福建连城南方红豆杉古树种群结构及动态特征
2022-08-04谢春平沈顺霆刘大伟俞筱押黄绍虎
谢春平,沈顺霆,刘大伟*,俞筱押,黄绍虎
(1.琼台师范学院理学院,海口 571127;2.南京森林警察学院/野生动植物物证技术国家林业和草原局重点实验室,南京 210023;3.黔南民族师范学院旅游与资源环境学院,贵州 都匀 558000;4.南充市林业科学研究所,四川 南充 637099)
种群(population)是物种生存、繁衍与进化的基本单位[1];它是同一物种在一定时空内所有个体的集合[2]。因此,通过对种群结构特征分析,不仅可探究区域种群对环境的响应以及环境对种群结构的塑造[3],而且还可了解种群的发展历史[4],为种群恢复及森林经营管理提供参考依据[5-6]。种群结构是提供有关种群动态有价值信息的主要方式,它反映了种群不同年龄、大小个体的数量配置情况,也反映了种群数量动态与变化趋势[7]。静态生命表是了解种群结构特征的另一有效途径。静态生命表和存活曲线清晰地展示种群数量变化的潜在机制,并可阐明种群生存的客观规律[8-9]。种群数量变化动态指数则更进一步展现出不同龄级间种群数量的差异波动,体现了种群在有外界干扰和无干扰下的发展[10]。因此,种群结构是各种群更新与恢复的基础[11],它也成为了评价森林群落健康状况的重要参数。与其他物种相比,珍稀濒危植物种群结构具有较大的特殊性和脆弱性[12-14]。通过种群年龄结构特征、静态生命表和存活曲线等种群生态学方法对珍稀濒危物种进行研究,对种群的管理及具体保护措施的提出更具有理论价值与现实意义。
红豆杉属(Taxus)全世界有11个种,主要分布于北半球温带、寒温带及热带、亚热带高山地区;国产红豆杉属含4种1变种,零星分布于西南、东北、华中、华南及华东等地区[15]。鉴于红豆杉属植物资源的稀缺性,以及在植物区系地理、药用植物资源开发等方面的研究具有特殊意义[16-17],其种群保护生态学研究备受学者关注。据史料记载,在欧洲人定居北美之前,加拿大红豆杉(T.canadensis)是北美森林的常见种,至少在5%~20%林分中广泛分布;但从20世纪起,由于有蹄类动物啃食、火灾、密集的森林开发以及农业和其他土地开垦,加拿红豆杉的分布区骤缩而被列为当地的保护物种[18]。在对29个墨西哥红豆杉(T.globosa)种群现状进行评估后发现,近一半的种群出现了更新不良或完全无法更新的状况,而土地利用的改变、森林砍伐和生境破碎化的进一步恶化极有可能导致这些地方的墨西哥红豆杉种群消失[19]。