非洲猪瘟病毒pH359L蛋白原核表达及多克隆抗体的制备
2022-07-25杨赛霞赵亚茹刘志杰杨吉飞张忠辉耿抒贤吕欣倩敬梦瑶牛庆丽
杨赛霞,赵亚茹,刘志杰,杨吉飞,张忠辉,樊 洁,耿抒贤,吕欣倩,敬梦瑶,殷 宏,2,牛庆丽*
(1.国家非洲猪瘟区域实验室(兰州)/中国农业科学院 兰州兽医研究所 家畜疫病病原生物学国家重点实验室,甘肃 兰州 730046;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,江苏 扬州 225009)
非洲猪瘟 (African swine fever,ASF) 是由非洲猪瘟病毒 (African swine fever virus,ASFV) 感染引起的一种急性、热性、高度接触性传染病,该病毒感染后致死率高达100%,目前尚无有效的疫苗和治疗药物。ASFV感染家猪和野猪,软蜱科中的钝缘蜱属(Ornithodoros)是ASF的储藏宿主和传播媒介[1]。该病引起的临床症状主要包括高热、发绀、厌食症和共济失调,甚至死亡。在急性和亚急性的ASF感染中,不同器官均表现出严重的出血,如肾瘀点和淋巴结的弥漫性出血、血管内凝血、肺水肿和血小板减少等[2]。ASF在我国首次暴发于2018年8月[3],截至2021年1月,已在31个省、市、自治区发生和流行,给我国生猪产业造成了严重损失[4]。世界动物卫生组织 (World Organization for Animal Health,OIE)将ASF列为必须通报动物疫病,我国将其列为重点防范的一类动物传染病。
AFSV是一种大的有囊膜的病毒,呈二十面体对称,平均直径约为200 nm,与环烯病毒、痘病毒具有相似的基因组和结构特征,均属于大的核质类DNA病毒(nucleoplasmic large DNA virus,NCLDV)[5]。病毒基因组为末端共价闭合的双链DNA,大小在170 ~190 kb之间,编码151~167个开放阅读框[6]。ASFV主要感染猪单核巨噬细胞,并在细胞中增殖;根据编码主要衣壳蛋白p72的基因(B646L)C端序列变异的遗传特征,ASFV可分为24种基因型[7],序列比对分析发现,我国流行的毒株与俄罗斯和中东欧国家流行的毒株均属于基因Ⅱ型[8]。
真核生物的转录有3种聚合酶(RNA聚合酶Ⅰ、Ⅱ、Ⅲ)和3种不同的启动子以及相关元件。RNA聚合酶Ⅱ由8~14个亚基组成,其自身不能单独启动转录,通用转录因子(TFIIB,TFIID,TFIIE,TFIIF和TFIIH)通过聚合酶Ⅱ组装在启动子DNA上,和定位因子TATA结合蛋白 (TATA binding protein TBP,TFIID的一个亚基) TBP共同形成精确起始转录的大型多蛋白质-DNA复合物[9]。
