重金属的生物体原位成像技术研究进展
2022-07-22裴诚诚聂亚光赵亚楠倪珅瑶
裴诚诚,聂亚光,赵亚楠,倪珅瑶,许 安,2*
(1.安徽大学 物质科学与信息技术研究院,安徽 合肥 230601;2.环境毒理与污染控制技术安徽省重点实验室,中国科学院合肥物质科学研究院强磁场科学中心,安徽 合肥 230031)
重金属是指密度大于4.5 g/cm3的金属,一般存在于自然环境中,环境中的重金属主要分布于大气、水、土壤,其来源可分为自然来源和人为来源。随着工业化、城市化进程的不断推进,重金属的污染程度和潜在生态风险正逐渐增大[1]。目前我国很多地区受到不同程度的重金属污染,其中大气中的重金属主要来源于工业生产和汽车尾气排放,而水体中的重金属主要来源于各类生产、生活污水排放[2]。重金属能够通过饮水、被污水灌溉的粮食和蔬菜等途径进入人体,从而危害人体健康[3]。
重金属对人体的危害具有隐蔽性,当少量重金属摄入时不易被察觉,但通过食物链在人体中积累到一定程度会造成慢性中毒,损害人体健康[4],因此检测生物体内重金属的含量及分布对于解释其致毒机制十分重要。不同重金属对人体的毒性不同,例如微量的镉(Cd)通过生物累积进入人体后会对肾、肺、肝、睾丸、脑、骨骼以及血液系统产生一系列损伤[5];饮水和食物导致的长期砷(As)暴露会显著提升罹患癌症风险[6];汞(Hg)主要通过消化道、呼吸道和皮肤等侵入人体,造成肺、肾、肝、神经系统等发生病变[7];而铅(Pb)会降低人体的免疫力,直接损害生殖系统[8]。目前针对重金属的毒理学一般借助模式生物,如秀丽隐杆线虫[9]、蚯蚓、小鼠等进行研究,将重金属在生物体内的积累与不同的生物学终点相结合进行分析,如Yu 等[10]发现铜离子(Cu2+)在小鼠肝脏积累后会导致肝脏的线粒体吞噬及细胞凋亡从而造成肝损伤;Yin 等[11]的研究表明铬离子(Cr6+)在鱼的肝、肾中积累后,会引起氧化应激从而造成器官损伤。
重金属在生物体内的分布及形态是决定其生物效应的核心要素,是引起毒性和损伤的物质基础,因此原位检测生物体内重金属是进一步探索其致毒机制的重要手段。现阶段的主要检测技术包括激光烧蚀电感耦合等离子体质谱法(LA-ICP-MS)、同步辐射以及金属感测荧光团成像等。由于发展周期较短,因此原位检测重金属分布的方法有限,同时不同检测方法的原理、测试精度、对样品大小及前处理方法的需求不同[12],需研究人员对这些技术手段有全面深入的了解,以选择对自身研究最有利的方法。基于此,本文将从技术原理、前处理方法和实际应用等方面介绍上述原位检测生物体内重金属的手段。
1 激光烧蚀电感耦合等离子体质谱法(LA-ICP-MS)
1.1 LA-ICP-MS的基本原理及特点
LA-ICP-MS具有很高的灵敏度,是一种痕量和超痕量分析技术,具有较低的检出限,几乎无污染且干扰相对较小,可进行生物体内重金属的原位成像分析。LA-ICP-MS主要由激光剥蚀(LA)系统和电感耦合等离子体质谱(ICP-MS)系统两部分组成,基本原理是将高能激光束聚焦于固体样品表面进行剥蚀取样,随后使用ICP-MS 分析产生的气溶胶[13]。前者是一种快速直接的微区取样技术,后者用于检测所取生物样品中的元素含量。由于高能激光束是直径为数微米的光斑,因此通过LA连续采样覆盖生物样品时,每一次采样都如同一个“像素点”,测定其中的元素含量能获取整个生物体的元素分布,具体工作流程及原理如图1 所示[14]。由于样品的激光剥蚀过程需在真空中进行,因此在分析生物体内元素分布前需对样品进行预处理,如冷冻干燥、切片等使其样品状态和大小以达到仪器检测的标准。