在欧洲,英国红豆杉(T.baccata)也因为土地的过度集约利用,包括高强度的森林采伐,导致整个欧洲的英国红豆杉种群减少,濒临灭绝的境地[20-22]。我国分布的红豆杉属植物均被列为国家Ⅰ级重点保护,国内学者也对该属物种展开了广泛的种群生态学研究,如在对东北红豆杉(T.cuspidata)的研究中发现,过度的干扰直接导致该物种在中国的种群规模和栖息地面积大范围减少,使其处于濒危状态,面临局部灭绝的高风险[23]。滇西北分布的云南红豆杉(T.yunnanensis)种群结构呈纺锤型,表明该区域的红豆杉属衰退种群,这主要是受其自身生物学特性及人为干扰共同作用所致[24]。在对江西南方红豆杉(T.wallichiana var.mairei)天然种群的研究中发现,江西不同区域南方红豆杉种群整体上呈现出幼苗、幼树极少,自然更新能力差的状况;同时种群结构为纺锤型呈衰退趋势,亟待人工促进天然种群的恢复[25]。而对福建龙栖山南方红豆杉种群的研究发现,该种的种子休眠期长、发芽率低,且幼苗较高的死亡率也是导致其濒危的一个重要原因[26]。综上可知,导致红豆杉属植物濒危的原因一方面是因其自身生物学特性所致,另外很大程度上来自外界的干扰。
我国南方村落有保护古树的文化传统,具有丰富的古树资源;但多数古树呈零星分布,能够形成古树群落的则较为罕见,因此较少学者关注古树种群。作者在福建连城县野外考察时发现,该县山区村落周边分布有南方红豆杉古树种群,而部分种群因风水林的性质得到了一定程度的保护。然而,这些红豆杉古树种群与人居环境十分接近,受到了较为强烈的干扰,与天然种群又有一定的差别。基于此,本文主要以福建连城县赖源乡郭地村和曲溪乡白石村分布的2个南方红豆杉古树种群为研究对象,通过对种群年龄结构特征、静态生命表等分析,揭示当前种群结构现状及预测种群的发展趋势。最后,旨在为该种群恢复与更新、古树资源保护等方面提供科学依据。
1 材料和方法
1.1 研究地概况
连城县地处武夷山南段东侧(24°13′~25°26′N,116°32′~117°10′E),是福建省西部地区的重点林区县。该县属中亚热带海洋性季风气候,年均温18.9℃,年均降水1 734 mm,年均日照时数1 760 h;年平均水汽压18.2 hPa,相对湿度78%,雨季多集中在春夏季,全年无霜期长达290 d以上[27]。