药食同源植物薤白的谱系地理学研究
2022-08-04莫忠妹
石 甜 莫忠妹 吴 敏 赵 财
(贵州大学生命科学学院/农业生物工程研究院,山地植物资源保护与保护种质创新教育部重点实验室,山地生态与农业生物工程协同创新中心,贵阳 550025)
薤白()又名小根蒜,为石蒜科(Amaryllidaceae)葱属()多年生宿根性草本植物,野生于山野和田间,全国大部分地区均有分布。薤白可作食用,鳞茎也可药用,营养价值丰富,是重要的药食同源植物之一,具有理气宽胸和行气导滞等功能。目前,对薤白的研究主要集中在化学成分、药理作用及生药来源等方面。少数学者对薤白的系统发育、亲缘演化关系等方面进行了研究,如黄德青等利用ITS 和LF序列探讨了薤白的系统位置,认为其可能代表一个新的组一级类群;杨鹏等和莫忠妹等分别对贵州和西南地区的薤白进行了亲缘关系研究,支持薤白作为一个单系类群,并认为其亲缘关系与其地理位置具有一定相关性。
地质历史和气候振荡是植物进化和遗传结构形成的重要驱动因素。新生代以来,特别是第四纪冰期,不同纬度地区动植物区系的地理分布格局、遗传分化和生态适应性受到寒暖气候变动、冰期和间冰期反复交错、地质事件及季风波动的较大影响。中国地形地势西高东低,地形多种多样,地貌类型复杂多样,自然环境存在较大差异。青藏高原强烈抬升是最重要的地质事件之一,其改变了全球气候并激发了东亚季风,使我国植物区系经向分带愈发明显。中国因其特殊的地理位置和复杂的气候环境,具有丰富的植物多样性和独特性。虽然在冰期时中国大部分地区没有被大规模的冰层覆盖,但全球气候发生了大规模变化,尤其是第四纪冰期和间冰期周期性交替造成的寒冷和温暖气候变化引起的气候波动和青藏高原抬升影响了季风强度进而影响当地气候波动,干湿交替深刻影响植物在中国的地理分布格局和历史演化进程。同时,复杂的地质运动、环境异质性和河流袭夺也深刻影响着现存植物的地理分布格局和种群遗传结构。
薤白作为草本植物,分布范围广,跨越不同气候带和多个生物多样性研究热点区域,是进行广布连续分布植物谱系地理学研究的理想材料。同时,草本植物对气候变化和环境波动敏感,DNA 碱基替换速率高于木本植物,能够提供更多的历史演化信息。目前,有关薤白谱系地理学的研究尚未见报道,因此本研究首次采集中国整个分布区的14 个薤白居群,利用叶绿体基因(AH16 和L-F)和核基因(ITS)序列进行PCR 扩增及测序,用以研究其谱系地理结构,探讨其遗传结构及单倍型的地理分布格局,并推测其在第四纪冰期的避难所,为进一步探讨中国植物区系演化及物种多样性提供一定的理论依据。
1 材料与方法
1.1试验材料
薤白材料于2016—2019年采自浙江、甘肃、吉林和四川等地,共14 个自然居群,各居群采集10~15个个体,位置信息采用全球定位系统测量(见表1)。野外采集薤白的新鲜叶片后立即用硅胶干燥保存,用于总DNA 提取。以葱属植物单花韭()作为外类群构建单倍型系统树。

表1 薤白居群地理分布信息Table 1 Geographical distribution information of populations of A.macrostemon
1.2 DNA 提取及扩增测序
采用改良的CTAB 法从硅胶干燥的叶片中分离总基因组DNA。查阅相关文献,参照Hamil‑ton、Oxelman 等、Taberlet 等和Wendel 等设计引物,利用3 对叶绿体基因片段(A-H16 和L-F)和核基因ITS 序列,获取引物序列并合成引物,用于筛选适合的基因片段。引物序列及PCR 扩增程序见表2。将扩增后检测结果符合标准的产物送往生工生物工程(上海)股份有限公司进行纯化及测序。