NCLDV具有相当复杂的RNA聚合酶。研究表明,与真核生物RNA聚合酶Ⅱ编码的14个亚基相比,ASFV编码了同源于真核RNA聚合酶Ⅱ亚基的RPB1(Pol Ⅱ的亚基)、2、3、5、6、7、10、TFIIB和TFIIS共9个亚基[10],有研究推测NCLDV在复制循环过程中,RNA聚合酶亚基基因存在缺失,因此可能会利用宿主RNA聚合酶亚基和通用转录因子,以提高它们在宿主细胞质内的转录效率[11]。TFIIB是构成RNA聚合酶Ⅱ启动复合物的几种通用转录因子之一。在真核生物细胞中,RPB3是RNA聚合酶Ⅱ的核心亚基,与RPB11亚基一起形成异二聚体,被认为是启动子识别中涉及的细菌α亚基同型二聚体的功能对偶物。RPB1、RPB2、RPB3和RPB11主要负责Pol Ⅱ的RNA催化[12]。研究表明,ASFV H359L基因与真核生物RPB3具有高度的相似性[10],因此,pH359L蛋白可能具有与ASFV转录相关的生物学功能。
目前国内外对ASFV pH359L蛋白在ASFV基因复制和转录过程中的功能尚未见报道。因此,本研究利用大肠杆菌原核表达系统表达ASFV pH359L蛋白,在其基础上制备多克隆抗体,为研究ASFV pH359L蛋白的功能和探索ASFV转录机制提供生物材料。
1 材料与方法
1.1 毒株、细胞、载体和实验动物猪肺泡巨噬细胞(PAMs)、SPF猪血清和ASFV阳性血清、基因组及ASFV(CN/GS/2018)毒株均由国家ASF区域实验室(兰州)保存,并在中国农业科学院兰州兽医研究所生物安全三级实验室中在PAMs中扩增。Trans 5α感受态细胞、BL21 (DE3)感受态细胞购自北京全式金生物技术有限公司;引物由生工生物工程(上海)股份有限公司合成;原核表达载体 pET-28a(+)由本试验保存;健康猪购自康乐县磊鑫生猪养殖有限责任公司;2月龄SPF级雄性新西兰大白兔购自中国农业科学院兰州兽医研究所动物实验中心。
1.2 主要试剂Q5 Hot start High-Fidelity 2× Master Mix、限制性内切酶NdeⅠ、EcoRⅠ和T4DNA连接酶购自NEB公司;质粒提取试剂盒、胶回收试剂盒购自OMEGA公司;IPTG购自生工生物工程(上海)股份有限公司;镍预装柱购自金斯瑞生物科技有限公司;弗氏完全佐剂、弗氏不完全佐剂、HRP标记的山羊抗兔及抗猪二抗均购自Sigma公司;预染蛋白Marker、超敏化学发光显色液及RIPA裂解液购自Thermo Fisher Scientific有限公司;RPMI 1640细胞培养基和胎牛血清(FBS)购自Gibco公司。
