栓皮栎种子萌发出苗特征与生理生化变化*
2022-07-20王佳茜李国雷
王 娜 王佳茜 李国雷 李 箐 朱 琳 李 田 刘 文
(1.北京林业大学落叶栎研究中心 北京 100083; 2.北京林业大学省部共建森林培育与保护教育部重点实验室 北京 100083;3.北京市西山试验林场 北京 100041; 4.内蒙古自治区林业和草原保护总站 呼和浩特 010020)
我国栎属(Quercus)树种数量位居世界第三,面积和蓄积量均列我国树种首位,具有极其重要的经济和生态价值。根据第九次全国森林资源清查结果显示,被列入A类珍贵树种的前10种中有6种为栎属树种; 栎属树种同时也是我国重要的储备林树种。栎属树种属于极难生根树种,扦插繁殖困难,而目前栎类树种苗木存量较少,能用于嫁接繁育的砧木数量有限。大田播种和容器播种是栎属树种最为常见的育苗方式(Salifuetal., 2008; Olietetal., 2009, 2011; Lietal., 2014; Liuetal., 2016)。自然条件下,栎属植物种子早出苗和晚出苗间的时间差异达数周(Peterson, 1983; Bonneretal., 1987; 李庆梅等, 2013),通过错峰出苗时间减少幼苗在早春被食草类动物取食风险(Yietal., 2010),这是栎属树种重要的生存策略。然而,苗木培育要求种子快速有序整齐地出苗和生长(Lietal., 2012),如果出苗持续时间过长,先生长出来的叶片将覆盖相邻幼苗的生长,阻挡光照和水分的吸收(Giertychetal., 2011; 刘艳等, 2012),将会导致苗木规格大小不一、质量参差不齐。因此,明确栎类种子出苗异质性对于调控其苗木质量具有重要意义。
栓皮栎(Q.variabilis)是栎属落叶阔叶乔木树种(吴明作, 1998),耐干旱、瘠薄能力强,广泛分布于我国华中、华北、华南、西北、西南地区(吴征镒, 1980),在植被恢复和重建中发挥着重要作用。激素和酶是种子重要组分并发挥着关键生理作用,目前,对栓皮栎的研究主要集中在种子更新与苗木培育(Lietal., 2014; Puerta-Pieroetal., 2006; Salvadoretal., 2005; Trubatetal., 2010)、生物学特性、育苗造林等方面(吴明作, 1998; 周建云等, 2010),但关于栎类种子自身萌发特性、出苗不齐的内含物变化研究及其相关关系的研究较少。本研究分析栓皮栎种子萌发和出苗特征,并分析延迟萌发和出苗不齐的机制,为制定栓皮栎出苗整齐性措施以及精准调控苗木质量提供理论依据和科学指导。