研究区内的土壤以黄红壤土和红壤土为主。连城县森林覆盖面积达80%以上,以米槠(Castanopsis carlesii)、福建青冈(Cyclobalanopsis chungii)等为优势树种构成了本区域的顶级原生阔叶林群落,在中国植被区系划分中属南岭东部常绿槠类林区。区域内的森林类型有落叶阔叶林、常绿阔叶林、常绿针叶林、针阔混交林和毛竹林等[28]。本研究选取了连城县境内有南方红豆杉古树分布的10个群落进行调查,面积最小的仅100 m2;但具有一定种群数量,满足研究基本条件的仅赖源乡郭地村(600 m2)和曲溪乡白石村(600 m2)的两个种群;所选群落包含了所有的古树个体。南方红豆杉古树群落内,除南方红豆杉为乔木层的优势树种外,乔木层的其他伴生树种有毛竹(Phyllostachys edulis)、杉木(Cunninghamia lanceo⁃lata)、马尾松(Pinus massoniana)、枫香(Liquidambar formosana)、木荷(Schima superba)、深山含笑(Mi⁃chelia maudiae)、棕榈(Trachycarpus fortunei)等;灌木层有连蕊茶(Camellia fraterna)、檵木(Loropeta⁃lum chinense)、高粱泡(Rubus lambertianus)、箬竹(Indocalamus tessellatus)和椤木石楠(Photinia david⁃soniae)等;草本层有水鬼蕉(Hymenocallis littoralis)、铁芒萁(Dicranopteris linearis)、卷柏(Selaginella tamariscina)、阿拉伯婆婆纳(Veronica persica)、繁缕(Stellaria media)和蛇莓(Duchesnea indica)等。
1.2 研究方法
1.2.1 径级划分
南方红豆杉木材坚硬,用生长锥钻芯而获取实际年龄难以实现,且伐木计数年龄又不适用于古树。因此,本文选用空间推时间的方法,即用胸径替代龄级反映种群结构[26]。将古树群落内的南方红豆杉所有个体划分为10个径级,即胸径(DBH)<10 cm及无胸径的个体的设定为Ⅰ级,往后每间隔10 cm划定为一级:10≤DBH<20,……,80≤DBH<90,而DBH≥90为第Ⅹ级,逐一统计各径级的个体数。
1.2.2 种群静态生命表和存活曲线
