虎耳草属异叶虎耳草复合群的数量分类学研究
2022-08-04马小磊贾留坤陈世龙高庆波
马小磊 贾留坤 曹 倩 陈世龙 高庆波*
(1. 中国科学院西北高原生物研究所青海省作物分子育种重点实验室,西宁 810008;2. 中国科学院大学,北京 100049;3. 中国科学院昆明植物研究所中国科学院东亚植物多样性与生物地理学重点实验室,昆明 650201)
虎耳草属(Tourn.ex L.)是虎耳草科(Saxifragaceae)最大的属,该属主要分布在北半球高山地区,全世界约有500 种,其中约有216 种分布于中国,139 种为中国特有。山羊臭组(sect.Haw.)是虎耳草属中最大的一个组,拥有极高的物种丰富度,共有175 种,中国分布有166种,其中112 种为我国特有。国内外多位学者基于形态学证据对虎耳草属的分类做了详细研究,发表了很多新种并对分类系统做了修订,但山羊臭组组下分类一直未能被解决。Gornall在1987年提出的分类系统中,将山羊臭组分为7 个亚组;潘锦堂先生于1991 年将中国的虎耳草属归入4 个亚组;《Flora of China》(下称FOC)中,Pan等未对国内的山羊臭组进行亚组划分,而是依据形态学特征分为7 个Key,并分别编写检索表。近年来,多位学者基于形态学和分子生物学证据得出一致的结果,即确定了山羊臭组是一个单系类群(但组内关系仍未解决),且支持将山羊臭组分为3 个亚组,其中唐古拉亚组(subsect.Engl.)单独聚在一个分支上,构成一个单系类群。
唐古拉亚组的分化时间短,辐射演化导致类群内部的遗传分化非常小,基于相对有限的分子生物学证据无法有效地建立亚组内的系统发育关系。基于形态差异,可将唐古拉亚组分为山地虎耳草复合群(complex)、异叶虎耳草复合群(complex)和狭瓣虎耳草复合群(complex),分别对应FOC 山羊臭组分种检索表Key3、Key5 和Key6。异叶虎耳草复合群有29 个种和4 个变种被FOC 收录并描述,4 个物种和1 变种被收录但无描述,此外还有部分新种未被收录或处理。异叶虎耳草复合群的共有特征为叶片灰绿色,叶缘具有突起的叶脉从基部延伸到先端。基于虎耳草属叶脉模式的研究表明,叶缘具有突起的叶脉从基部延伸到先端的特征只在异叶虎耳草复合群中出现,暗示该类群可能有共同的起源。
数量分类学是具有一定效度的一种分类学研究方法,可以为完善分类系统的类群划分提供参考,近年来仍被许多学者用于解决疑难类群的分类学问题,也被用于研究属内杂交以及种内变异。本研究基于国内各大标本馆馆藏的虎耳草属异叶虎耳草复合群物种标本,利用聚类分析和主成分分析等方法,分析复合群内物种间的形态差异,探讨异叶虎耳草复合群的种间关系,并结合地理分布式样分析其分布格局,进行类群划分,以期为进一步探明唐古拉亚组系统发育关系提供证据。
1 材料与方法
通过对中国科学院植物研究所标本馆(PE)、昆明植物研究所标本馆(KUN)、西北高原生物研究所青藏高原生物标本馆(HNWP)及青藏高原植物适应与进化课题组该类群标本进行查阅,共统计标本1 192 份,包括31 个种、2 个变种,每份标本统计42 个性状。所有标本均提取采集信息,并依据中国数字植物标本馆(CVH)的采集记录进行信息增补。分类性状选择参考《中国植物志》《Flora of China》《西藏植物志》《云南植物志》,对于部分缺失性状的补充,优先选取同一采集号标本、同一采集地标本或同一物种不同个体间出现次数最多的性状。
对标本信息进行数字化过程中,舍弃部分鉴定有较大争议、信息有大量缺失或有关键性状缺失的标本,最终选定657 份标本进行后续分析,包括24个种、2个变种(见表1)。对42个性状进行甄别,排除种内存在较大变异的性状和判别过程主观性过强的性状,最终选定24 个分类性状用于后续分析(见表2)。使用IBM SPSS Statistics 26 软件(下称SPSS)进行主成分分析(Principal Component Analysis,PCA),并以每个物种作为运筹分类单元(OTUs),基于平方欧氏距离使用组间平均法进行Q 型聚类分析。以每个物种作为OTUs 聚类时,性状选取每个物种标本中出现次数最多的性状,部分性状无法判别时,则参考物种描述选定,选定的性状用SPSS对赋值标准化后进行聚类分析。