1.3 H359L基因扩增及重组质粒构建根据GenBank登录的ASFV HLJ/18 (MK_333180) 毒株H359L基因序列(Gene ID:QBH90556)设计特异性引物:H359L-F:5′-CGCCATATGGAAAAAAT-TTTCCAAAACGTGGAAATCAAAC-3′,H359L-R:5′-CCGGAATTCTTAAGCAATCAGTTCATCAACATTTTTTTCAAGAATTTG-3′,在上、下游引物的5′端分别加入EcoRⅠ和NdeⅠ酶切位点(下划线)。采用PCR方法对H359L基因进行扩增,反应体系为(50 μL):Q5 Hot start High-Fidelity 2×Master Mix 25 μL,上、下游引物(10 μmol/L)各2.5 μL,ASFV基因组2 μL,ddH2O 18 μL。PCR反应程序:98℃预变性30 s;98℃变性10 s,55℃退火30 s,72℃延伸30 s,共35个循环;72℃延伸2 min,4℃保存。通过琼脂糖凝胶电泳鉴定PCR扩增产物的大小,用胶回收试剂盒纯化回收。用Nde Ⅰ和EcoR Ⅰ分别酶切目的基因片段和pET-28a(+)表达载体,用胶回收试剂盒纯化回收酶切产物,经T4DNA连接酶连接目的基因和表达载体,并转化至E.coliTrans5α感受态细胞,涂布LB平板(卡那青霉素抗性),挑取单克隆,扩大培养后提取质粒进行双酶切鉴定,鉴定正确的单克隆送至生工生物工程(上海)股份有限公司测序,将酶切和测序均正确的单克隆经菌液培养后提取质粒(pET-28a-H359L)于-20℃保存备用。
1.4 pH359L重组蛋白的诱导表达及纯化将重组表达质粒pET-28a-H359L转化至E.coliBL21(DE3) 感受态细胞,挑取单克隆接种于3 mL LB(卡那青霉素抗性)液体培养基过夜培养,取1 mL过夜表达的菌液接种到1 L LB培养基,37℃,200 r/min 扩大培养至D600 nm到0.6~0.8,加入终浓度为0.1 mmol/L的IPTG,37℃诱导表达6 h;12 000 r/min 离心15 min,收集菌体沉淀进行超声破碎,4℃,12 000 r/min离心15 min,分别收集上清和沉淀,SDS-PAGE分析蛋白表达形式。镍柱层析法纯化目的蛋白,纯化完成后分别用6,4,2,0 mol/L尿素透析,透析时间分别为4,6,8,2 h;重组蛋白经SDS-PAGE检测及浓度测定后分装,-80℃保存备用。