生命表的内容包括:x代表种群径级;ax表示x径级中种群实际存活个体数;lx代表种群中自x径级为始的标准化存活个体数(1 000);dx表示从x至x+1径级区间内种群个体的标准化死亡数;qx代表从x至x+1径级区间内种群个体的标准死亡率;Lx代表种群中自x至x+1径级期间平均存活个体数;Tx表示从x至大于x径级的种群总个体数;ex代表步入x径级种群个体的生命期望;Kx表示植物种群致死率(消失率)。生命表中的各项之间关联紧密,可通过实测ax数值以及以下公式计算[29]:
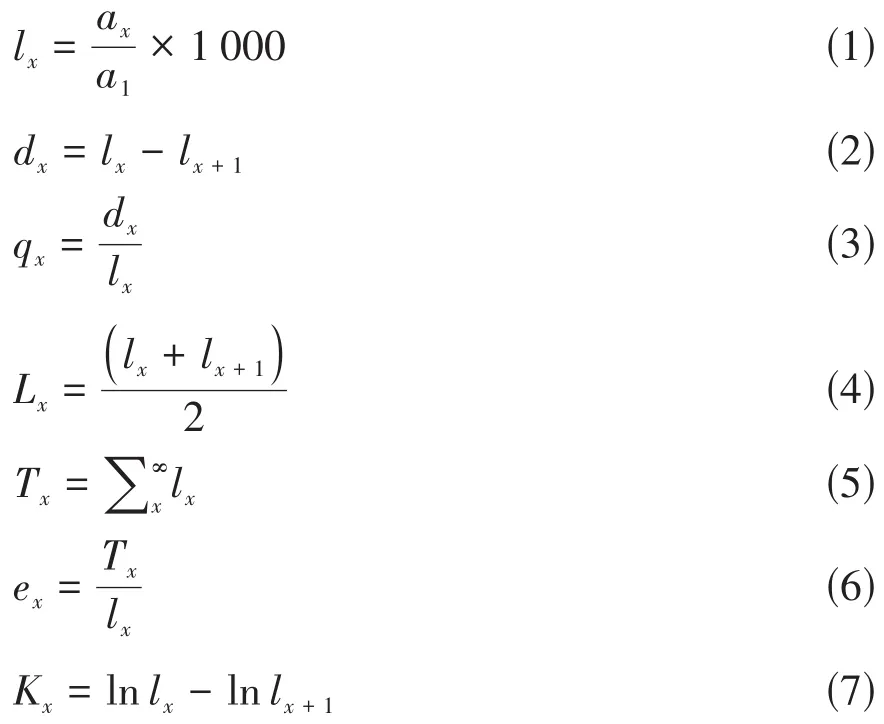
上述公式中:a1表示种群中第Ⅰ径级实际存活个体数,lx+1代表种群中第x+1径级的标准化存活个体数。依照南方红豆杉种群径级结构数据编制种群静态生命表和描绘生存曲线[2,8,30]。
1.2.3 种群动态量化分析
①无外部干扰时的植物种群年龄结构的数量变化动态指数:
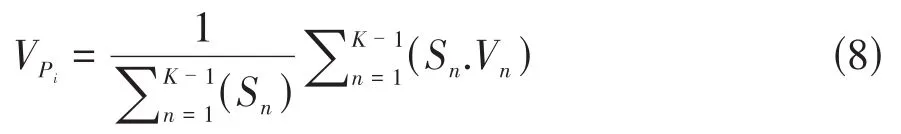
②随机干扰时植物种群年龄结构数量变化动态指数:
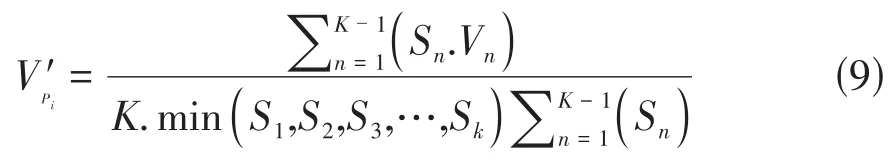
③种群对外界干扰所承担的风险概率:
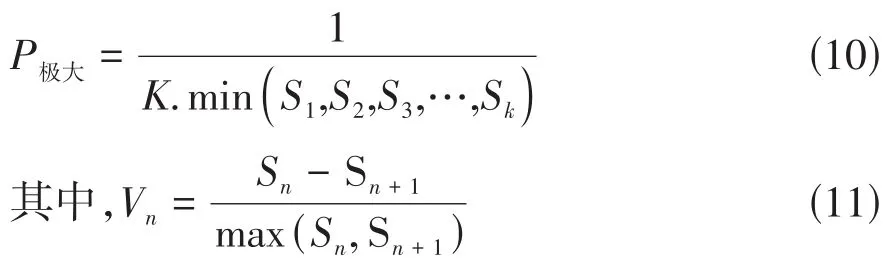
Sn与Sn+1分别为第n与第n+1年龄级种群个体数,min(…)分别表示取括号中数列极小值[7,10]。
2 结果与分析
2.1 种群结构特征
据调查,2个南方红豆杉古树种群共有47株个体,其中最大胸径142 cm,最小胸径8 cm,种群各径级的个体数存在较大差异(图1)。整体上,种群数量偏少、结构不规则,两地种群个体数的峰值均出现在Ⅱ径级,分别占种群总数25%和18.18%;但径级Ⅰ的幼树与幼苗缺失,个体存活率较低。这反映出古树种群更新力低、补充不足的特点,对古树种群未来繁衍极为不利。从赖源乡郭地村的种群结构来看,Ⅰ径级和Ⅵ径级个体存在缺失的情况,种群结构出现断层现象。该地种群的第2峰值则出现在大径级的Ⅳ和Ⅷ,分别占整个种群数量的16.67%。从上述数据可以看出,该地古树种群径级结构波动明显,种群在发展的过程中的稳定性差;加之Ⅰ径级个体缺失和种群断层,都表明该地种群属于衰退型。与赖源乡郭地村相比,曲溪乡白石村的种群龄级结构呈连续分布状,但径级Ⅰ的个体数较少,仅占整个古树种群的9.09%,故种群更新能力显弱。Ⅱ~Ⅴ径级,种群个体数之和占总体比例的近60%,从径级Ⅵ~Ⅸ则呈递减状。因此,曲溪乡白石村的种群近似呈中间大两头小的纺锤型结构,为弱增长型种群。