图1 利用LA-ICP-MS检测秀丽线虫中银(Ag)含量的原理及工作流程[14]Fig.1 Schematic and workflow for the detection of silver(Ag)in C. elegans by LA-ICP-MS[14]
激光的工作条件(包括烧蚀模式、光斑大小和激光能量密度等)对检出限、分辨率和分析时间均有一定影响,测试前需确定合适的操作条件以保证分析的准确性。对于LA-ICP-MS,选择合适的空间分辨率十分重要,这不仅是一个光学设计问题,同时也取决于许多其他因素,如元素浓度、探测功率等。Brinkhaus等[15]总结LA-ICP-MS准确分析需满足3个条件:①激光剥蚀取样需具有代表性,即激光剥蚀产生的气溶胶的组成与样品组成相同;②具有高的气溶胶传输效率,即传输过程中气溶胶损失较少;③具有高的离子化效率,即气溶胶颗粒能在等离子体中完全离子化。
1.2 LA-ICP-MS在重金属原位成像中的应用
受限于样品仓尺寸和LA 扫描速度,LA-ICP-MS 一般用于检测小型动物,如线虫等[15-17]。Brinkhaus等[15]通过LA-ICP-MS研究了L1期线虫对锰(Mn)的吸收,将经过暴露的线虫离心冲洗冷冻干燥后直接进行检测,结果展示了线虫体Mn元素的分布。Wang等[14]利用L3期线虫检测了纳米二氧化钛(TiO2NPs)对Cd吸收的影响,结果显示TiO2NPs有助于Cd在线虫生殖细胞和卵中的积累(图2),从而增强了Cd的多世代毒性。Crone等[18]将L4期线虫暴露在含铂(Pt)的溶液中,洗涤后用冰甲醇进行固定,利用LA-ICP-MS检出Pt元素主要位于肠道及线虫头部。该技术可运用到其他动物体内金属元素含量的检测,对无法达到LA击穿厚度的大型动物,可通过器官或组织切片进行分析。Becker等[19]通过将小鼠心脏在液氮中冷冻后进行切片检测,得到的结果很直观地展示了小鼠心脏金属元素的分布,并观察到心肌中的重金属富集程度高于血液,与左心室相比,右心室的锌(Zn)、Mn、Cu 浓度更高。Togao等[20]用冷冻干燥将小鼠整个肾脏、肝脏和大脑样本处理后进行切片,通过图像结果对比发现Pb在肝脏中分布均匀,在肾脏和大脑中分布不均匀,其中在大脑中Pb 主要积聚于髓质而非皮质。Pornwilard等[21]报道LA-ICP-MS 分析证实了Wilson 病伴随有小鼠肝部铁(Fe)、Cu 和Zn 的异常值,具有作为常规诊断依据的潜力,为实验和临床肝病学提供了新的诊断证据。Egger 等[22]通过对人体组织切片的检测,发现除了肾脏、骨骼和肝脏中有积累外,Cd在肌肉和脂肪中的含量很低,在直肠及肾脏中分布不均匀,Zn在人脑中分布不均匀,推测其在大脑中不同位置的作用不同。

图2 利用LA-ICP-MS分析线虫体内Cd和Ti的分布[17]Fig.2 Distributions of Cd and Ti in C. elegans revealed by LA-ICP-MS[17]
除了动物个体和组织器官外,LA-ICP-MS 同样广泛应用于植物体内重金属的检测,如Becker等[23]在不同尺寸的薄片组织切片上观察到多种重金属元素的分布。杨红霞等[24]通过对暴露于Cd溶液的印度芥菜进行原位分析,发现Cd 大量积聚于韧皮部与木质部组成的维管组织,且Cd 与钙(Ca)具有相似的分布规律,说明重金属元素进入植株体内吸收运输过程与其它元素相伴。Promchan等[25]通过LAICP-MS 对单个大米纵截面进行重金属分布分析,以此区分大米样品的类型,从而为稻米颗粒分类提供了新依据。