1.5 重组蛋白的反应性鉴定纯化后的pH359L重组蛋白经SDS-PAGE电泳后半干法转印至PVDF膜,加入5%脱脂乳室温封闭1 h;分别将ASF阳性血清和阴性血清1∶500稀释后4℃孵育,过夜,TBST洗膜5次,每次5 min;辣根过氧化物酶 (HRP) 标记的山羊抗猪IgG 1∶5 000稀释后室温孵育1 h,TBST洗膜5次,每次5 min;利用ChemiDoc XRS(BIO-RAD)凝胶成像系统分析重组蛋白与ASF阴、阳性血清的反应性。
1.6 兔抗pH359L蛋白多克隆抗体的制备将纯化的pH359L重组蛋白(300 μg/只)与相同剂量的弗式完全佐剂乳化后,采用背部皮下多点注射的方法首次免疫新西兰大白兔,每间隔2周用150 μg蛋白和等体积弗式不完全佐剂乳化加强免疫;三免后,通过耳缘静脉采血并收集血清,于-20℃分装保存。
1.7 多克隆抗体效价的测定采用间接ELISA方法测定制备的抗ASFV pH359L蛋白多克隆抗体的效价。采用0.4 μg/孔pH359L蛋白包被ELISA反应板,4℃过夜;PBST洗涤3次,用2% BSA,37℃封闭1 h;PBST洗涤3次,将血清以1∶1 000~ 1∶512 000倍比稀释,100 μL/孔,加入到反应板中,于37℃孵育1 h;PBST洗涤3次,100 μL/孔加入1∶5 000 稀释的HRP标记的山羊抗兔IgG,37℃孵育1 h;PBST洗涤3次,每孔加入100 μL TMB,37℃避光显色5 min,每孔加入100 μL H2SO4(2 mol/L)终止反应;酶标仪测定D450 nm值,根据P/N≥2.1计算抗体效价。
1.8 多克隆抗体的Western blot检测ASFV(MOI=0.2)感染PAM后48 h,同时用未感染ASFV的PAM作为对照组,分别收集感染和未感染的细胞样品,于4℃,1 000 r/min离心10 min,PBS洗涤1次。用含1%蛋白酶抑制剂的RIPA裂解液冰上裂解细胞沉淀30 min,4℃,12 000×g/min离心30 min,取上清进行蛋白浓度测定,加入2×Loading buffer 100℃煮沸10 min。经SDS-PAGE电泳后转印至PVDF膜,制备的多克隆抗体1∶200稀释后加入PVDF膜4℃孵育过夜;洗膜后加入1∶5 000 稀释的HRP标记的山羊抗兔IgG室温孵育1 h;利用ChemiDoc XRS(BIO-RA)凝胶成像系统分析制备的抗体与ASFV天然蛋白的反应性。
