穿龙薯蓣的花序特征研究
2022-08-04张雨晴高润梅党裳霓冯宇梅白甜雨
张雨晴 高润梅 党裳霓 冯宇梅 杨 柳 白甜雨
(山西农业大学林学院,太谷 030800)
穿龙薯蓣()为薯蓣科(Di‑oscoreaceae)薯蓣属()根状茎组(Sect.Ste‑nophora Uline)多年生缠绕草本植物,在我国主产于东北和华北地区的低山林下。现代医药工业从薯蓣属植物根状茎中提取的薯蓣皂苷元(dios‑genin),是合成甾体激素类药物的重要原料。穿龙薯蓣是我国薯蓣属中分布最广、蕴藏量最大的一种,其薯蓣皂苷元平均含量为0.93%~2.26%,是工业化生产最有利用前途的种类之一。近年来,由于生境破坏和大量采挖,穿龙薯蓣野生资源正在减少,而人工栽培的皂苷含量较低,并随着栽培代数增加,种质不断衰退,药材质量问题亟待解决,良种选育已成为保护该种野生资源和解决药材原料供应的首选课题。
穿龙薯蓣繁殖系统复杂,其花序特征具多样性,现有的胚胎学、遗传学和花序类型等方面的研究较少,关于薯蓣科的分类与系统演化也一直存在争议,不利于育种工作的开展。花序是许多花按一定顺序排列的花枝,在空间和时间上控制花朵的开放,可直接决定植物的繁殖成功与否。花序表型的多样性在一定程度上反映了植物功能和进化的意义,可显示植物间的亲缘关系及其系统发育的演化过程,也为花序特征与环境的适应提供线索。
鉴于此,本研究以观察测量为主,结合石蜡制片技术,对穿龙薯蓣雌雄花序的形态特征、数量性状及发育过程等分别进行研究,并根据现有研究资料对其花序类型、结构与功能的相关性、分类学意义等方面进行探讨,旨在为穿龙薯蓣的良种选育、杂交育种、种子产量和质量的提高等工作提供基础资料,也为揭示该种与薯蓣属其他种的亲缘关系及分类地位提供依据。
1 材料与方法
以山西省野外采挖的穿龙薯蓣根状茎为繁殖体,引种栽培于山西农业大学林业站(37°25′N,112°34′E)。林业站位于山西省晋中市太谷区,属暖温带大陆性气侯,春温高于秋温,夏季暖热多雨,冬季长而偏冷,年均气温6~19 ℃;年均降水量为250~620 mm,降雨主要集中分布在7-9月,年均湿度54.5%左右,年均日照时间2 339.9 h。
2020 年5 月至2021 年10 月,随机选取栽培植株的雌雄花序,进行形态观察与拍照。OLYMPUS SZ61 体式显微镜下观察花序,Mshot MS60 摄像头拍照;剔除特殊及发育不良的花序后,随机选取70个雌花序/果序、100 个雄花序,进行花数、果数统计,游标卡尺(0.1 mm)测量长度;采集发育初期的雌雄花序,50%的FAA固定液((50%乙醇)∶(冰乙酸)∶(甲醛溶液)=89∶6∶5)固定,石蜡包埋制片,KEDEE KD-3358 切片机切片,厚度8~10 μm,二甲苯脱蜡,番红—固绿对染法染色,中性树胶封片;OLYMPUS CX31 生物显微镜观察,Mshot MS60摄像头拍照。
2 结果与分析
2.1穿龙薯蓣雌花序的发育与形态结构
穿龙薯蓣雌花花被顶端六裂,浅黄色,具退化雄蕊(图版Ⅰ:A),退化雄蕊具药室(图版Ⅰ:B),但无正常雄蕊的多层药壁结构,不产生花粉且横截面小于正常花药(图版Ⅰ:B~C);子房下位,具3室,每室胚珠2枚(图版Ⅰ:D~E)。
穿龙薯蓣雌花花期6-8 月,果期8-10 月。雌花序平均长度为79.99 mm,最长可达200 mm 以上,最短仅有10 mm。雌花序为单生于叶腋的穗状花序,小花向顶发育(图版Ⅰ:F)。雌花序上平均有11.78朵小花,平均小花密度为0.14朵/mm,但每雌花序平均仅有6.78 枚果实,平均结实率为57.56%。9月之后天气渐冷,此时虽有新雌花及雌花序产生,但未能开放及形成果实(图版Ⅰ:G)。
2.2穿龙薯蓣雄花序的发育与形态结构
穿龙薯蓣雄花花期5-10 月,平均长度为82.68 mm,最长可达300 mm以上,最短仅为11 mm左右,每花序平均58.32 朵小花,小花密度平均为0.66 朵/mm。雄花花被顶端六裂,浅绿色至浅黄色;花药6 枚,着生于花被基部;苞片披针形,短于花被。