1 材料与方法
1.1 试验材料
2019年8月27日—2019年9月30日在北京市平谷区丫髻山林场,每隔3天采集1次栓皮栎种子。采集半同胞家系种子,以减少种子之间的遗传差异。所采集种子浸种去除瘪粒、虫蛀种子和杂质,再用(50±3)℃的温水浸泡种子30 min去除虫卵,室内平铺于桌面自然风干10~16 h。栓皮栎种子质量差异较大,为减少种子初始物质对萌发出苗的影响,对鲜种子逐一称重,选取4.0~4.5 g的种子装袋贮藏于1~5 ℃的种子贮藏箱中低温贮藏3个月,贮藏期间进行翻动。
1.2 试验方法
1.2.1 种子处理与播种 筛选500粒未萌发且无虫孔的大小相似、颜色相近的健康栓皮栎种子,播种前用清水浸泡24 h,期间对栓皮栎种子翻动,使其充分浸泡、吸水、膨胀,取出后再用清水进行冲洗,当年12月份播种。育苗基质选用丹麦品氏Seeding型草炭(pH6.0,粒径0~6 mm)与国产蛭石以3∶1的比例均匀混合,育苗容器选用DeepotTMD60(上口径6.4 cm,深度36 cm,体积983 mL; Stuewe & Sons, Inc., Corvallis, America)。播种前对种子和育苗基质消毒处理,用多菌灵消毒后,先将栓皮栎种子播入托盘中,每天翻动一遍,筛选出萌发的栓皮栎种子,将种实播种于D60容器,覆土厚度1 cm,充分浇水,标记萌发日期,直至连续10天无萌发种子为止。播种后,置于温室(平均温度23 ℃,平均湿度68%)每隔2天用喷雾器喷水,保持基质湿润直至出苗。期间对栓皮栎种子出苗情况进行动态监测,标记出苗日期,并拍照记录。
1.2.2 种子萌发出苗统计 当第1株栓皮栎种子出苗时(播种后第21天)破坏取样,每3天进行1次,共破坏取样6次,即取样时间t1-t6分别为播种后第21、24、27、30、33和36天。每次破坏取样,分别选取该时期未萌发的栓皮栎种子、刚萌发(胚根突破种皮)的栓皮栎种子和刚出苗(幼苗露出地表面2~3 cm),5个重复,每个重复3粒种子,共计210粒(播种后30天栓皮栎种子萌发结束,因此在t5和t6取样时只有出苗状态的栓皮栎种子),用液氮冷冻5 min后取出,置于-80 ℃超低温冰箱中保存。
1.2.3 出苗量计算 当首粒栓皮栎种子出苗后,每天记录其萌发出苗数量,连续10天未有种子出苗,终止统计。
栎类种子萌发出苗动态通常呈反曲线型,根据Hay等(2014)提出的probit分析方法进行数据转换,使出苗率与出苗时间建立正态分布模型:
g=Φ(γ)=Φ(β0-β1p-1)。