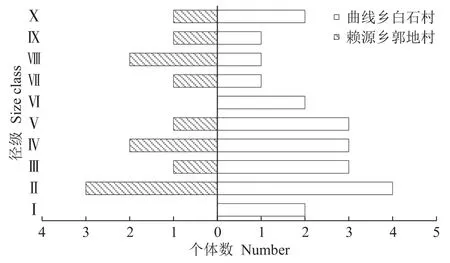
图1 连城南方红豆杉古树种群径级结构Figure 1 Diameter class structure of T.wallichiana var.mairei population in Liancheng County
2.2 种群静态生命表
静态生命表显示(表1),两地南方红豆杉古树种群大体上随着径级的增加,存活数(lx)呈下降的趋势。表1中,两个南方红豆杉古树种群的死亡率(qx)均有负数出现,如赖源乡种群的Ⅲ和Ⅶ径级、曲溪乡种群的Ⅰ和Ⅸ径级,这与数学假设相矛盾;但负数仍具有生态学意义,即表明古树种群数量处于衰减的波动变化中[31]。进入x径级的种群个体生命期望值ex能客观反映个体生命力和平均生存能力的高低。随径级逐渐增大,两地古树种群的生命期望值也存在一定的差异,赖源乡郭地村最大值为9.0,出现在第Ⅲ径级,反映出种群在该径级最具活力;曲溪乡白石村Ⅰ级生命期望值最大为11.0,种群活力最大。此外,从消失率(Kx)最大值出现的情况看,两地均是在中等径级出现,分别在Ⅴ(5.81)和Ⅵ(0.69),且赖源乡的种群消失率要明显高于曲溪乡。由此可见,达到中等径级后,种群的活力和生命期望值均开始下降。
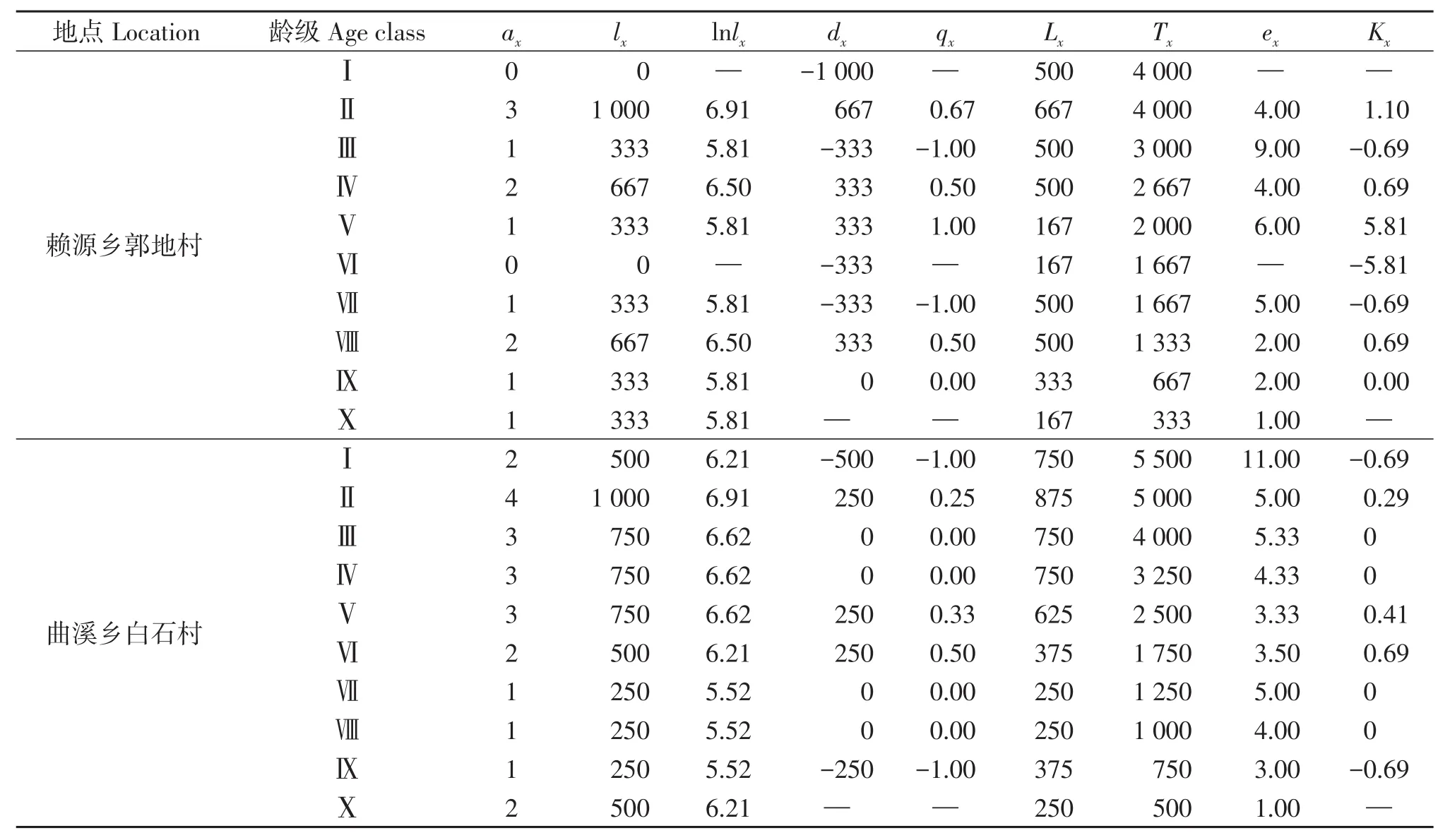
表1 连城南方红豆杉古树种群静态生命表Table 1 Static life table of T.wallichiana var.mairei population in Liancheng County
2.3 存活曲线
存活曲线显示(图2),赖源乡郭地村南方红豆杉古树种群与标准的Deevey-Ⅰ、Deevey-Ⅱ和Deevey-Ⅲ型存活曲线有较大的差别[32]。由于种群在Ⅰ和Ⅵ径级个体缺失,致使曲线表现出不规则“M”型,这与Deevey-Ⅱ型的b1变型近似,即在种群生长的多个时期出现波峰波谷式的交替变化,个体存活率差异十分明显。这一现象的产生原因可能与种群周期性动态变化及与该地种群生境的敏感性有关。一方面外部生境的干扰作用使种间竞争加剧,部分物种的生态位重叠,尤其是更新层物种对各类资源的需求的相似性较大;另外,若种群在早期丧失主动权(如进入乔木层较晚),则种群在后续的竞争则处于劣势。曲溪乡白石村的南方红豆杉古树种群存活曲线与Deevey-Ⅲ型则较为接近,整体呈现“凹型”,即种群幼苗期个体死亡率较高。采用Hetthe & Louck提出的幂函数和指数函数模对两个古树种群的存活曲线进行检验[33],均不显著。这可能与古树本身的种群结构特性及生境特殊性有关。