表1 虎耳草属异叶虎耳草复合群物种标本记录Table 1 Specimen records of S.diversifolia complex

表2 异叶虎耳草复合群分类特征及编码Table 2 Taxonomic characteristics and encoding of S.diversifolia complex
对标本采集信息进行整理和修订。对于标本上出现的小地名,由于历史原因有变更的地名,采集信息为英文、采集地采用音译拼写方法的地名均参考中国早期标本采集地名考数据库和《中国地名演变手册(1912 年以来省市县新老地名)》进行人工校正;对于无采集地信息、采集地范围过大、采集信息模糊等无法确定采集地点的标本记录予以舍弃。整理获得的采集信息以县级单位进行合并,共得到443 份地理分布信息,涵盖青海省和甘肃省、陕西省、四川省、湖北省、云南省、西藏自治区93个县和尼泊尔一处采集记录,包括异叶虎耳草复合群30 个种和1 个变种。通过Google Earth和百度地图开放平台坐标拾取功能查询分布地的经纬度坐标信息,使用ArcGIS 10.2 软件绘制异叶虎耳草复合群物种分布图。
2 结果与分析
2.1主成分分析
通过主成分分析法(PCA)提取7 个特征值大于1 的主成分,累计贡献率为67.748%(见表3)。第1 主成分贡献率为17.842%,第2 主成分贡献率为16.709%,第3 主成分贡献率为10.721%,前3 个主成分累计贡献率为45.272%。依据主成分分析中主成分负载量(见表4),第1 主成分中“基生叶叶基心形”“中下部茎生叶大小”“花序形态”“中下部茎生叶基部心形”“花梗被毛长度”“中下部茎生叶形态”的绝对权重比较高,分别为0.788、0.769、0.673、0.644、0.633、0.625;第2主成分中“萼片脉纹于先端汇合”“中下部茎生叶是否具柄”“存在茎生叶基部抱茎”“上部茎生叶基部心形”的绝对权重较高,分别为0.803、0.781、0.730、0.692;其余主成分中,“中下部茎被毛情况”“顶端茎生叶被毛情况”的绝对权重较高,分别为0.601、0.653。

表3 异叶虎耳草复合群主成分分析的特征值和贡献率Table 3 Eigenvalues and percent of PCA of S. diversifo‐lia complex

表4 异叶虎耳草复合群形态性状主成分分析前3个主成分负载量Table 4 Eigenvector of the first three greatest principal component in PCA of S.diversifolia complex
2.2地理数据分析
根据异叶虎耳草复合群的物种分布图(见图1),发现异叶虎耳草复合群拥有3 个多样性中心,喜马拉雅山地区、环四川盆地高山地区,以及横断山地区。排除广布种的干扰后发现3 个分布中心之间有较明显的地理间隔,仅少数物种(、等)在喜马拉雅山地和横断山地区2 个多样性中心均有分布。位于横断山的3 个县(市)和香格里拉县境内采集到12 种、丽江市境内采集到10 种、木里县境内采集到10 种,拥有相较于其他地区更高的物种多样性。
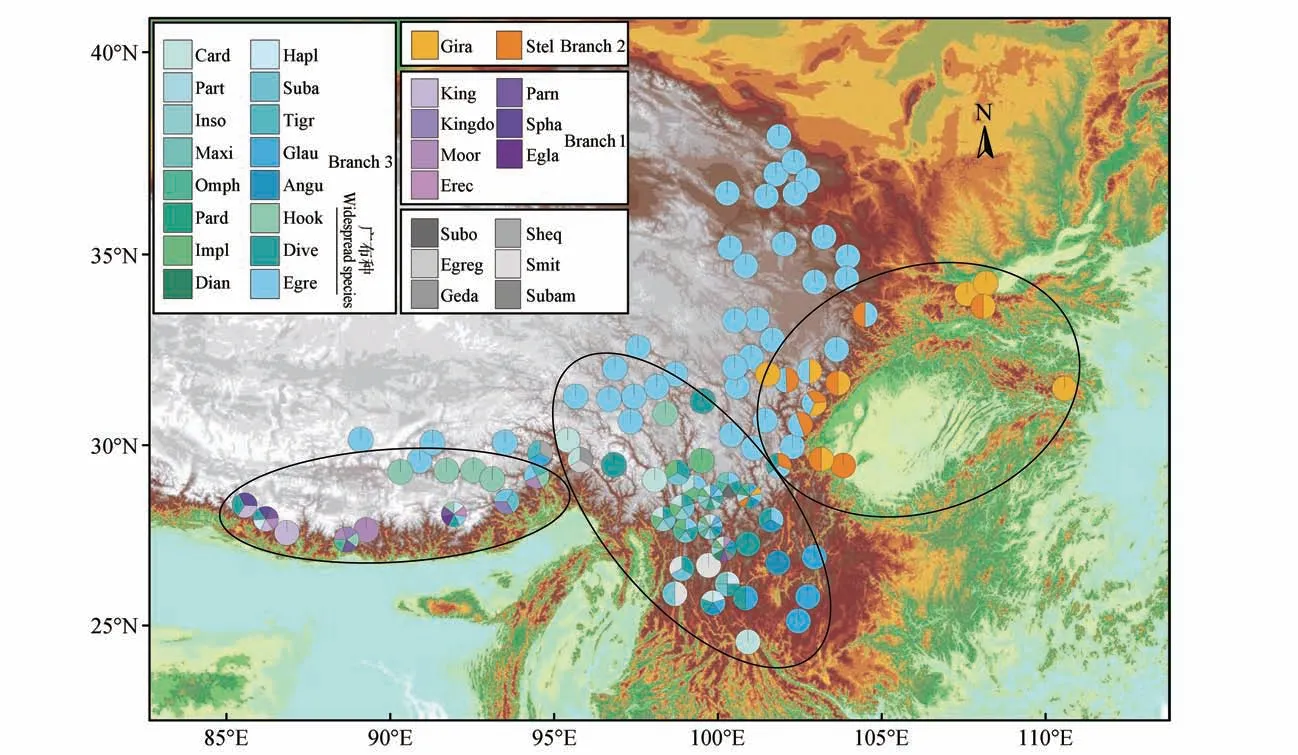