式中: g为种子出苗率占播种种子的比例;γ为线性预测因子,即出苗概率;β0为最大出苗概率;β1为萌发率Probits统计量随天数的倒数减小速率;p为出苗天数。也可看做种子出苗速率参数(ER)。ER50%=(β0-γ)/β1, 此处γ=0, 因此ER50%=β0/β1可以根据公式计算中位萌发速率(ER50%),即栓皮栎种子出苗速率,ER50%越大,出苗速度越快。根据出苗统计量计算栓皮栎种子出苗速率,可对栓皮栎种子整个出苗时期进行划分(史文辉, 2018)。
1.2.4 酶活性测定 SOD活性测定采用氮蓝四唑(NBT)还原法(张志良,2009),POD活性测定用愈创木酚法(李合生,2000)。
1.2.5 内源激素测定 采用高效液相色谱仪测定GA3、IAA、ABA激素含量(Lietal., 2011)。
1.2.6 营养物质及其分解相关酶活性测定 可溶性糖和淀粉含量测定采用蒽酮比色法(李合生,2000)。α-淀粉酶和β-淀粉酶活性测定采用3,5-二硝基水杨酸显色法(李合生,2000)。
1.3 数据分析
采用SPSS 19.0(IBM 19.0 for Windows, SPSS Statistic )软件进行试验数据的分析处理,利用Sigmaplot 12.5软件(Systat Software International, USA)作图。
2 结果与分析
2.1 栓皮栎种子萌发出苗特征
2.1.1 种子出苗规律 在整个出苗过程中,栓皮栎种子出苗率中位数GR50%为0.025(图1)。由图2可知,依据日均3%出苗率标准将出苗过程划分为上升期(t1、t2)、高峰期(t3、t4)和下降期(t5、t6)。播种后1~24天为出苗上升期,占整个出苗周期的40%,此期间出苗种子数占出苗总数的11%; 25~30天为出苗高峰期,占整个出苗周期的10%,此期间出苗种子数占出苗总数的44%; 31~60天为出苗速率下降期,占整个出苗周期的50%,此期间出苗种子数占出苗总数的45%。在播种后第21天开始出苗,在第30天左右,栓皮栎种子出苗数量达到峰值,栓皮栎种子每日绝对出苗率升至最高15.4%,随后逐渐下降,播种60天后栓皮栎种子出苗结束,累计出苗率为66.6%。栓皮栎种子自身存在延迟萌发和出苗不整齐的现象,最先出苗和最后出苗的种子之间相差可达40天。

图1 栓皮栎种子出苗统计量与播种后时间倒数的线性关系Fig. 1 Linear relationship between the emergence statistics of Q. variabilis seeds and the reciprocal time after sowing