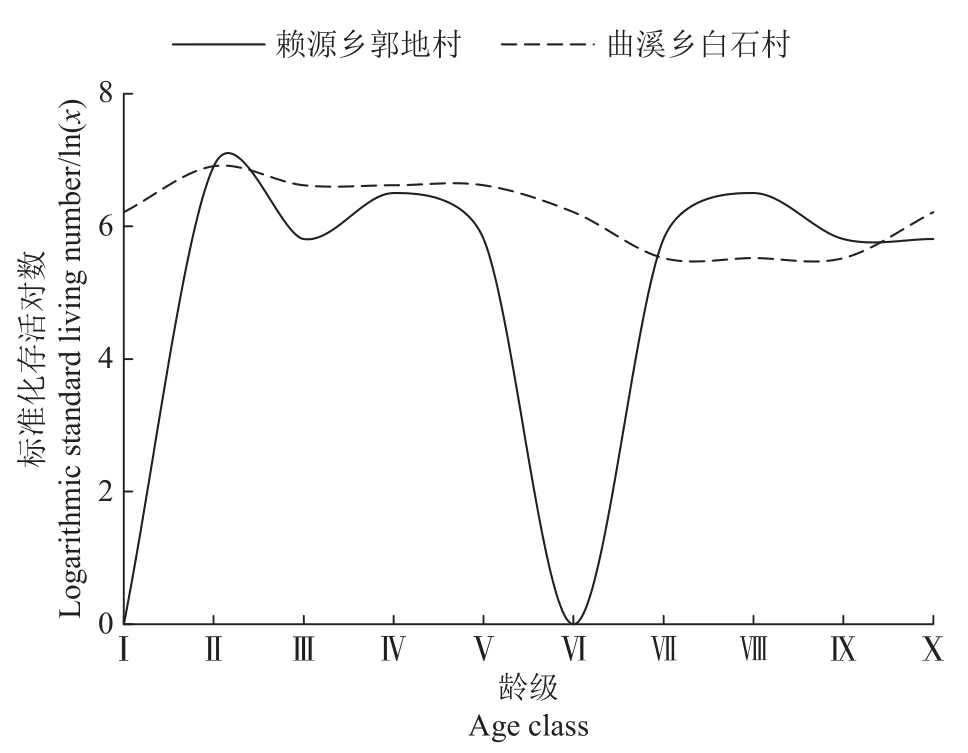
图2 连城南方红豆杉古树种群的存活曲线Figure 2 Survival curves of T.wallichiana var.mairei population in Liancheng County
2.4 种群数量动态特征
种群动态量化分析清楚地显示出两地古树种群在不同径级间的数量变动(表2)。赖源乡郭地村的数据显示,从V1~V10的数值分别为-100.0、66.7、-50.0、50.0、0.0、-50.0、50.0、0.0和100.0,由于中间Ⅵ径级个体数为0,故该层数据缺失。曲溪乡白石村的 V1到 V10则为-50.0、25.0、0.0、0.0、33.3、50.0、0.0、0.0、-50.0和100.0。由于正、负和零分别表示种群的增长、衰退和稳定,因此各径级间存在明显的波动现象。其次,在没有外部干扰时两地种群的数量变化动态指数VPi分别为-3.03和7.5,而存在外部干扰时的数量变化动态指数VPi'则分别为-0.34和0.75。由此可知,不论是否存在外部干扰赖源乡郭地村的南方红豆杉古树种群均属衰退型,而曲溪乡白石村的则为弱增长型;这也与种群结构特征相互印证。两个种群的随机干扰风险极大值(P极大)分别为0.11和0.10,均表明它们对外界干扰具有较高的敏感性。
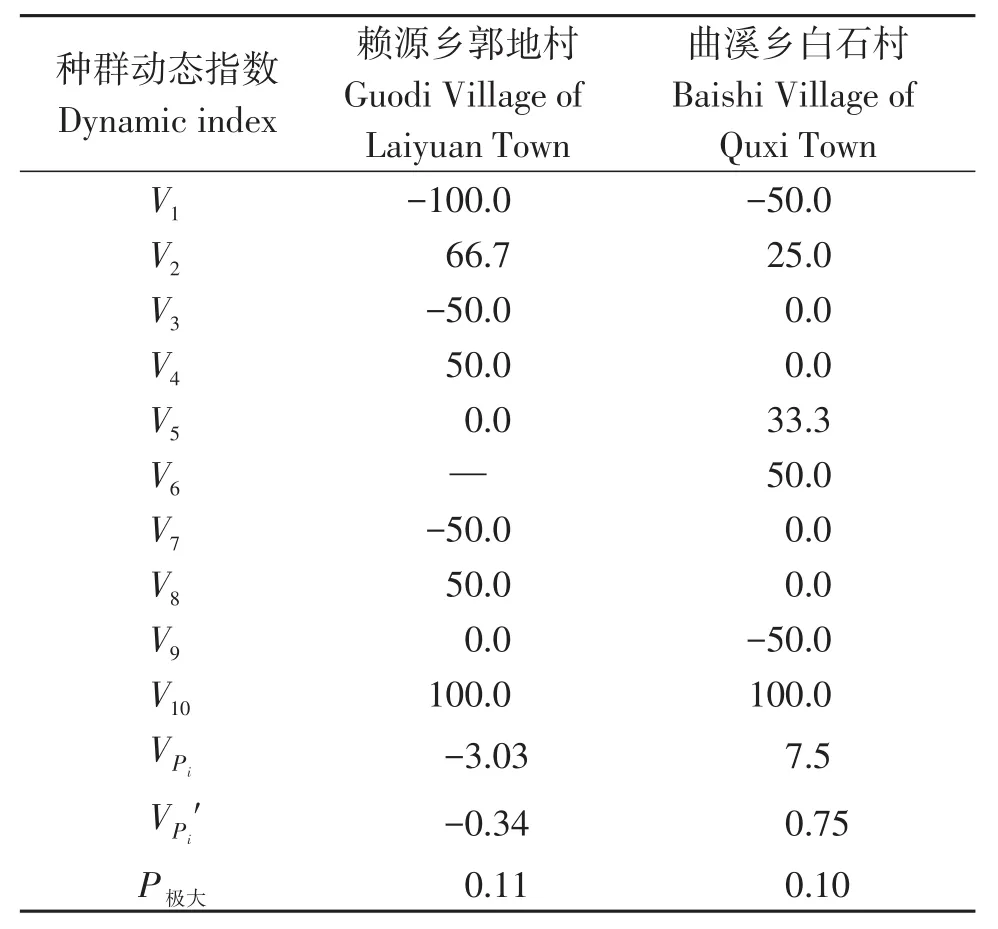
表2 连城南方红豆杉古树种群的种群动态变化指数Table 2 Dynamic indices of T.wallichiana var.mairei population in Liancheng County
3 讨论
虽然种群长期定位观测所获取的动态数据较短暂调查的静态数据更有价值[34],但种群结构特征、静态生命表的信息至少反映了当前种群在不同龄级的个体分布、更新、迁移和死亡等数量动态,仍可为几年或几十年的种群发展趋势做出有效的预测[35-36]。种群结构不合理,幼树幼苗不足,是当前连城县南方红豆杉古树种群面临的严峻问题。从种群结构特征来看,虽然曲溪乡白石村的南方红豆杉古树种群结构要优于赖源乡郭地村,但两地均存在幼树幼苗严重缺失的局面,这将是制约种群发展的关键因素。