1.9 间接免疫荧光试验(IFA)ASFV(MOI=100)感染PAM 24 h后弃上清,PBS洗2次,用4%多聚甲醛室温固定10 min,PBS洗5次,每次10 min;用0.3% TrionX-100透膜15 min,PBS洗5次,每次10 min;用5% BSA封闭2 h后,加入以5% BSA稀释的兔抗pH359L(1∶50)多克隆抗体,于4℃孵育过夜,PBS洗5次,每次10 min;FITC标记的山羊抗兔的二抗1∶200稀释后室温避光孵育1 h,PBS洗5次,每次10 min;1 mg/L的 DAPI染色1 min,PBS洗5次;加入少量PBS,置于激光共聚焦显微镜下观察结果。
2 结果
2.1 H359L基因扩增及重组质粒的构建将PCR扩增产物经1%琼脂糖凝胶电泳分析,检测到与预测大小一致的单一条带(图1A)。将目的基因连接到pET-28a载体后,对构建的pET-28a-H359L重组质粒进行双酶切鉴定,结果获得1 089 bp和pET-28a载体片段2条带(图1B)。挑取阳性克隆送至生工生物工程(上海)股份有限公司测序,测序结果与GenBank 登录的H359L基因序列相似性为100%,无点突变,表明成功构建了重组质粒。
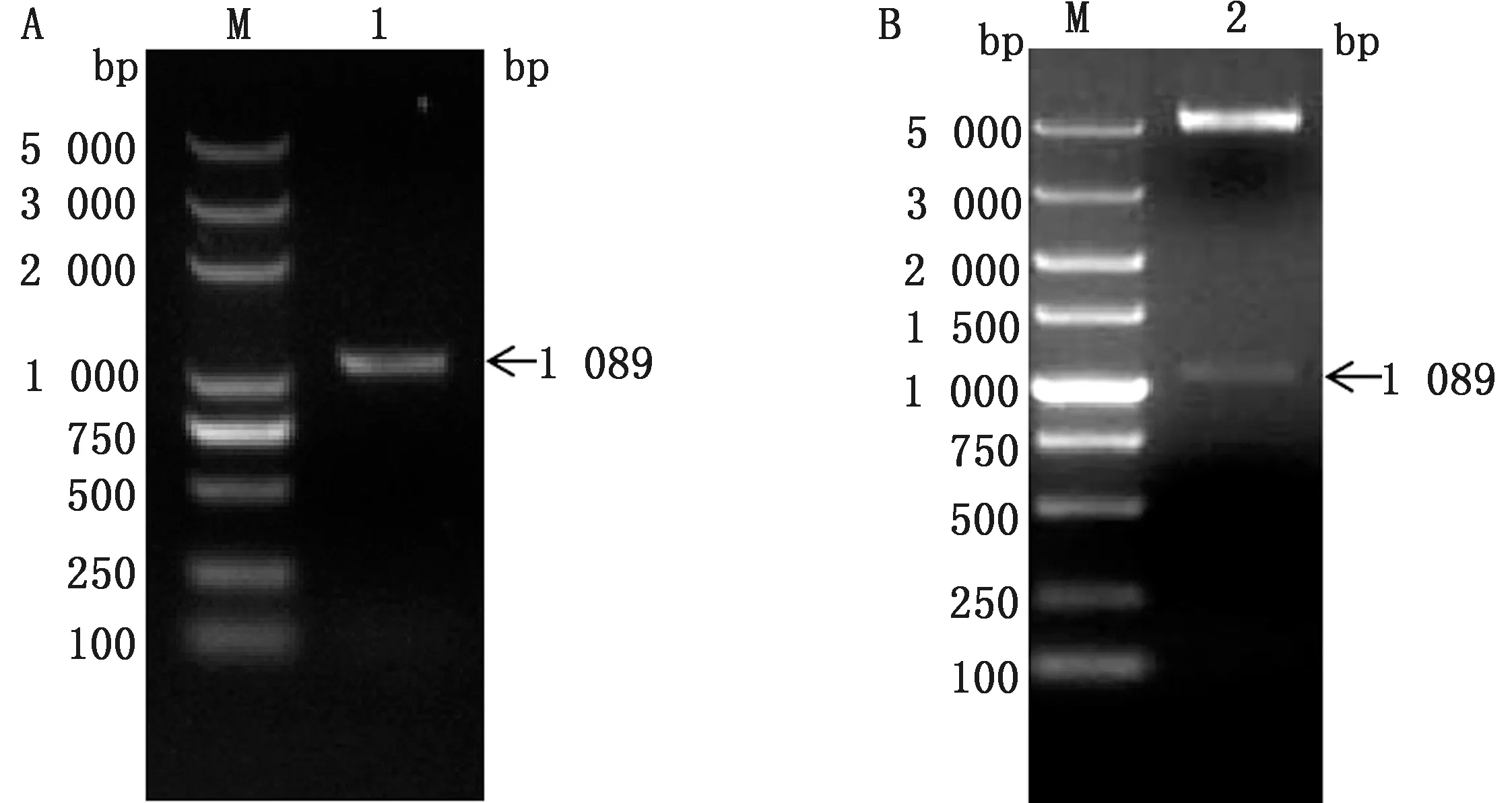
M.DL5000 DNA Marker;1.H359L基因 PCR 产物;2.NdeⅠ和EcoRⅠ双酶切pET-28a-H359L 质粒产物
2.2 pH359L重组蛋白的原核表达与纯化将阳性重组质粒转化至BL21感受态细胞后,加入终浓度为0.1 mmol的IPTG诱导表达6 h,收集菌体超声裂解进行可溶性分析。SDS-PAGE结果表明,IPTG诱导表达的pH359L重组蛋白以包涵体形式表达(图2A),经Ni柱纯化后的pH359L蛋白大小为43.0 kDa,获得与预期大小一致、单一的蛋白条带(图2B),表明得到纯度较好的pH359L蛋白。
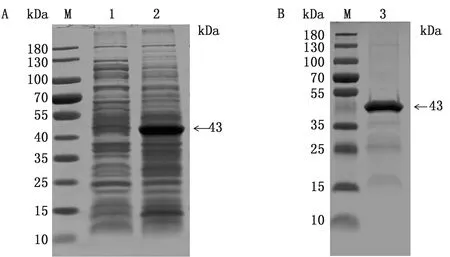
M.蛋白 Marker;1.pH359L诱导表达上清;2.pH359L诱导表达沉淀;3.pH359L纯化蛋白
2.3 重组蛋白反应原性鉴定将纯化后的重组蛋白SDS-PAGE电泳后转印至PVDF膜进行蛋白反应原性分析。Western blot检测结果表明,pH359L蛋白可以与ASFV阳性血清发生特异性反应,与阴性血清不发生反应(图3)。