图2 栓皮栎种子绝对出苗率和累计出苗率Fig. 2 Absolute emergence rate and cumulative emergence rate of Q. variabilis seeds
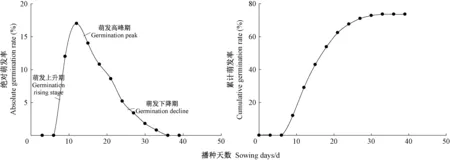
图3 栓皮栎种子绝对萌发率和累计萌发率Fig. 3 Absolute germination rate and cumulative germination rate of Q. variabilis seeds
2.1.2 种子萌发规律 由图3中可知,播种后第6天开始萌发; 在第9~12天时,萌发达到峰值,日均最大萌发率为17%; 之后,种子萌发数逐渐降低,在第33天萌发结束,累计萌发率为73.6%。播种后的9~14天为萌发高峰期,当日平均萌发率达5.5%,所占时间为整个萌发周期的18.2%,此期间萌发种子数占萌发总数的42.1%; 播种后1~8天为萌发上升期,占整个萌发周期的24.2%,此期间萌发种子数占萌发总数的16.3%; 播种15~33天为萌发下降期,占整个萌发周期的57.6%,此期间萌发种子数占萌发总数的41.6%。最早萌发的种子与最晚萌发的种子之间相差27天,萌发周期较长,这可能是导致后期栓皮栎种子出苗周期长,出苗不齐的原因之一。
2.1.3 种子萌发-出苗相关性分析 由图4可知,先萌发的种子不一定出苗时间早,栓皮栎种子萌发和出苗是2个相对独立的过程,栓皮栎种子胚根萌发和胚芽出苗均存在休眠现象。种子萌发出苗过程中,从播种到萌发天数的中位数为14天,从播种到出苗天数的中位数为29天,从萌发到出苗天数的中位数为16天; 种子从播种到萌发天数的众数为11天,从播种到出苗天数的众数为26天,从萌发到出苗天数的众数为15天(图5)。种子萌发虽有不同步,但是离群点较少,整体较为集中,而萌发到出苗阶段离群点与播种到出苗类似,说明栓皮栎种子出苗过程中出苗周期长,出苗不整齐主要由于萌发到出苗阶段不齐导致。
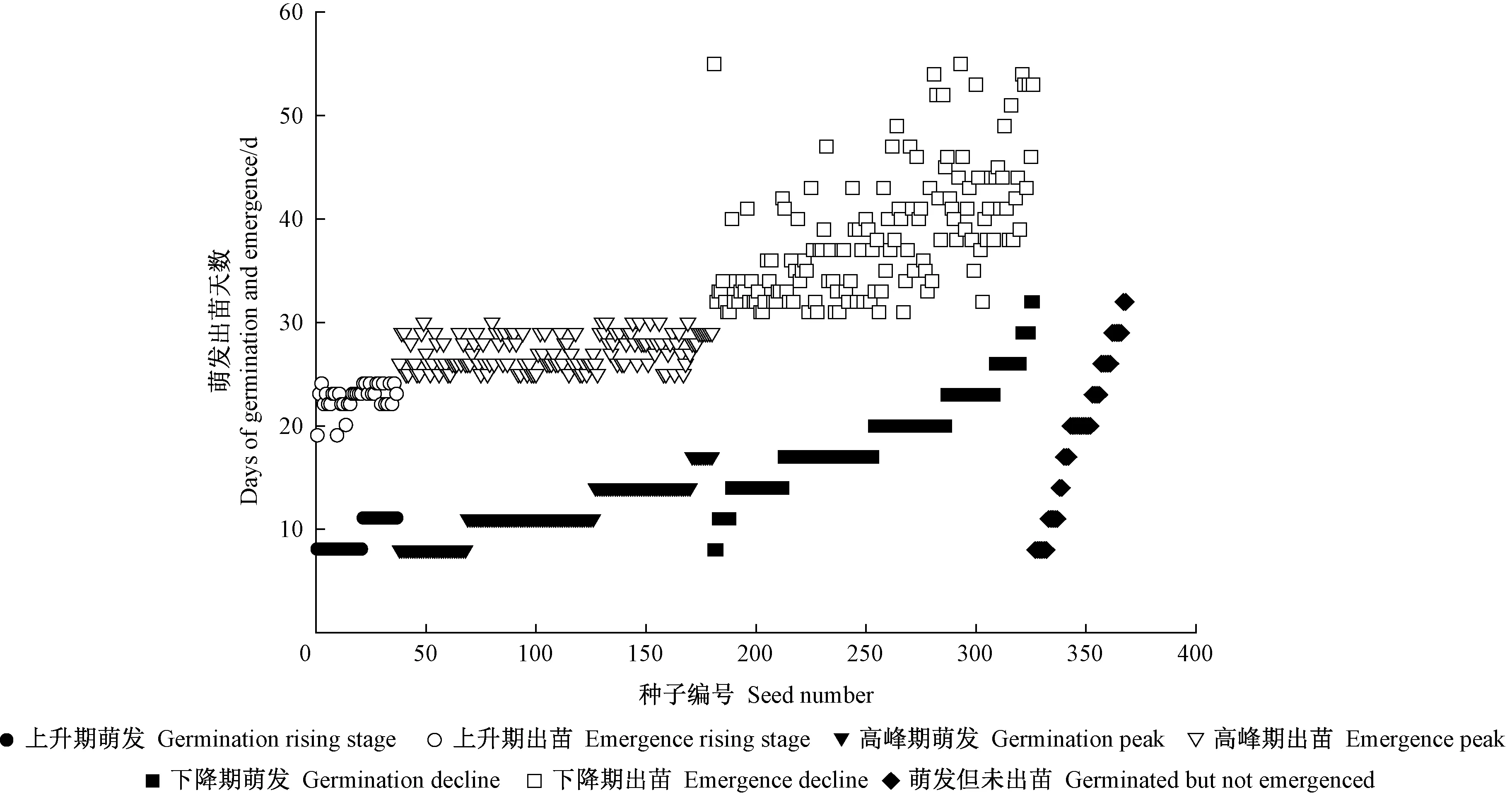
图4 栓皮栎种子萌发出苗关系Fig. 4 The relationship between the germination and emergence of Q. variabilis seeds

图5 栓皮栎种子萌发出苗情况箱线图Fig. 5 The boxplot of germination and emergence of Q. variabilis seeds