表2 PCR引物信息及反应程序Table 2 Primer and protocols of PCR
1.3数据分析
利用DNAstar软件和MEGA 7.0软件对测序数据进行编辑和手工核对调整。运用DNaSP 5.0 软件统计单倍型数量,计算各居群单倍型多样性()、核苷酸多样性()、Tajima’s值以及Fu & Li’s值,并进行失配分析以检测是否经历扩张事件。薤白总遗传多样性()、居群内平均遗传多样性()、遗传分化系数(和)利 用PERMUT 计 算。基 于 简 约 性 原 则 利 用Network 软件中的Median-Joining 构建单倍型网络图。利用Arlequin 软件包中的AMOVA 分析计算群体遗传结构以及检测居群内和居群间遗传变异,并计算遗传分化系数(),基因流()进一步揭示居群的分化程度。以近缘种单花韭为外类群,利用MEGA 7.0 软件和MrBayes 3.2.1软件分别采用邻接法(NJ)、最大似然法(ML)和贝叶斯法(BI)构建的叶绿体及核基因单倍型系统树。
2 结果与分析
2.1薤白叶绿体单倍型和核基因基因型序列变异与遗传多样性
薤白叶绿体A-H+16+L-F 联合序列成功测序160个个体,测序所得数据对位排列后序列长度为1 930 bp,包含9 个变异位点,G+C 含量为35.52%(见表3)。运用DNaSP 软件得出薤白总单倍型多样性为0.866,大盘山(DPS)和天水(TSS)的单倍型多样性最高(0.545);总核苷酸多样性为0.780×10,大盘山(DPS)具有最高的核苷酸多样性(0.34×10)(见表4)。薤白ITS 序列成功测序165个个体,经比对后其长度为633 bp,包含214个变异位点,G+C 含量为50.22%。总单倍型多样性为0.835,剑阁(JGX)具有最高单倍型多样性(0.667);总核苷酸多样性为64.78×10,其中天水(TSS)核苷酸多样性最高,为7.170×10(见表4)。

表3 psbA-trnH、rps16、trnL-F及ITS序列信息Table 3 Information of psbA-trnH,rps16,trnL-F and ITS

表4 薤白居群内单倍型分布及多态性信息Table 4 Haplotype distribution and polymorphism of haplotypes in population of A.macrostemon
2.2薤白叶绿体和核基因单倍型分布与种群遗传结构
对A-H+16+L-F 联合序列进行了单倍型检测,共得到11 个单倍型(H1~H11),其中单倍型H3 的个体数为35 个,出现的频率最高,分布范围最广,在大盘山(DPS)、内蒙古(NMG)及银川(YCS)均有分布;其次为单倍型H5,分布在广州(GZS)、黄石(HSS)及通化(THS)(见图1)。基于核基因ITS序列共得到15个基因型(H1-H15)。其中单倍型H1 的频率最高,包含60 个个体,在大盘山(DPS)、黄石(HSS)、怀化(HHS)、信阳(XYS)、福州(FZS)均有分布(见图2)。薤白的叶绿体单倍型网络分析结果显示,以单倍型H3 和H5 为中心分别衍生出其他单倍型,推测H3 和H5 为古老单倍型(见图3A)。薤白的核基因单倍型网络图显示薤白以单倍型H1 为中心,衍生出H2 和H6 两个单倍型,同时衍生出其他单倍型,推测H1 为古老单倍型(见图3B)。以近缘种单花韭为外类群,利用MEGA 7.0 软件分别构建叶绿体及核基因单倍型系统树,建树结果与单倍型网络图基本吻合(见图4)。利用核DNA 序列的15 种单倍型构建的系统发育树可以看出,单倍型H1、H2、H6 与H11~H15聚为一支;其余单倍型聚为一支(见图4)。

图1 薤白14个居群叶绿体单倍型地理分布饼状图表示每个居群中单倍型的频率,下同Fig.1 Haplotype geographical distribution of chloroplast sequences in natural populations of A.macrostemonThe pie chart shows the frequency of haplotypes in each population,the same as below

图2 薤白14个居群核基因单倍型地理分布Fig.2 Haplotype geographical distribution of nuclear sequences in natural populations of A.macrostemon

图3 薤白种群单倍型网络A.叶绿体;B.核基因Fig.3 Haplotype network of A.macrostemon populationsA.The haplotype network of chloroplast sequences;B.The haplotype network of nuclear sequences