LA-ICP-MS对许多重金属元素均具有高灵敏度和高准确性,可同时检测生物样品中多种重金属元素的分布,同样还可以通过外部校准进行半定量分析,以快速、直接进行生物原位分析(表1)。LA是LAICP-MS技术的核心,未来的分析应在成像分辨率、扫描速度以及对样品的穿透性等方面进行优化。

表1 LA-ICP-MS在生物成像中的应用Table 1 Application of LA-ICP-MS in bio-imaging
2 基于同步辐射(Synchrotron radiation)的成像方法
2.1 同步辐射成像的基本原理及特点
同步辐射光源是一种具有从远红外到X射线波长范围的连续光谱,具有高强度、高稳定性以及宽波段等特点,同步辐射技术是利用同步辐射光源轰击样品,激发样品中各元素的特征X射线,或与样品发生交互作用时产生各种信号,然后接收和分析相应信号,从而获得被轰击样品的化学组成、原子价态及结构组成等信息的技术[27],可用于原位分析生物样品。基于同步辐射技术的分析手段有很多,如同步辐射X射线荧光光谱(SRXRF)、同步辐射X射线吸收光谱(SRXAS)、X射线荧光显微镜(XFM)等[28],这些测试方法具有选择性强、灵敏度高等共同特性。根据不同的实验目的选择合适的同步辐射方法,无需复杂前处理过程即可检测生物样品中的重金属元素,实现原位无损分析。同步辐射技术的工作原理见图3[29]。

图3 识别晶体结构的微X射线吸收近边缘光谱(µ-XANES)及微衍射(µ-XRD)技术原理图[29]Fig.3 Schematic diagram of micro-X-ray absorption near-edge spectroscopy(µ-XANES)and micro-diffraction(µ-XRD)techniques for identifying crystal structures[29]
2.2 同步辐射方法在重金属原位成像中的应用
由于同步辐射光源具有极强亮度,高垂直性,所以SRXRF具有检出限低,对样品损伤小,微米级光束(可进行微区扫描、成像、结构等分析,提供微观精细信息)分辨率高,以及可同时检测样品中多种元素等特点。SRXRF的使用范围广,可运用到大部分生物体内的元素分析。Zhang等[30]将大肠杆菌在含镧(La)溶液中进行培养,再将线虫接到含La元素的琼脂板(NGM)中,利用SRXRF原位检出La在线虫L4时期体内的分布,推断线虫体内的La元素是通过食物大肠杆菌被摄入和积累。Gao等[31]通过SRXRF测得线虫体内K和Cu在体内的含量,发现暴露于Cu纳米颗粒可导致K和Cu含量升高且线虫中的Cu分布发生变化。袁静等[32]对Pb污染的土壤中蚯蚓进行原位分析,通过SRXRF发现蚯蚓体内富集一定量的Pb,且主要集中在蚯蚓前部,此外还发现Pb在蚯蚓消化道表面固定,但未产生毒性效应,因此推断蚯蚓消化道表面可以阻止重金属运移到其他器官。Al-Ebraheem等[33]使用化学固定剂和冷冻干燥制备了人体表皮组织样本,通过光斑尺寸为10µm×10µm的SRXRF检出样品中Fe、Cu在人体中的含量分布。Zhao等[34]通过SRXRF检出Hg和硒(Se)在大蒜根、叶和茎中的含量,Se使茎中的Hg积累增加,但根和叶中的Hg积累减少,且Se抑制了Hg从根到叶的转运。Meng等[35]分析显示稻米中的无机汞(Hg2+)在加工过程中会被清除,而毒性更大的甲基汞(MeHg)则被保留。同样可用于元素原位检测的还有XFM,Gao等[31]选用光束尺寸3µm×5µm的XFM观察Cu纳米颗粒在线虫中的生物累积,在成虫期,暴露于纳米颗粒后Cu和K的水平升高,Cu的生物分布与未暴露对照组相比,其在头部聚集明显。James等[36]通过XFM观察到线虫从幼年到产卵后肠细胞中的Fe有所增加,但不足引起线虫的衰老,实际衰老的原因是铁蛋白功能丧失和细胞稳态的丧失,因此线虫衰老程度与线虫体内Fe的含量并无直接联系。