图1 异叶虎耳草复合群的物种分布Branch 1.喜马拉雅山南坡分支;Branch 2.环四川盆地山区分支;Branch 3.横断山分支;图中每个物种以不同颜色标注,每个饼图代表一个县级单位的异叶虎耳草复合群物种多样性;黑色椭圆表示3个物种多样性中心;灰色物种表示其属于异叶虎耳草复合群,有标本采集信息但未参与聚类,其中Subo代表S.subomphalodifolia J.T.Pan;Smit代表S.smithiana Irmsch.;Egreg代表S.egregioides J.T.Pan;Sheq代表S.sheqilaensis J.T.Pan;Geda代表S.gedangensis J.T.Pan;Subam代表S.subamplexicaulis Engl.et IrmschFig.1 Species distribution of S.diversifolia complexBranch 1.Himalayan Branch;Branch 2.Mountains Around Sichuan Basin Branch;Branch 3.Hengduan Mountains Branch;Species are color-coded.Each pie chart represents species diversity at county level.Black ellipses represent the three centers of species diversity.Gray sectors represent spe‑cies that belong to S.diversifolia complex but not included in our clustering.Where Subo is represents S.subomphalodifolia J.T.Pan,Smit is repre‑sents S.smithiana Irmsch.,Egreg is represents S.egregioides J.T.Pan,Sheq is represents S.sheqilaensis J.T.Pan,Geda is represents S.gedangensis J.T.Pan,Subam is represents S.subamplexicaulis Engl.et Irmsch
2.3聚类分析
以每个物种作为OTUs 进行聚类,对比经典分类结果和地理分布,以平方欧氏距离等于12.5 为划分线,可将异叶虎耳草复合群分为6 个分支(见图2):分支A 仅一个种;分支B 包括和;分支C 仅一 个 种;分支D 包 括、S.和;分支E 包括和;分支F 包括其余的15 个种和2个 变种。结合地理分布式样,分支A、B、C、D 均分布于喜马拉雅山地区,分支E 则分布于环四川盆地的高山环境,分支F 除广布种外,主要分布在横断山地区。因此,可将异叶虎耳草复合群分为3 个分支,即喜马拉雅山南坡分支、环四川盆地山区分支,以及横断山分支,其中横断山分支包含一个广布种。参考碎石图,选择前2个主成分绘制二维坐标空间,用每份标本因子得分绘制散点图(见图2),将657 个OTUs 按所在分支用不同颜色标注在散点图中,依照形态聚类结果和地理分布式样区分的3 个分支分离情况良好。

图2 异叶虎耳草复合群物种主成分分析散点图(A)和形态学聚类结果图(B)Branch 1.喜马拉雅山南坡分支;Branch 2.环四川盆地山区分支;Branch 3.横断山分支Fig.2 Scatter plot of PCA and morphological clustering results of S.diversifolia complexBranch 1.Himalayan Branch;Branch 2.Mountains Around Sichuan Basin Branch;Branch 3.Hengduan Mountains Branch
3 讨论