南方红豆杉种子一般具有深度休眠的特性,需要在种子成熟后的2年左右才会萌发;在这段时期将经历被真菌感染(霉变)、种子腐烂和动物取食等风险,真正能够萌发的种子不多[25]。邓青珊等在皖南仙寓山鸟类对南方红豆杉种群的更新及分布格局的研究中指出,掉落于树冠下的种子在母树下虽能萌发,但存活率几乎为零;但经鸟类吞食后随粪便排出的种子萌发率却得到明显的提高[37]。这说明,能够天然萌发的南方红豆杉种子与鸟类形成了互惠关系,即南方红豆杉的种子为鸟类提供了食物,而鸟类则帮助其传播和提升萌发率。
从种群的空间分布格局来看,红豆杉属内的植物在幼龄期多为聚集分布[24,38-39],即多数种子在成熟后散落在母树树冠下集中萌发,使得幼苗和幼树在种群初期高度集中于母树下[39]。在种群发展的过程中,因微环境及种群个体差异,种内竞争的激烈程度要大于种间竞争,使得大量个体在种群的初期死亡。在调查的样地内,还有一些亚热带常见优势树种的幼苗,如苦槠、青冈和木荷等,它们对当地环境的适应和对资源的竞争力也不会弱于南方红豆杉。随着群落的进一步发展,能够进入群落中层的南方红豆杉个体数更少,最后仅有少数在群落中得到稳定;这在生命表中也有所体现。
南方红豆杉属耐阴植物,在幼苗的建成期对强烈的直射光敏感,但过度荫蔽又使其生长不良[25]。因此,当古树群落形成后,群落结构稳定,郁闭度高,满足南方红豆杉幼树幼苗的生境减少。同属的云南红豆杉也是在幼年期对光较为敏感,成年后需光性增强,而早期多数幼树幼苗均被环境筛除[24]。虽然曲溪乡白石村的南方红豆杉古树种群呈弱增长,但Ⅰ径级个体数少,各径级之间无法实现较好地逐级递补,这种弱增长将难以持续。当受到外界环境干扰后,种群则易出现衰退;负的种群动态变化指数及随机干扰风险极大值也证明了该种群对当前环境具有较高的敏感性,这与其他一些珍稀濒危植物对环境的反应相一致[30,40]。
外界干扰对红豆杉属植物种群发展所造成严重影响已被许多研究所证实[18-19,21-22]。类似的,强烈的外界干扰是影响当前南方红豆杉古树种群发展的关键性因素之一。作者在所调查的10个南方红豆杉古树种群发现,多数分布在道路两侧,附近常有墓地、养蜂场和垃圾堆积处等人类活动的痕迹;而古树种群又多与毛竹林为邻。频繁的竹林管理、竹笋采挖和捕鸟等人类活动对南方红豆杉幼树幼苗的建成有极大的影响。