穿龙薯蓣的雄花序为混合花序,腋生或于侧枝顶生。花序整体由基部向顶部逐渐发育为小花(图版Ⅱ:A),为向顶发育,花序整体为无限花序;随后各小花的苞片下分别分化出新的小花,新小花与原小花排列成“y”形(图版Ⅱ:A~B),形成侧轴,之后不断交替地从新生的小花苞片下沿花序轴两侧分化出小花,形成小花序(图版Ⅱ:C~G);小花序顶花先开,向基发育,其类型为单歧聚伞花序(图版Ⅱ:E);小花序末端蝎尾状卷曲(图版Ⅱ:F)并沿花序轴排成两列(图版Ⅱ:G),为蝎尾状聚伞花序。穿龙薯蓣雄花序的花序主轴(一级花序)为无限花序,侧轴(二级花序)为蝎尾状单歧聚伞花序,整体属于混合花序,形似穗状。混合花序的发育大致分为3个阶段。发育早期,每个侧轴上仅1~2 朵小花,单歧聚伞花序形态尚不明显,与穗状花序十分相似(图版Ⅱ:H);发育中期,随着花序不断伸长,基部侧轴形成有多朵小花的单歧聚伞花序,顶部侧轴新生,仍仅1~2 朵小花(图版Ⅱ:I~K);发育后期,雄花序生长到一定程度后不再伸长,花序基部与顶部侧轴均形成3朵以上小花组成的单歧聚伞花序(图版Ⅱ:L~N)。一些较短花序的顶部单歧聚伞花序上具有3朵以上小花,并非新生花序(图版Ⅱ:O)。
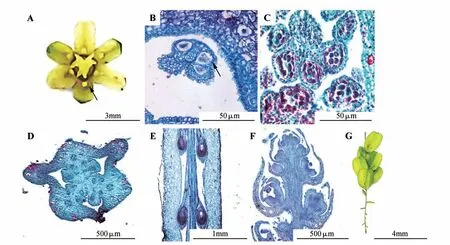
图版Ⅰ穿龙薯蓣雌花与雌花序A.雌花(箭头示退化花药);B.雌花横切面(箭头示退化花药,具有药室但无药壁多层结构,且不产生花粉);C.花药横切面,药壁具多层结构且产生花粉;D.子房横切面(示3 子房室);E.子房纵切面(示每子房室中2 胚珠);F.雌花序纵切面,小花向顶发育;G.果序,顶部新生小花不开放Plate Ⅰ The female flowers and inflorescences of D.nipponicaA.The female flower,arrow shows the vestigial anther;B.Female flower cross section,arrow shows the vestigial anther,has pollen sac but does not multilayer structure of anthers and produce pollen;C.Male flower cross section,the anthers is multilayered and pollens are produced;D.Ova‐ry cross section,showing three ovary chambers;E.Ovary longitudinally section,two ovules per loculus;F.Female inflorescence longitudinally section,flowers develop sequentially from base to top;G.The infructescence,the top flower will not open
此外,存在一些圆锥状的雄花序(图版Ⅲ:A)。这些圆锥状花序基部存在一些远大于苞片的退化叶片(图版Ⅲ:A~B),其基部花序与腋生花序形态相同,为混合花序(图版Ⅲ:A~B);圆锥状花序的侧轴自基部至顶逐渐短缩,直至顶部形成混合花序(图版Ⅲ:A~C),有些圆锥状花序主轴还有明显的缠绕性(图版Ⅲ:D),与具有顶生花序的侧枝生长发育形态一致(图版Ⅲ:E~G)。此外,早期圆锥状花序由基部向顶部不断发育形成花序而并非小花,并伴有叶片形成(图版Ⅲ:H),所以这些圆锥状花序实际为叶片退化的侧枝。
3 讨论
3.1穿龙薯蓣花序的形态特征及功能意义
