植物疾病,有学者研究发现链格孢属会产生链格孢醇等多种有毒的代谢产物,这些毒素可使人慢性或急性中毒,有些还有致癌作用[12-13]。枝孢属和镰刀菌属也是有害菌,其中枝孢菌属会使植物产生叶斑,从而影响其光合作用,严重时还会使果实颜色变得暗淡[14];镰刀菌属可以感染很多植物,会引起植物根茎的腐败等多种病害,在代谢过程中会产生毒素,造成植物死亡,而且目前防治工作也是非常艰难的[15]。链格孢属在4产区中都大量存在,镰刀菌属在SZS产区大量存在,建议产区提前防治。
魏玉洁等[16]研究发现,新疆葡萄产区中酿酒葡萄果皮上优势的细菌菌属有Pseudomonas、Sphingomonas和Adhaeribacter。PARK等[17]和HIRANO等[18]研究发现,Pseudomonas、Acinetobacter和Kaistobacter是葡萄和叶片中的优势细菌菌属。LEVEAU等[19]研究发现Pseudomonas是霞多丽葡萄上的优势细菌菌属。本研究发现的优势细菌菌属有:Ralstonia、unidentified_Chloroplast、Massilia、Gluconobacter、Sphingomonas、Serratia、Pantoea、Pseudomonas。通过文献对比发现,葡萄果皮的优势细菌属既有相同也有区别。其中Ralstonia具有吸附水体镉的作用[20];Massilia具有消除环境中的多环芳烃菲、抗重金属、产酶溶磷等功能[21-22];Gluconobacter具有产葡糖酸盐、参与维生素C合成等功能[23];Serratia具有降解油脂的功能[24];Pantoea具有抑制黑曲霉的功能[25]。检测到的优势细菌属有些可能不是葡萄酒中常见的菌属,但大多数是有益菌,对提高葡萄酒的质量有间接作用。
4 结论
本研究利用高通量测序技术,对宁夏贺兰山东麓XXQ、YNX、QTX、SZS 4个产区的30份葡萄果皮样品的微生物群落结构进行分析,结果表明:
(1)在属水平上,无论是细菌多样性,还是真菌多样性,产区间都存在明显差异;
(2)不同产区赤霞珠葡萄样品中最为丰富的真菌属为链格孢属、枝孢属、镰刀菌属、毛壳菌属、亚隔孢壳属。产区微生物多样性有差异,但也有相似之处,产区距离相距越远,微生物相似度也越低;而在细菌属水平上,优势菌属为罗尔斯通氏菌属、unidentified_Chloroplast、马赛菌属、葡糖杆菌属、鞘氨醇单胞菌属,细菌的分布与产区地理位置相关性较小;
(3)不同产区霞多丽葡萄样品中最为丰富的真菌属为链格孢属、枝孢属、Naganshia、有孢汉逊酵母属和曲霉属,产区距离相距越远,微生物相似度也越低;而在细菌属水平上,优势菌属为沙雷氏菌属、罗尔斯通氏菌属、泛菌属、unidentified_Chloroplast和假单胞菌属,产区距离越近,相同的微生物种类也最多。
(4)同产区不同酿酒葡萄品种微生物的分布与采收期密切相关,酿酒葡萄采收期越晚,葡萄果皮上的真菌微生物种类也越多;而细菌的多样性却越低。
本研究对贺兰山东麓产区的微生物多样性进行了详细的分析,可为宁夏贺兰山东麓产区微生物资源库的构建、优良菌种的筛选、产区葡萄酒风格的确定以及提高产区葡萄酒质量提供理论基础。