M.蛋白 Marker;1.阳性血清;2.阴性血清
2.4 多克隆抗体的制备及抗体效价测定将纯化的pH359L蛋白用弗氏佐剂乳化后免疫新西兰大白兔,三免后收集血清,应用间接ELISA方法进行抗体效价测定。结果表明,抗pH359L蛋白多克隆抗体的抗体效价达到1∶512 000(图4),产生了较好的免疫反应。
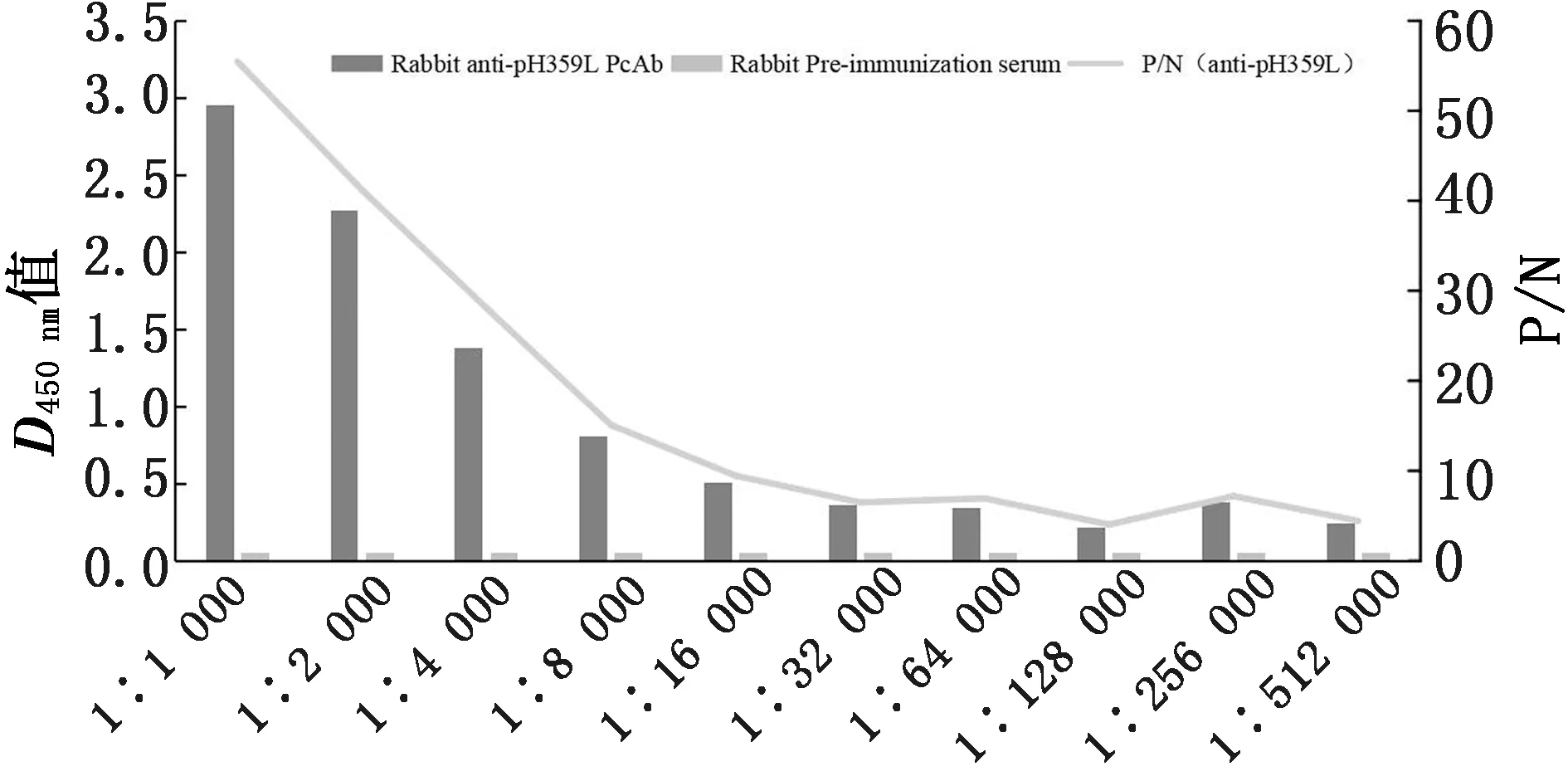
图4 兔抗pH359L多克隆抗体效价测定
2.5 多克隆抗体反应性鉴定利用ASFV感染PAM后制备的病毒抗原进行多克隆抗体的特异性鉴定。将本研究制备的pH359L多克隆抗体1∶200稀释后作为一抗进行Western blot分析,结果显示,抗pH359L蛋白的多克隆抗体能够与ASFV天然抗原发生特异性反应,表明制备的多克隆抗体具有良好的反应性(图5)。
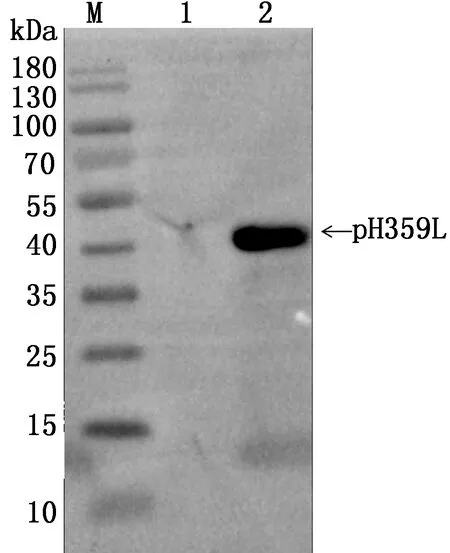
M.蛋白 Marker;1.ASFV未感染组;2.ASFV感染组
2.6 IFA验证ASFV毒株CN/GS/2018感染PAM 24 h后,用4%多聚甲醛固定,利用抗pH359L(1∶50)多克隆抗体进行免疫荧光分析。结果表明,抗pH359L多克隆抗体可识别ASFV感染PAM表达的天然蛋白pH359L(图6)。
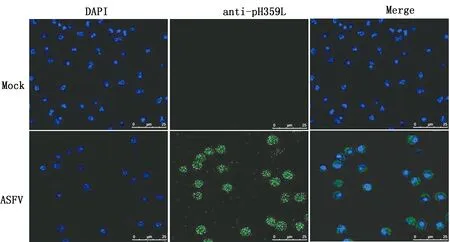
图6 兔抗pH359L蛋白多克隆抗体对天然ASFV H359L蛋白识别的检测
3 讨论
自1921年ASF首次报道后,主要流行于撒哈拉以南非洲地区。2007年格鲁吉亚暴发 ASF,随后疫情迅速蔓延至亚美尼亚、阿塞拜疆和俄罗斯。2014年ASF传入东欧大部分国家并逐步呈现出扩大流行的趋势[13]。2018年ASF于我国辽宁沈阳首次暴发[3],逐渐扩散到全国,对我国养猪产业造成巨大的经济损失和前所未有的打击。ASFV主要感染单核、巨噬细胞等免疫细胞,感染后导致严重的免疫损伤和免疫抑制[14]。目前尚无有效商品疫苗,急需阐明其感染、致病和免疫机制。但由于ASFV编码众多蛋白,且大多数蛋白功能未知,严重阻碍了对其致病机制研究的步伐。
ASFV与痘病毒类似,属于核质大DNA病毒(NCLDV)[10]。在ASFV感染细胞的过程中,病毒DNA复制起始于细胞核,存在短暂的入核过程,随后部分病毒基因复制和病毒组装发生在细胞质中。然而病毒DNA复制核内阶段的作用及其机制并不清楚;另外,复制的中间体也在ASFV感染细胞的细胞核和胞质中合成[15]。以上结果表明细胞核可能提供了小规模的转录模式。20%的ASFV基因组用于编码与其mRNA的转录和修饰有关的20多种基因[10]。