花序是指许多花按一定顺序排列的花枝,大多数被子植物的花序分为无限花序和有限花序两大类。但仍有部分花序类型介于二者之间,同一花序上同时存在有限与无限2种花序类型,因此它们被称为“混合花序”(mixed inflorescence)。混合花序的概念最早于1949 年由Bailey 定义,2019 年正式收录于第2 版《植物学名词》。1985年版《中国植物志》描述穿龙薯蓣的雄花序为“腋生的穗状花序,基部常由2~4 朵集成小伞状,至花序顶端常为单花;……”。2000 年《Flora of Chi‑na》将穿龙薯蓣雄花序特征补充为:“穗状花序单生,雄花序基部由2~7 朵小花形成小聚伞花序(cy‑mule)或小伞形花序(umbellule),偶有小聚伞花序膨大到形成侧枝,花序上部小花单生,无梗或有短花梗;……”。现今大多数资料仍引用1985年版《中国植物志》的描述。然而穗状花序应为不分枝的无限花序,与穿龙薯蓣雄花序特征不符,也并未提到花序顶生现象;“伞形花序”为无限花序,本研究对雄花序进行了大量解剖,但并未发现此类型;“花序上部小花单生”描述的可能是发育中期的雄花序(图版Ⅱ:I)。在穿龙薯蓣雄花序发育中期,在基部已经形成具有多朵小花的单歧聚伞花序,顶部的单歧聚伞花序顶花还未开放,苞片下新的小花并不明显,呈现为单生花的状态(图版Ⅱ:J~K);底部的单歧聚伞花序可能由于小花数较少,花序轴不明显;另因蝎尾状卷曲,形似簇生或伞形状(图版Ⅱ:J)。

图版Ⅱ 穿龙薯蓣雄花序与蝎尾状聚伞花序A.雄花序纵切面,小花向顶发育,箭头示小花苞片下正在分化出新的小花;B.图A 中小花的放大,小花苞片下正在分化出新的小花;C.雄小花及苞片下新产生的小花;D.具有3朵小花的蝎尾状聚伞花序;E.蝎尾状聚伞花序,小花自苞片下交替产生;F.蝎尾状聚伞花序,侧面蝎尾状卷曲;G.蝎尾状聚伞花序,正面小花排列在花序轴两侧;H.发育早期雄花序;I.发育中期雄花序;J.图I中花序底部的放大;K.图I中花序顶部的放大;L.发育后期雄花序;M.图L中花序顶部的放大;N.图L中花序底部的放大;O.较短雄花序Plate Ⅱ The male inflorescences and cincinnus of D.nipponicaA.Male inflorescence longitudinally section,flowers develop sequentially from base to top,the arrows show new florets differentiating under bracts;B.A magnification of the flowers in Fig.A,new florets differentiating under bracts;C.Male florets and newly formed florets under bracts;D.Cincinnus with three flowers;E.Flowers grow alternately beneath bracts in cincinnus;F.Cincinnus laterally scorpioid curled;G.The presence of cincinnus,flowers arrange on either side of the inflorescence axis;H.Early male inflorescences;I.Medium-term male inflorescences;J.A mag‐nification of the bottom of inflorescence in Fig.I;K.A magnification of the top of inflorescence in Fig.I;L.Latter male inflorescences;M.A mag‐nification of the top of inflorescence in Fig.L;N.A magnification of the bottom of inflorescence in Fig.L;O.Shorter male inflorescencew
