转BtCry1Ac与API基因107杨对节肢动物群落多样性及稳定性的影响*
2022-07-20任敏霞张子恒曾月霞王利峰杨敏生刘军侠
任敏霞 李 探 张子恒 曾月霞 王利峰 杨敏生,4 刘军侠,4
(1.河北农业大学林学院 保定 071000; 2.河北省承德市兴隆县行政审批局 承德 067000;3.湖南省衡阳市林业科学研究所 衡阳 421200; 4.河北省林木种质资源与森林保护重点实验室 保定 071000)
长期采用常规农药防治农林害虫,会导致害虫产生抗药性、害虫再猖獗及杀虫剂污染环境等严重问题,转基因植物应用为防治虫害提供了一条新途径(周晓静等, 2018; 赵蓬晖等, 2010)。目前已成功将抗虫基因转入多种植物,且部分已进入田间释放与商业化种植阶段(杨双, 2018),但其对靶标、非靶标生物及生态环境的影响存在争论(姜韬, 2018; 夏敬源等, 1999; 武二忠, 2011; 凌闵, 2020),转基因植物在带来经济效益的同时,其生态安全性问题不容忽视。对其生态安全性研究主要集中于: 外源基因的稳定性、靶标害虫是否产生抗性、对非靶标生物的影响、对群落结构的影响、对土壤微生物的影响、是否会产生基因漂移与水平转移及其直接或间接的环境影响。有研究发现外源基因能稳定存在于植物中(李立, 2009),也有研究发现转基因杨树外源基因表达量产生变化,发生了基因沉默现象(吕威等, 2018)。目前研究表明转基因杨树对靶标害虫表现出较高抗性,个体数量显著减少(任亚超, 2013; 王璞等, 2020),且抗虫性可以持续到下一代,延缓下一代幼虫的发育(刘冬艳等, 2015)。转基因杨树(Populus)在抑制靶标害虫的同时,对非靶标生物的影响存在一定争论,多数研究结果表明未对非靶标害虫及天敌产生明显不利影响(胡建军等, 2010; 田亚坤, 2014),甚至对天敌具有一定增殖作用(姚丽等, 2016),但也有研究表明转基因杨树对靶标害虫的持续控制作用导致非靶标害虫叶蜂(李探等, 2018)及刺吸类害虫数量有所增加(陈辉惶等, 2012),寄生性天敌种类与数量相对减少(张芳, 2004; 胡建军等, 2007)。到目前为止的研究未发现对群落、土壤微生物及环境的明显不利影响(Wangetal., 2018; 胡建军等, 2010)。
杨树因其速生、用途广泛、基因组小、易离体操作等特点,被用作转基因林木研究的模式树种(睢韡等, 2019; 丁莉萍, 2016)。转基因林木生态安全性及对环境的影响,可能需较长时间才能显现出来,因此需对其进行长期监测。此外,采用常规方式及嫁接方式繁殖的转基因杨树抗虫性均较高; 嫁接繁殖的转基因杨树还可有效避免或降低外源基因发生漂移及转移的风险,提高其环境安全性(王智, 2014)。目前对嫁接繁殖转基因杨树的群落影响研究较少,本研究以嫁接转基因107杨(Populus×euramericana‘Neva’)为对象,探讨嫁接转基因杨树对节肢动物的群落组成、多样性及稳定性的影响,为转基因杨树生态安全性评价积累数据,并为其推广与应用提供科学依据。
1 材料与方法
1.1 试验材料
本试验林分为伐根嫁接的转基因107杨(BtCry1Ac和API基因),株系#1、#2、#3、#9均为经室内筛选,对鳞翅目害虫美国白蛾(Hyphantriacunea)和杨扇舟蛾(Closteraanachoreta)等具有高抗虫性,转入基因的毒蛋白表达水平为#1>#2>#9>#3(杨艳丽等, 2012)。于2014年在107杨伐根上嫁接转基因107杨各株系及对照(未转基因107杨为欧美杨107号,属于美洲黑杨×欧洲黑杨“无性系”,P.×euramericanacv.‘74/76’)1年生枝条,成林后用于试验调查。2015—2019年的林龄分别为2、3、4、5、6年。
1.2 试验地概况