2.2 不同时间萌发出苗种子的酶活性变化
前期萌发的种子POD活性显著高于后期萌发的栓皮栎种子,而前期出苗的种子则POD活性较低。随着出苗时间的增长,种子出苗速率加快,POD活性也逐渐升高(图6)。栓皮栎种子在出苗高峰期(t3~t4时期)POD活性较高,显著高于其他时期。与POD活性变化不同,前期萌发的栓皮栎种子内SOD活性最低,随后活性逐渐升高,在播种后第27天(t3),SOD活性升至最高。与种子的萌发类似,出苗初期,SOD活性较低,随着出苗的速率增加(t1~t4),SOD活性也呈升高趋势,并于t4达到峰值。
2.3 不同出苗时间种子的内源激素变化
由图7可知:萌发早的种子GA3含量高,萌发晚的种子GA3含量较低; 早期出苗和萌发的种子,GA3的含量显著高于未萌发的种子,高峰期出苗的种子中GA3的含量显著高于萌发未出苗的种子、未萌发的种子。由此可见,GA3含量与栓皮栎种子萌发出苗速率相关。萌发出苗早的种子IAA含量较萌发晚的高,出苗早的种子IAA含量也高于出苗晚的种子。种子萌发出苗过程中,IAA含量变化与GA3含量变化相似,表明IAA与GA3之间可能存在协同作用,共同促进栓皮栎种子的萌发及出苗过程。在萌发高峰期,未萌发的种子中ABA含量显著高于萌发和出苗的种子。随着ABA含量的降低,栓皮栎种子出苗数量逐渐增多。
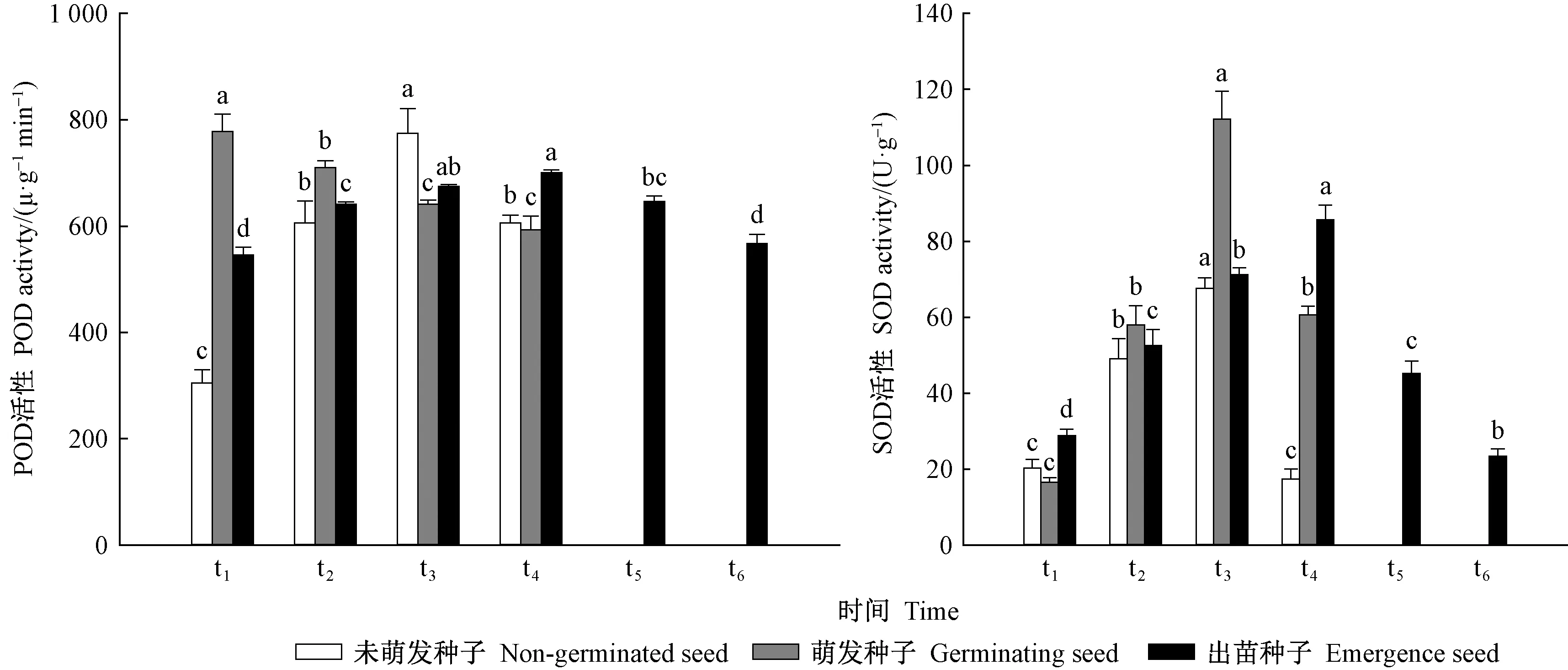
图6 不同时间萌发和出苗的种子POD和SOD酶活性变化Fig. 6 Changes of POD and SOD enzyme activities in seeds germinated and emerged at different timest1:播种第21天; t2:播种第24天; t3:播种第27天; t4:播种第30天 ;t5:播种第33天; t6:播种第36天。下同。t1: 21st day of sowing; t2: 24th day of sowing ;t3: 27th day of sowing ;t4: 30th day of sowing ;t5: 33rd day of sowing; t6: 36th day of sowing. The same below.字母不同表示差异显著(平均值±SE,α=0.05,n=5)。下同。Bars marked with different letters differ significantly in each variable (mean ± SE, α=0.05, n=5).The same below.