单歧聚伞花序(monochasium)是指分枝从苞片腋内生出且每级分枝只有1 个的聚伞花序。这一概念最早在1875 年由Eichler 提出,目前认为单歧聚伞花序有螺状聚伞花序(bostryx/helicoid cyme)、蝎 尾 状 聚 伞 花 序(cincinnus/scorpioid cyme)、扇状聚伞花序(fan/rhipidium)和镰状聚伞花序(drepanium)4 种类型。2015 年,闫瑞亚等对单歧聚伞花序的类型及划分进行了研究综述,穿龙薯蓣雄花序上的侧轴小花序与其描述的蝎尾状聚伞花序特征一致,即分枝在轴的两侧交替发生,排成两列,花序末端蝎尾状卷曲。除腋生花序外,穿龙薯蓣雄花序还于侧枝顶生;另有部分花序呈圆锥状,圆锥状花序与侧枝生长发育状态一致,所以这些圆锥状花序实际是由多个花序聚在一起而成的花枝,即叶片退化的侧枝(图版Ⅲ:A~D)。此外,由于侧枝顶端分生组织逐渐由营养生长转变为生殖生长,叶片退化,枝条逐渐转变为花序,最后形成顶生花序,与侧枝腋生花序共同组成圆锥状花序(图版Ⅲ:A~D)。

图版Ⅲ 穿龙薯蓣雄花序的多样性A.圆锥状的雄花序;B.图A 中花序底部的放大,箭头示退化叶片;C.图A 中花序顶部的放大;D.具有缠绕性的圆锥状雄花序;E.侧枝顶生花序;F.图E中花序顶部的放大;G.侧枝中部短缩的雄花序;H.早期圆锥状雄花序纵切面Plate Ⅲ Diversity of male inflorescences in D.nipponicaA.Male inflorescences resembling panicle;B.A magnification of the bottom of inflorescence in Fig.A,arrows show the degraded leaves;C.A magnification of the top of inflorescence in Fig.A;D.Male inflorescences resembling panicle and twining;E.Terminal inflorescence on lateral branches;F.A magnification of the top of inflorescence in Fig.E;G.Male inflorescence of lateral branches midsection short;H.Early male inflo‐rescences resembling panicle longitudinally section
花是植物器官中进化水平最高、构造最为复杂的生殖器官,在植物延续后代过程中起着关键的作用。花序作为功能单位,其结构在很大程度上受到自然选择的影响。穿龙薯蓣雄花序是侧轴为蝎尾状单歧聚伞花序的无限花序,即混合花序,使得雄花序上花的密度和数量远高于雌花。另外,雄花序的平均长度大于雌花序,雄花整体花期也比雌花长约2个月,这些特征皆有利于植物传粉。穿龙薯蓣野生状态下,常生于林下,缠绕于乔木或灌木上,光照不足。雄株兼具顶生花序和腋生花序,较之单纯的腋生或顶生花序,生殖功能的整体分布更均匀,也更适应林下环境,利于进行异花授粉。此外,无限花序的植物可根据气候调整花的数量,更能适应干旱半干旱气候。
3.2穿龙薯蓣花序特征的分类学意义
自20世纪初,人们对薯蓣科植物从形态学、孢粉学、基因组学等多方面进行研究,发现其虽为单子叶植物,但具有双子叶植物的许多特征,关于薯蓣科的分类地位,科内、属内分组及是否与双子叶植物相联系一直是植物系统学上争论的问题。在早先常用的恩格勒分类系统中,薯蓣科位于百合目之下;1998 年发布的APG 系统将薯蓣科从百合目中分离出来,单独列为薯蓣目。2003 年发布的APG Ⅱ系统将薯蓣目调整合并为薯蓣科、水玉簪科(Burmanniaceae)和纳茜菜科(又称沼金花科)(Nartheciaceae)3科,薯蓣科内下分蒟蒻薯属()、多子薯蓣属()、丝柄花属()及薯蓣属4 属;2016 年发布的APG Ⅳ系统暂时沿用此分类,但仍考虑将蒟蒻薯属和水玉杯属独立为蒟蒻薯科(Taccaceae)及肉质腐生科(Thismiaceae)。薯蓣属内的分组也存在颇多争议,我国薯蓣属植物分为5~8 组,本研究以《中国植物志》的6 组分法作为参考。分组虽有争议,但普遍认为穿龙薯蓣所在的根状茎组为薯蓣属中演化最原始的一组。