在这10个调查地中,多数古树种群仅有3~5株个体,呈零星分布状,唯有赖源乡郭地村和曲溪乡白石村的南方红豆杉形成了一定的规模。这两个种群同时也是当地的风水林,具有特殊的文化和宗教意义[41-42];这在一定程度上对种群起到了保护作用[43]。但是,走访中发现,频繁的人类活动(祭祀等)及人为有意或无意地移除幼苗是导致这两个种群Ⅰ径级个体数少的重要原因。
有学者提出,红豆杉属植物濒危的主要原因是其幼树幼苗的生长过于缓慢,与群落内的其他物种的竞争力差所致,而非普遍认为的人为干扰[44]。从调查结果看,南方红豆杉的竞争能力较弱而使分布受限,只能存在于没有快速生长树种的群落内。但是应根据每个调查地的实际情况进行判断,本研究中人为干扰对种群所造成的影响仍是不可忽略的。
基于南方红豆杉古树种群在连城的分布现状以及调查分析结果,作者建议:①赖源乡郭地村和曲溪乡白石村应建立南方红豆杉种群保护小区。同时利用风水林的性质,结合中国传统生态哲学思想营造出“人与自然和谐共处”的群落。这样既保护了古树,种群又得到了保护和恢复,同时对古树后备资源的孕育起到了较好的作用。②由于古树群落多数已郁闭,南方红豆杉幼苗在母树下难以成活,因此可在法律法规的指导下进行适当移植,进行迁地保护。③对南方红豆杉古树种群下的杂灌进行移除,减少种间竞争。④加大对周边村民生物多样性保护教育,提升保护意识。一方面让村民减少对种群的干扰,尤其是杜绝踩踏、放牧、拾取种子等行为;另一方面,也可让村民协助林业管理部门进行日常的监测与维护,出现病虫害或外界破坏时及时上报处理。
本文仅就南方红豆杉古树种群的结构特征进行了初步的研究,但微环境对种群的影响、种间竞争、群落结构对物种的影响以及种群长期的动态过程等均未涉及;同时这两个种群本身又具有风水林的性质,形成过程与人类活动有密切关系。这些方面对种群的发展均有较为深刻的影响,有待进一步深入探讨。
4 结论
通过对福建连城县2个南方红豆杉古树种群结构及动态特征研究表明,2个种群的Ⅰ径级个体数缺失严重,其中赖源乡白石村种群的径级结构不完整,为典型的衰退状;曲溪乡白石村为弱增长型,但仍是以中等径级为主的种群,不利于种群的发展。当前种群结构不良的原因除了物种本身的生物学和生态学特性因素外,人为干扰和种间竞争也是限制其种群发展的主要原因。因此,采取合理的保育措施,提升南方红豆杉种群种子的萌发率及增加小径级的个体数是促进种群恢复的关键。