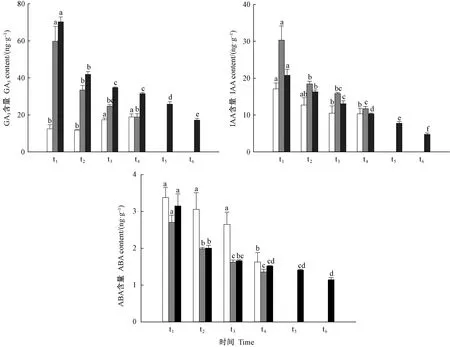
图7 不同时间萌发和出苗的种子内GA3、IAA和ABA含量变化Fig. 7 Changes of GA3, IAA and ABA content in seeds germinated and emerged at different times
2.4 不同时间萌发出苗种子内营养物质含量及其分解相关酶活性变化
由图8可知,萌发早的种子可溶性糖含量较萌发晚的种子低; 萌发种子的可溶性糖含量普遍低于未萌发的种子,在萌发阶段t1时期可溶性糖含量最大降幅为56.18%,以上结果表明种子萌发时期大量消耗自身营养物质进行供能。出苗早的种子可溶性糖含量较出苗晚的种子低,随着出苗时间延长,可溶性糖的分解代谢显著减弱。前期萌发的种子淀粉含量较后期萌发的种子高; 前期及后期出苗的种子的淀粉含量较出苗高峰期低。以上结果表明前期出苗的种子,主要通过可溶性糖进行供能; 随着出苗时间的延长,可溶性糖逐渐被分解消耗,淀粉发挥的作用逐渐显著。
由图9可知,萌发早的种子α-淀粉酶活性较萌发晚的种子低。由于种子萌发阶段主要通过分解可溶性糖进行供能,淀粉分解量少,萌发早的种子β-淀粉酶活性较萌发晚的种子低。种子萌发过程中需要淀粉进行分解供能,种子内β-淀粉酶活性增加,可加快淀粉的水解。出苗时间早的种子β-淀粉酶活性较出苗时间晚的种子低,表明随着出苗时间的延长,β-淀粉酶活性升高,加速种子内淀粉水解,为种子出苗提供能量。

图8 不同时间萌发和出苗的种子内可溶性糖和淀粉含量变化Fig. 8 Changes of soluble sugar and starch content in seeds germinated and emerged at different times
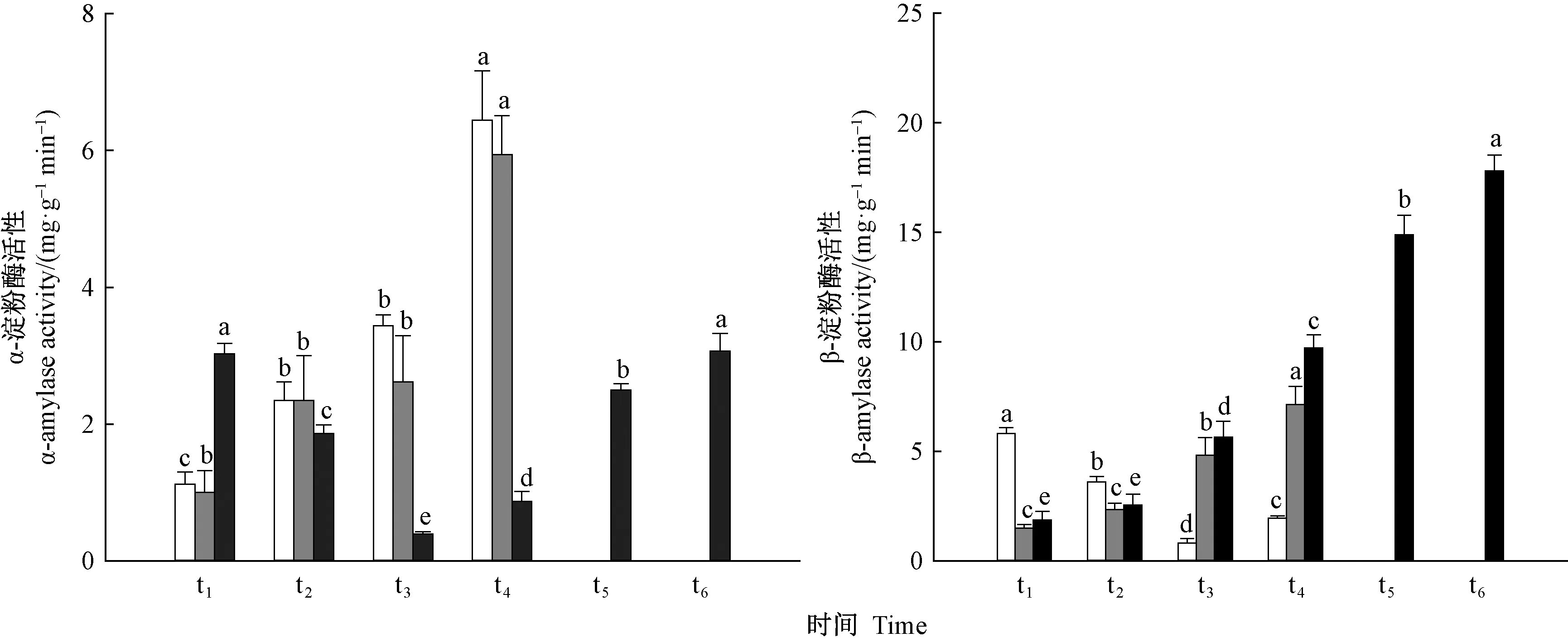
图9 不同时间萌发和出苗的种子内α-淀粉酶和β-淀粉酶活性变化Fig. 9 Change of α- amylase and β- amylase activityin seeds germinated and emerged at different times
3 讨论
3.1 栓皮栎种子萌发出苗特征