花序是物种描述的重要表型性状,在分类学研究和系统发育研究中起着重要的作用。目前,主流观点认为被子植物最原始的花序类型是具叶的聚伞花序,可简化为单花或演化为无限花序;其中一种无限花序的起源途径为:通过顶生花的初期抑制形成过渡状态的聚伞圆锥花序(thyrse/thyrsus),随之每个侧轴的花简化到一朵,形成总状花序。聚伞圆锥花序是混合花序中最为常见的类型,其主轴为无限花序,侧轴为有限花序,外观与圆锥花序高度相似,如丁香属()植物、玄参属()植物等。
2008 年,Remizowa 等对薯蓣目纳茜菜科植物花发育进行解剖学研究,认为该科植物花序类型为总状花序。2021年,Nuraliev 等对薯蓣目肉质腐生科(初版APG 系统,后归为薯蓣目水玉簪科)水玉杯属植物的开花特征进行了研究,认为该属几种植物的花序类型为单歧聚伞花序。而薯蓣科内植物花序类型多样,蒟蒻薯属、多子薯蓣属植物被认为拥有与石蒜科植物相似的聚伞花序,丝柄花属植物多单生或为聚伞花序。就薯蓣属植物的花序特征而言,雌花序为腋生的穗状或总状花序,基本无争议,雄花序却具多样性:《中国植物志》描述其为穗状、总状或圆锥花序;《Flora of China》补充毡毛薯蓣()、云南薯蓣()等的雄花序为聚伞圆锥花序,马肠薯蓣()、纤细薯蓣()等的花序基部具有小聚伞花序。在周生翅组(Sect.)内,圆锥状雄花序是区分日本薯蓣()及其他几种的重要特征。2010 年,Remizowa 等利用扫描电子显微镜对根状茎组的山萆薢()花序发育进行研究,认为山萆薢雄花序是侧轴为蝎尾状单歧聚伞花序的无限花序,这与穿龙薯蓣雄花序特征相似。因此,穿龙薯蓣、山萆薢等植物的雄花序特征为有限花序向无限花序演化的中间状态,证明薯蓣属植物花序很可能发生于此途径,薯蓣目3 科及薯蓣科4属植物的花序特征也与此相符。
植物为了获得更高频的远系杂交能力和最大程度的选择优势,由雌雄同花祖先经历至少100次的独立进化形成单性花。在一些物种中,单性花在发育初期是两性的,后来随着雄蕊或雌蕊停止发育变成了单性花,因此存在没有功能的退化器官,同一科不同植物之间也存在器官停止发育阶段的差异。吴宝成等认为,薯蓣科内除薯蓣属外的其他3 属均为两性花,而薯蓣属内,演化程度较高的基生翅组(Sect. Opsophyton)的黄独(),其雌花虽具雄蕊,但雄蕊退化成半透明的杆状突起;较原始物种的退化雄蕊形态上接近正常雄蕊,如根状茎组的马肠薯蓣、盾叶薯蓣()等雌花中的退化雄蕊具有完整的花丝、花药和花粉,且花粉具有一定萌发率;而穿龙薯蓣雌花的退化雄蕊具有花粉囊,但结构不完整,且不产生花粉,证明其在薯蓣属中处于较为原始的过渡地位。
闫瑞亚等指出目前国内一些教材和植物志等对聚伞花序类型及其概念存在混淆和较为模糊的表述。《中国植物志》及《Flora of China》对粘山药()、柔毛薯蓣()、毡毛薯蓣等的雄花序描述为“雄花数朵组成小聚伞花序,再排列成穗状花序”,并未具体描述聚伞花序的类型,亦将雄花序描述为穗状花序。《中国植物志》及《Flora of China》描述复叶组(Sect.Lasiophyton)、根状茎组、周生翅组、基生翅组等均存在部分雄花序呈圆锥状,这些圆锥状的花序是否也是叶片退化的侧枝,以及是否存在顶生花序有待进一步明确。此外,山萆薢雄花有1.7 mm 左右的花梗,因此雄花序被描述为圆锥聚伞花序,但穿龙薯蓣雄花普遍无梗或偶有短梗,不能被称作圆锥聚伞花序,故本文仅描述其为混合花序。
4 结论
本研究发现,穿龙薯蓣雌雄花序的形态及数量特征能更好保证传粉的成功率,增强对环境的适应性;穿龙薯蓣及其亲缘植物花序类型与有限花序向无限花序演化过程相符,具原始性;雄花序特征与现有资料的描述不尽相同,且无法以现有花序名词准确定义,本文仅较为粗略地称其为混合花序。自然界中的花序式样实际上远多于人类已知的类型,需不断研究并更正。因此,继续深入研究并明确薯蓣科植物的花序类型和花序特征,不仅有利于栽培与繁殖工作的进行,更有助于其分类地位的研究、科内与属内的分组演化、亲缘关系的探讨及植物花序类型的划分。