主成分分析中,特征值大于1 的前7 个主成分的累计贡献率较小,说明山羊臭组植物在演化过程中的性状变异较小,无明显的规律性,这可能和山羊臭组物种是较晚演化出的类群,并在青藏高原地区经历快速辐射和多次扩散事件有关。虎耳草属物种多样化和喜马拉雅—横断山地区气候变化以及地理环境等因素密切相关,而气候作用下形成的宽松生态位和高的生态位进化速率造就了唐古拉亚组的物种的快速多样化。依据主成分分析结果,传统分类中认为是关键性状的“基生叶花期凋落”“花瓣是否具斑点”“花梗被毛情况”等对所有主成分的贡献值较低;检索表中使用次数较多的“萼片被毛情况”和“花萼是否反折”在前3个主成分中的贡献值并不高,这些性状的分类价值在以往分类系统中或许被高估了。根据对标本观察,“基生叶花期凋落”受标本采集者的采集习惯以及标本制作方法的影响较大;“花瓣是否具斑点”在标本中难以观察。在一些物种中,“萼片是否被毛”(例如,和)和“萼片是否反折”(例如,)并非稳定性状。对标本观察发现和在“中下部茎生叶被毛情况”、“花萼是否反折”“花萼被毛情况”“萼片边缘是否具睫毛”“花梗被毛情况”等方面均有较大变异,两个物种的关键区别是萼片是否被毛,但这显然不是一个稳定性状。传统分类中认为重要的性状,如中下部“茎生叶是否具柄”“中下部茎生叶形态”“花序类型”“中下部茎生叶大小”“萼片脉纹于先端汇合”“叶基心形”等,在前3 个主成分中有较高的贡献值,本研究肯定了其作为关键分类性状的合理性。依据这些性状可以对3个分支进行区别:喜马拉雅山南坡分支的共有特征为植株高大,上部叶片无柄,叶片心形,近抱茎,上下部叶片近同形,花序多为聚伞花序;横断山分支的主要特征为茎上部叶片多少具柄,茎上部叶片椭圆形或披针形,叶基楔形,极少心形,大部分物种上部叶片明显小于下部叶片;环四川盆地山区分支与其他分支的主要区别特征为具有相较于其他分支更低的植株高度,叶片为三角形或三角状卵形,并且小于其他分支,花序具单花或仅有少量花,这可能和其分布在高山草甸有关。从喜马拉雅山南坡分支到横断山分支再到四川盆地分支表现出植株大小和叶片大小逐渐变小的趋势。
聚类分析的部分结果和前人的研究一致,和聚在一个分支(分支B),、和聚为一支(分支D),两个分支在潘锦堂1991 年的系统中分别隶属于小泡系(ser.J.T.Pan)和春丕系(ser.J.T.Pan);和聚为一支(分支E),该分支叶片边缘突起的叶脉并没有从基部贯穿到先端,和其他分支有明显区别。结合聚类分析结果和分布式样发现,异叶虎耳草复合群的物种多样性分布格局与于海彬等对青藏高原特有种子植物多样性分布格局的研究一致,该复合群物种在喜马拉雅—横断山脉的物种多样性最为丰富。横断山分支由15 个种和2 个变种组成,占所研究的异叶虎耳草复合群物种的三分之二,该分支的物种集中分布于大雪山、梅里雪山和贡嘎山之间的区域。聚类结果显示,环四川盆地山区分支与喜马拉雅山南坡分支的关系更近一些,这与地理分布不符,笔者推测3 个分支的祖先类群在较早的时间就已经扩散到3个多样性中心分范围,随后才发生适应性辐射,由于横断山独特的气候和地理条件,使得横断山分支的物种快速辐射,拥有更多的特有性状,而另外两个分支则保留了较多的祖系类群的特征。优越虎耳草是异叶虎耳草复合群中的一个广布种,形态性状聚类显示其应该属于横断山分支。贾留坤等对优越虎耳草进行了谱系地理学研究,发现该物种居群没有大规模扩散信号。结合异叶虎耳草复合群的分化时间,笔者推测优越虎耳草可能是从横断山附近起源,首先扩散到与现在大致相同的分布范围,后由于冰期等多种因素的影响,形成彼此隔离的避难所,随后又发生原地扩散形成现在的分布式样。
虎耳草属是青藏高原上特有种最多的属之一,对青藏高原生物多样性研究和保护均具有很高的价值。分类学是生物学研究最基础的工作之一,对异叶虎耳草复合群进行分类学研究,能够为该类群物种的其他研究工作提供便利。对异叶虎耳草复合群进行数量分类学的研究虽不足以用于分类系统修订,但能基于大量数据得出一个具有最大预测性的纯表征分类,为进一步探究和理解唐古拉亚组内部系统发育关系提供思路。