重金属对环境和人体健康的危害不仅受浓度影响,还取决于其形态,SRXAS可原位检出生物体内重金属及微量元素的形态,为不同形态的重金属元素的毒性效应研究提供帮助。Weekley等[37]用SRXAS检测了由食物进行亚硒酸盐暴露的小鼠体内Se的形态及分布状态,发现在肾脏和肝脏中存在Se—Se和Se—C结构,肾脏中还存在未在肝脏中发现的Se—S结构,从而为在未来的疾病预防和治疗中有效合理地使用硒化合物奠定了基础。Diacomanolis等[38]用SRXAS将小鼠组织暴露于不同浓度的Cd2+溶液中,发现Cd2+在肝脏和肾脏中的配位环境一致,大多数Cd2+会与组织中金属硫蛋白结合。Porcaro等[39]通过将拟南芥暴露于高浓度Cd2+后,检出Cd2+在根和叶毛状体的维管中积累,造成植物生长受阻。
由于不同同步辐射装置检测出的信息不同,联用技术在原位成像方面发挥了很大作用。Guimarães等[40]利用同步辐射X射线荧光微束分析(SR-µXRF)发现As主要在虾的头部和腹部节段有局部积累(图4B),同时用µ-XANES在虾的头部对As进行表征,并与LA-ICP-MS结果进行对比分析,确定砷化物和砷酸胆碱是可能存在的形态,证实大多数As 以基本无毒的砷酸胆碱进行代谢(图4A、B)。Weekley等[41]用XAS和XFM联用技术检测了Se和Cu在小鼠肾脏中的分布及形态,发现在肾脏部位中这两种元素含量较高,而XAS分析显示在肾组织中存在Se—Se键,而非Se—Cu键,Cu在肾脏中与S、N元素相结合。Homma-Takeda等[42]通过SR-µXRF成像结合µ-XAFS,对小鼠组织进行化学染色,以1µm×1µm的空间分辨率分析获得铀(U)在小鼠肾脏中的分布。Kopittke等[43]将豇豆的根暴露在As(Ⅲ)和As(Ⅴ)溶液中24 h,通过SRXRF与µ-XRF联用检测根中As的含量,发现As(Ⅴ)的积累量更多。

图4 虾体内不同砷形态的荧光µ-XANES光谱(A),及虾体内的As、Ca、Br 元素分布(SR-µXRF)(B)[40]Fig.4 µ-XANES spectra of K-edge fluorescence of arsenic species in shrimp(A),and element distribution of As,Ca,Br in shrimp by SR-µXRF(B)[40]
基于同步加速器技术可对含水和新鲜植物组织中的金属和类金属进行原位检测,且不会造成明显的损伤[44]。同步辐射相比于LA-ICP-MS 的分辨率更高,且可检出重金属在生物体内的分布及形态,从而解释其不同形态重金属所造成的毒性效应,但该技术受设备机时和样品大小的限制。
表2列出了部分同步辐射在生物成像中的应用。

表2 同步辐射在生物成像中的应用Table 2 Application of synchrotron radiation in bio-imaging
3 金属感测荧光团(Metal sensing fluorophore)成像
3.1 金属感测荧光团成像的基本原理及特点