图4 薤白单倍型系统发育树A.叶绿体基因单倍型系统发育树;B.核基因单倍型系统发育树;分支上的数字分别代表NJ树、ML树和BI树的支持率,未标明支持率处表示NJ树、ML树和BI树存在略微差异Fig.4 The phylogenetic tree of A.macrostemonA.The phylogenetic tree of cpDNA haplotypes;B.The phylogenetic tree of nrDNA haplotypes;The Numbers on the branches represent the support rates of NJ,ML and BI trees respectively,and the unmarked support rates indicate that there is a difference among NJ,ML and BI trees
利用PERMUT 计算得出,薤白叶绿体片段总遗传多样性为0.868,居群内平均遗传多样性为0.111;居群间遗传分化系数为0.866、为0.918(>0.05);基因流为0.019。核基因片段总遗传多样性为0.890,居群内平均遗传多样性为0.130;居群间遗传分化系数为0.855、为0.984(>0.05);基因流为0.004。AMOVA 分析结果显示,叶绿体基因和核基因在薤白居群间遗传变异(cpDNA:92.84%;nrDNA:98.40%)显著高于居群内遗传变异(cpDNA:7.16%;nrDNA:1.60%),说明遗传变异主要存在于居群间。遗传分化系数均大于0.25(cpDNA:0.928;nrDNA:0.984),说明薤白群体的遗传分化程度很高,存在明显的隔离(见表5),且叶绿体和核基因序列居群间遗传分化系数均为>,表明该物种具有明显的谱系地理结构。
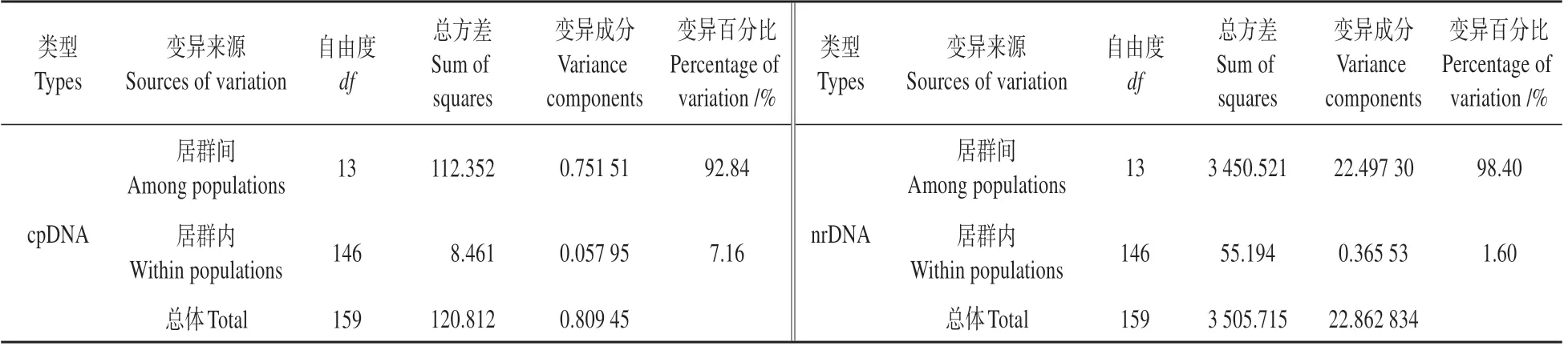
表5 薤白种群的分子方差分析结果Table 5 Analysis of molecular variance(AMOVA)in A.macrostemon populations
2.3种群历史变化动态分析
利用DNaSP5.0软件对薤白的Tajima’s值以及Fu&Li’s值进行计算,得出其叶绿体基因Ta‑jima’s统计值为-0.040 56(>0.10);Fu & Li’s值为1.225 72(>0.10);Fu & Li’s值为0.930 56(>0.10);Fu’s值 为-1.723。此 外,利 用DNaSP5.0对薤白进行失配分析以检测是否经历扩张事件,其叶绿体失配分析曲线呈单峰分布(见图5A)。这些研究结果表明,薤白经历过近期扩张过程。同时,利用DNaSP5.0 计算得出的核基因Taji‑ma’s统计值为0.111 90(>0.10);Fu&Li’s值为2.866 30(<0.02);Fu & Li’s值为1.874 44(<0.05);Fu’s值为50.426。利用DNaSP5.0 完成的核基因失配分析曲线为多峰曲线(见图5B),期望值与观测值相悖,违背居群扩张模型,说明薤白居群并未经历显著的群体扩张,而是处于动态平衡状态。