栓皮栎种子存在休眠现象,但种子休眠程度不一,致使休眠浅的种子较早即可萌发,休眠程度深的种子萌发时间晚。栓皮栎种子自身种皮较厚较硬,也可能是由于种皮的机械束缚,导致种子萌发周期较长。而出苗过程则受温度、水分、光照以及种子内部营养物质、酶活性以及激素含量多种因素影响共同控制。部分栓皮栎种子可以萌发但最终不一定能出苗。Peterson(1983)对水橡(Quercusnigra)树的研究表明,种皮组织中存在化学抑制剂,这可能是导致栎树种子发芽延迟和出苗不整齐的原因。红栎(Q.ruber)树种子通常延迟发芽,通过分层或其他方法克服休眠后,栎树种子体内代谢活性增加(Kaul, 1985)。去除栎树种子的基部种皮可以促进种子萌发(Association, 1985),去除部分子叶也可以达到同样的效果,且少量子叶的去除并不影响栎幼苗的后期生长(Houetal., 2010)。但无论去除种皮还是去除部分子叶,种子出苗周期长、不同步现象未得到改善,这会导致容器育苗后出苗的栓皮栎种子由于光照和水分的摄取不足,苗木质量下降,苗木生长缓慢。本研究表明,栓皮栎出苗不齐可分为2个阶段进行研究,即播种-萌发阶段和萌发-出苗阶段,而萌发-出苗阶段不整齐是导致栓皮栎出苗不齐的主要原因。
3.2 活性氧与种子萌发出苗的关系
SOD和POD等抗氧化酶是种子活性氧系统的重要构成部分,能有效预防种子代谢过程中产生的活性氧物质在种子中的积累(Kosteretal., 2000)。POD在植物中普遍存在,其活性对植物种子萌发及生长具有重要意义(王艳华, 2005),POD活性在种子出苗高峰期达到最大值,说明较高的POD活性可以促进栓皮栎种子出苗过程。POD活性与SOD活性呈极显著正相关,POD参与调节分解代谢IAA的过程,能氧化分解IAA使其失去活性,进一步影响GA含量(李昭玲等, 2015),以维持植物的正常生长,调节植物体的生长。Roberts(1973)认为POD增强了PPP途径在种子发芽过程中的主导作用从而促进种子发芽。SOD首先将植物体内的自由基氧化成H2O2,然后POD将其还原成水(浦梅, 2016)。SOD活性在种子出苗高峰期达到最大值,说明较高的SOD活性可以促进栓皮栎种子出苗过程。因此,POD、SOD活性较高时,有利于清除种子内部累积的有毒物质,保持了种子的高活力(汪晓峰等, 2001),有利于栓皮栎种子的出苗过程。根据本研究的结果推测,栓皮栎种子的萌发与出苗可能需要一定阈值的SOD活性,且这一阈值在不同种子个体之间存在差异。早萌发出苗的种子可能需要较低的SOD活性即可,而大部分种子需要较高的SOD活性才能萌发出苗,这也导致了栓皮栎种子出苗不齐的现象。