金属感测荧光团成像是通过荧光染料将金属离子染色,从而检测生物体内重金属的一种技术。图5 展示了荧光PET(光诱电子转移)传感器识别重金属的成像原理[45]。金属荧光传感器主要具有两个基本特征:①金属结合部位至少有一个荧光团,与待测金属结合后,用荧光显微镜检测被荧光染色的离子;②不同荧光染料对金属离子有特异性,因而能检出生物体内多种重金属的分布[46]。
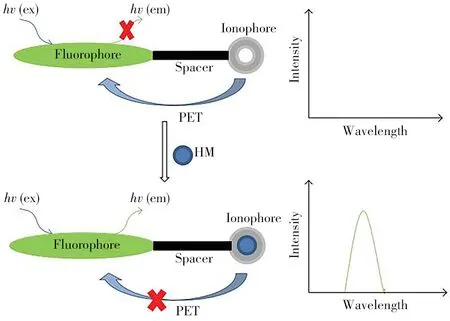
图5 荧光PET传感器识别重金属的原理[45]Fig.5 Schematic of heavy metal recognition by fluorescent PET sensors[45]
3.2 金属感测荧光团成像在重金属原位成像中的应用
金属感测荧光团技术在线虫中应用广泛,Huang等[47]通过一种基于超灵敏双链DNA 特异性染料和无标记核苷酸传感器,将8种金属离子染色后,发现对Pb2+具有很高的选择性。Huo等[48]开发了一种Pb2+的荧光探针(TPI),其通过催化脱丙炔化反应生成肟,能在80 s内快速检测到细胞内低浓度的Pd2+。荧光探针也可原位检测线虫体内的重金属元素,如Kaletta等[49]在线虫培养基上添加Zn特异性荧光染料,用荧光显微镜进行分析发现在线虫体内的Zn2+主要集中在肠细胞的囊泡中。Chung等[50]利用氰化物传感器与Cu2+之间的荧光作用判断是否存在Cu2+。有研究报道了一种Fe3+选择性荧光传感器,该传感器对Fe3+有62倍的荧光增强,可用于线虫体内Fe3+的分布检测[51]。Chen 等[52]用一种含巯基和羧酸基团的罗丹明腙衍生物作为Hg2+的选择性荧光传感器,可观察到Hg2+在线虫体内的积累,由于荧光检测的灵敏度高,可进一步探索Hg2+在体内的运移情况,这是首次用荧光传感器观察到生物体内nmol级别的Hg2+。
荧光感测技术不仅适用于线虫体内金属的检测(表3),同样也可用于其他生物体内金属离子的检测。Lakshmi等[53]发现含罗丹明单元的3种新型聚芳醚树枝状衍生物在含有其他竞争金属离子的情况下可选择性地感应小鼠体内的Hg2+。Hirayama 等[54]通过Cu 的荧光传感器揭示了小鼠体内Cu 含量的变化,为研究健康和疾病状态下Cu 的生理行为提供了新的技术手段。Chereddy 等[55]合成了4 种基于罗丹明的选择性识别金属离子的化学传感器,其中D 是1 种添加了三唑的无色化学传感器,分别考察未处理和Cu2+孵育的成纤维细胞的显微镜和荧光显微镜照片(图6A、B、D、E),发现在存在其他竞争性金属离子的情况下,D仍可与Cu2+形成粉红色的复合物(图6C、F)。Shu等[56]用小鼠的Cu2+-EDTA单克隆抗体捕获EDTA中的Cu2+,通过UV降解Cu2+-EDTA螯合物从而释放游离Cu2+,并利用CdSe/ZnS量子点荧光的猝灭效应检测Cu2+。Xu等[57]利用荧光染料EPNP 检测出芥菜植株和细胞中Hg2+的分布和运输,Hg2+被荧光染料染色后的共聚焦显微镜图像显示Hg2+主要积聚在溶解液泡而非细胞核或线粒体,研究结果为开展植物中金属离子的分布、转运研究提供了新的技术手段。