图5 薤白种群失配分析A.叶绿体基因失配分析;B.核基因失配分析Fig.5 Mismatch analysis of A.macrostemonA.The mismatch analysis of cpDNA;B.The mismatch analysis of nrDNA
3 讨论
3.1遗传多样性
本研究利用的叶绿体基因A-H+16+L-F联合序列结果显示,叶绿体基因遗传变异高于被子植物叶绿体DNA 的平均遗传多样性0.67,核基因ITS 序列变异也高于核基因变异平均值0.137。遗传多样性与物种地理分布、生物学特性、种群大小及繁育体系等诸多因素相关。目前,利用不同分子标记探讨草本植物遗传多样性的研究较多,如无苞香蒲()、宽叶香蒲()和长苞香蒲()结果显示其遗传多样性较低,可能与其具有较高的自交率和较强的营养繁殖能力有关。本研究中,薤白居群间叶绿体遗传多样性和核苷酸多样性都低于核基因,这可能是与叶绿体基因的相对保守性及其母系遗传方式相关,叶绿体DNA 片段的基因流大部分是通过种子来传播,而核基因的基因流传播方式包括花粉和种子。此外,分布狭窄的物种遗传多样性低于广布种,而薤白为广布种,数量较大,适应性较强,可以保留相对丰富的遗传多样性,这也是一个薤白遗传多样性较高的原因。
3.2遗传结构
地质历史气候变化、地势条件、生境异质性,以及种子基因交流程度等诸多因素与植物种群的遗传结构密切相关。通过对叶绿体A-H+16+L-F 联合片段分析,结果显示居群遗传分化系数小于,表明居群间存在显著的谱系地理结构。总遗传多样性显著高于居群内平均遗传多样性,说明薤白居群间没有频繁的基因交流,而基因流(=0.019)数值较低,也证实薤白居群间仅存在较低的基因交流。这一结果与其他物种的研究结果一致,如基于cpDNA 联合数据对迎红杜鹃亚属特有种进行的遗传结构分析,显示该亚属两个种的遗传变异主要存在于居群间,且显著小于,说明具有明显的谱系地理结构;朱仁斌对文冠果()的遗传结构分析得出该物种居群间出现强烈的遗传分化,居群间基因流较小,存在谱系地理结构(<)。基于对核基因ITS 序列分析,可知薤白居群<(=0.855,=0.984,>0.05),表明居群间存在显著的谱系地理结构。总遗传多样性(=0.890)与居群内平均遗传多样性(=0.130)差值较大,基因流(=0.004)数值较低,薤白居群间只存在很低的基因交流,说明薤白居群间没有频繁的基因交流。
通过对叶绿体基因和核基因分析显示大于,表明薤白的地理分布存在谱系地理结构,与基于单倍型数据构建的系统发育树分支结果一致,形成了与遗传距离相关的谱系地理格局。AMO‑VA 分析结果显示,叶绿体基因和核基因在薤白居群间遗传变异分别为92.84%和98.40%,均显著高于居群内遗传变异,说明薤白遗传变异存在于居群间。叶绿体基因和核基因的中性检验都不支持种群经历过扩张,但叶绿体歧点分布支持种群扩张模式,而核基因歧点分布不支持空间扩张模式。综上所述,认为薤白居群没有经历过近期扩张,这可能是薤白种子结实率低、不易向外扩散传播及生境原因;并且本研究结果显示,薤白14个居群间叶绿体基因和核基因的基因流仅为0.019 和0.004,因此认为地理隔离引起的有限基因流也是造成薤白居群间存在显著遗传分化的原因之一。
3.3薤白种群地理分布格局及冰期避难所
冰期避难所是由于极端环境变化而导致物种迁移和生存的地方。根据冰川避难所理论,遗传多样性高、古单倍型和地方性单倍型较多的地区可能是该物种在冰期的避难所,这些区域在环境波动期间具有相对稳定的生态条件,因此促进了遗传多样性的积累。薤白在我国不同地区复杂的地形及环境中均有分布,在适应不同生境过程中,它们经历了不同的遗传历史,而生境片段化和人为干扰会导致其具有谱系地理结构和强烈的遗传分化。第四纪冰期期间,全球范围气候和地势条件产生巨大变动,对所有动植物都造成很大影响,导致生物物种进行地理位置上大规模的转移。根据溯祖理论,一般位于单倍型网络图中间位置的为古老单倍型,可据此推断避难所位置。
李慧敏对槲树()进行冰期避难所推测,认为末次间冰期时秦巴山区为槲树最大的冰期避难所,而长白山地区可能是槲树的另一个冰期避难所;张华杰揭示在冰期时期,山莓草()在亚洲存在几个避难所,其中我国东北长白山和日本是山莓草在亚洲的重要避难所,青藏高原东部存在多个微型避难所;对大花旋蒴苣苔()的谱系地理学研究也认为东部地区存在其冰期避难所。本研究中,结合cpDNA序列和ITS序列进行分析,大盘上、天水和通化等地区显示出较高的遗传多样性,并且存在古老单倍型。结合前人研究,推测薤白在第四纪冰期时可能在分别位于东部、秦岭和长白山的大盘山、天水和通化地区存在几个冰期避难所。这些分析可为类似草本植物的进化提供参考,丰富我们对东亚草本植物分子系统与生物地理学的认识。