3.3 内源激素与种子萌发出苗的关系
种子休眠与植物激素的相互作用相关。对IAA、ABA和GA的成熟和分层水栎胚胎进行了定量和定性分析表明,随着栎属植物种子的发育,IAA和ABA含量增加,而随着栎属植物种子成熟度的增加,IAA和ABA含量降低(Blancheetal., 1980)。Hopper(1982)研究认为种皮和栎树种子休眠之间存在着显著的相关性。本研究栓皮栎种子萌发过程中,种子内GA3、ABA和IAA含量都呈下降趋势,种子内GA3、IAA含量较高时,种子萌发较快,仅ABA含量不足以控制栓皮栎种子的休眠和萌发; 出苗晚的种子中,由于GA3、IAA含量低,种子的出苗时间也随之增加。ABA抑制栓皮栎种子萌发和出苗过程可能也依赖于较低水平的GA3和IAA。随着种子持续吸胀,ABA含量降低,对种子萌发和出苗的抑制作用减弱,使得种子达到出苗高峰期。未萌发的栓皮栎种子内GA3和IAA含量都显著低于萌发的栓皮栎种子,而未萌发的栓皮栎种子内ABA含量高于已经萌发的种子,因此,在栓皮栎种子萌发出苗过程中,同样存在植物激素的协同交叉调控,栓皮栎种子的萌发出苗受到多种激素的精细调控。
3.4 营养物质含量及其分解相关酶活性与种子萌发出苗的关系
种子可溶性糖是糖代谢的中间产物,主要由种子中淀粉的不断降解形成。种子中可溶性糖的含量影响淀粉在种子中的降解速率和耗速率(刘雅帅,2008)。糖的消耗以及能量转化速率的差别可导致种子萌发和出苗不整齐。未成熟或处于萌发早期阶段的种子内可溶性糖含量较高(杨博,2010),本研究中未萌发的栓皮栎种子内可溶性糖和淀粉含量高于同期已经萌发和出苗的栓皮栎种子。活力低的种子,不能为胚生长提供能量,是种子不能萌发和出苗的原因之一。
本研究还发现,在萌发过程中,栓皮栎种子内α-淀粉酶和β-淀粉酶活性逐渐升高,分解淀粉为栓皮栎种子萌发提供物质和能量; 栓皮栎种子出苗过程中,出苗前期种子内α-淀粉酶活性降低,对淀粉利用率降低,β-淀粉酶活性逐渐升高,分解部分淀粉,保证种子的正常代谢活动,随着出苗时间的增加,出苗后期种子内α-淀粉酶和β-淀粉酶活性升高,对淀粉水解作用增加,为种子出苗提供物质和能量。
4 结论
栓皮栎种子萌发和出苗周期分别为4周和6周,萌发和出苗周期跨度大,栓皮栎出苗不整齐,对苗期管理带来极大困难。POD和SOD主要影响栓皮栎种子的萌发阶段; GA3和IAA促进栓皮栎种子出苗,含量越高出苗速度越快; ABA抑制栓皮栎种子的出苗。