图6 未经处理的成纤维细胞(A),与Cu2+(5µmol/L)孵育的成纤维细胞(B),以及与Cu2+(5µmol/L)和D化学传感器(1µmol/L)孵育的成纤维细胞(C)的显微图像;未经处理的成纤维细胞(D),与Cu2+(5µmol/L)孵育的成纤维细胞(E),以及与Cu2+(5µmol/L)和D化学传感器(1µmol/L)孵育的成纤维细胞(F)的荧光显微图像[55]Fig.6 Microscopic images of untreated fibroblast cells(A),cells incubated with Cu2+(5µmol/L)(B),and cells incubated with Cu2+(5µmol/L)and D(1µmol/L)(C);fluorescence microscopic images of untreated fibroblast cells(D),cells incubated with Cu2+(5µmol/L)(E),and cells incubated with Cu2+(5µmol/L)and D(1µmol/L)(F)[55]

表3 生物感测荧光团成像的应用Table 3 Application of fluorescence sensor in bio-imaging
荧光团成像的核心机制是通过特异性荧光将特定的金属元素染色后进行直接观察。由于荧光染料的特异性,该方法在只针对生物体内某一种金属元素时具有很好的选择性,但和LA-ICP-MS及同步辐射技术相比,若同时将几种金属染色,则可能造成一定的干扰,因此并不适于同时研究生物体内多种重金属元素的含量及分布。
4 总结与展望
原位检测技术除以上介绍的3种方法,激光诱导击穿光谱(LIBS)[58-59]、拉曼光谱(需要制作基底间接检测[60])等技术在原位检测生物体内重金属方面的应用也在逐渐增加。根据已有文献显示,重金属在生物体内的原位成像已成为毒理学、病理学、环境化学等多学科领域的重要研究手段,是揭示重金属与生物体相互作用的重要工具。通常可根据研究需要选择合适的检测方法,LA-ICP-MS和同步辐射可以检测生物体内的几乎所有重金属元素[61-63]。其中LA-ICP-MS可检测的重金属种类与ICP-MS相同。而同步辐射技术一般用于检测生物体内元素价态及元素结合位点,如As形态的检测,Cu在生物体内结合方式的检测等[64-65]。金属感测荧光团成像已用于Ag、Cd、Zn、Hg、Pb、Cu等元素的检测[66-69]。尽管庞大的应用需求推动了这些检测手段的快速发展,也产生了许多具有典型代表性的工作,但受限于其自身的局限性,这些技术仍存在不足,如LA-ICP-MS的分辨率有待提高,样品大小和厚度受限,同步辐射技术的设备机时有限,难以开展大规模检测,金属感测荧光团在多种重金属同时染色时存在干扰等(表4)。通过对上述3种方法的总结,可发现样品大小、检测时间、空间分辨率是生物体原位检测时无法同时满足的需求:目前进行分析的样品主要以小型动物的整体或是大型动植物的组织切片为主,类似线虫、蚤等小型动物虽能做到整体的重金属分布检测,但分辨率不足以区分其内部组织器官,大型动物仅以切片样品进行检测,很难对重金属在生物整体层面的传输和富集进行表征,同时无限制扩大样品大小也会导致检测分辨率无法适应,从而造成检测时间无法控制,因此在技术条件存在较大限制的情况下,研究者更需在选择检测方法时谨慎权衡。此外,目前的检测方法还无法实现活体生物的检测,一些重金属与生物相互作用的细节无法被揭示,时间尺度上的动态变化过程只能通过截取不同暴露阶段的不同生物样品进行探索,无损且能够对测试生物进行实时监测的分析方法将是相关技术未来发展的方向。

表4 不同技术的优缺点及使用范围Table 4 Advantages,disadvantages and application scope of different technologies