ASFV通过调控宿主基因的转录,以满足病毒生存和增殖[16]。目前,已报道A238L、A224L、D250R等病毒基因可调控宿主基因转录。A238L基因是IκB的同源基因,是NF-κB通路的抑制因子,它的表达能阻止NF-κB通路的激活[17],进而阻止被感染细胞的炎性因子、Ⅰ型干扰素、肿瘤坏死因子及IL-8等多种细胞因子的转录[18]。A238L蛋白还能和PP3C(protein phosphatase 3C)的催化位点结合,影响PP3C的活性并抑制NFAT(nuclear factor of activated T-cells)转录因子的激活,进而调控多种细胞因子如IL-2、GM-CSF、IL-3和IFN-γ基因的表达[19]。A224L能降解Myc/Max/Mad通路中的Mad蛋白[20],Myc/Max/Mad通路对细胞的生长、分化和凋亡具有重要的调控意义。在该通路中,Myc与 Max蛋白形成二聚体并与特定的DNA序列结合从而影响特定基因的转录与表达,Mad与 Max结合后则会抑制Myc与Max形成二聚体。A224L可通过降解Mad蛋白从而激活细胞内与生长、翻译相关蛋白的转录[16]。D250R与宿主细胞的脱帽酶2 (decapping enzyme 2 Dcp2)具有共同的Nudix基序[21],研究表明D250R在体外能对mRNA进行脱帽,在ASFV感染过程中具有对宿主细胞和病毒mRNA降解的功能。
由于ASFV pH359L与真核生物中主要负责聚合酶Ⅱ催化的核心亚基RPB3比较相似,因此蛋白pH359L在病毒转录过程中可能发挥重要作用[10]。本研究构建原核表达质粒pET-28a-H359L,IPTG诱导后经可溶性分析表明pH359L蛋白以包涵体形式表达,镍柱亲和层析纯化出纯度较高蛋白。经Western blot验证,pH359L重组蛋白能够和ASF阳性血清发生反应,为ASFV血清学诊断方法提供了依据。将纯化的蛋白免疫新西兰大白兔制备抗pH359L蛋白的多克隆抗体,经间接ELISA方法测定本研究制备的多抗效价达到1∶512 000以上,Western blot和IFA检测结果表明制备的抗pH359L多克隆抗体能够特异性识别ASFV感染PAMs后编码的pH359L蛋白,表明多克隆抗体具有很好的反应性和特异性。本试验纯化的pH359L蛋白及其免疫兔制备多克隆抗体,为实验室后续研究蛋白的功能及ASFV转录相关机制奠定基础。