试验地位于唐山市滦南县国营林场苗圃,滦南县属暖温带沿海半湿润区大陆性气候,年均气温11℃,年均降水量约660 mm,年均日照2 853 h,无霜期186 天。试验样地采用随机区组设计,划分为4个区组,每个区组种植转基因107杨株系#1、#2、#3、#9及对照107杨(CK)各30 株(5 列× 6 行),区组内株系间随机块状排列,株距为2 m,行距5 m,区块面积为300 m2,不同区组以黑杨(美洲黑杨×欧洲黑杨“无性系”)作为隔离带,隔离带宽度4 m,整块地地形地貌气候一致,除正常的水肥管理外,均未施用化学农药。
1.3 调查方法
调查于2015—2019年4—10月进行,每20~25天调查1次。采用随机取样方法,在每个区组中各株系分别随机选取5株样树,调查其上节肢动物种类与数量。树冠调查: 在选取的植株东、西、南、北4个方向,上下各选取50 cm样枝,观察并记录样枝上节肢动物种类与数量。主干调查: 调查样株距地表3 m内主干上节肢动物种类与数量。地面调查: 在选取的样地内按照五点式取样法,选取5个1 m×1 m的样方,记录节肢动物种类与数量。对于野外调查不能鉴定的种类,将其编号,装入酒精试管中,带回实验室进行鉴定(张巍巍等, 2011; 胡金林,1983)。
1.4 试验林基本生长情况
采用随机取样的方法,每个小区随机选取5株树,共100株,对试验林基本生长状况进行了调查。随年份的变化,树高、胸径及枝下高均表现为一定程度的增长,不同年份转基因107杨生长量产生波动,#2株系相对稳定,#3株系生长量相对较低。
1.5 数据分析方法
通过以下公式进行群落特征指数分析。
1)Shannon-Wiener多样性指数(diversity index):
式中:S为物种数;Pi为相对多度。
2)Pielou均匀度指数(evenness index):
J=H′/Hmax,Hmax=lnS。
式中:Hmax为给定物种完全均匀时的群落多样性指数;S为物种数。
3)Berger-Parker优势度指数(dominance index) :
C=Nmax/N。
式中:Nmax为优势类群的数量;N为群落中总的个体数。
4)通过以下指标比较群落中不同株系的稳定性,值越大说明在种间数量上的制约作用相对较大,天敌物种数相对较多,对植食性昆虫有较强的制约作用,表明群落相对稳定。Ss/Si: 总群落物种数与群落总个体数比值;Sn/Sp: 捕食性天敌与植食性昆虫物种数比值;Sa/Sb: 捕食性天敌与植食性昆虫个体数比值;Sp/Sb: 植食性昆虫物种数和个体数比值;Sn/Sa: 天敌物种数和个体数的比值。
1.6 数据处理
依据转基因107杨调查数据,逐年统计节肢动物组成以及节肢动物不同功能类群种类和数量,比较不同株系不同年度间转基因107杨节肢动物群落及各功能类群组成、群落特征指数及稳定性差异。
采用PAST2.17计算转基因107杨群落特征指数,利用SPSS 21.0中单因素(One-way ANOVA)方差分析对转基因107杨亚群落组成、群落特征指数进行差异显著性分析。双因素(Two-way ANOVA)方差分析对转基因107杨不同株系和不同年份亚群落组成及群落特征指数进行差异显著性分析。相关数据整理、绘图与制表在Microsoft Excel 2010中完成。
2 结果与分析
2.1 节肢动物群落基本组成
节肢动物群落基本组成见表1。共获得节肢动物54 091头,分属13个目。节肢动物群落物种数以鳞翅目(Lepidoptera)、鞘翅目(Coleoptera)、双翅目(Diptera)、膜翅目(Hymenoptera)与蜘蛛目(Araneida)占优势。个体数量以鳞翅目占优势,其次为半翅目(Hemiptera)、膜翅目、蜘蛛目、鞘翅目、双翅目,分别占5年数据总和的48.2%、15.0%、14.2%、8.5%、8.4%、4.1%。脉翅目(Neuroptera)、直翅目(Orthoptera)与其他类群个体数量相对较少,趋于稳定。
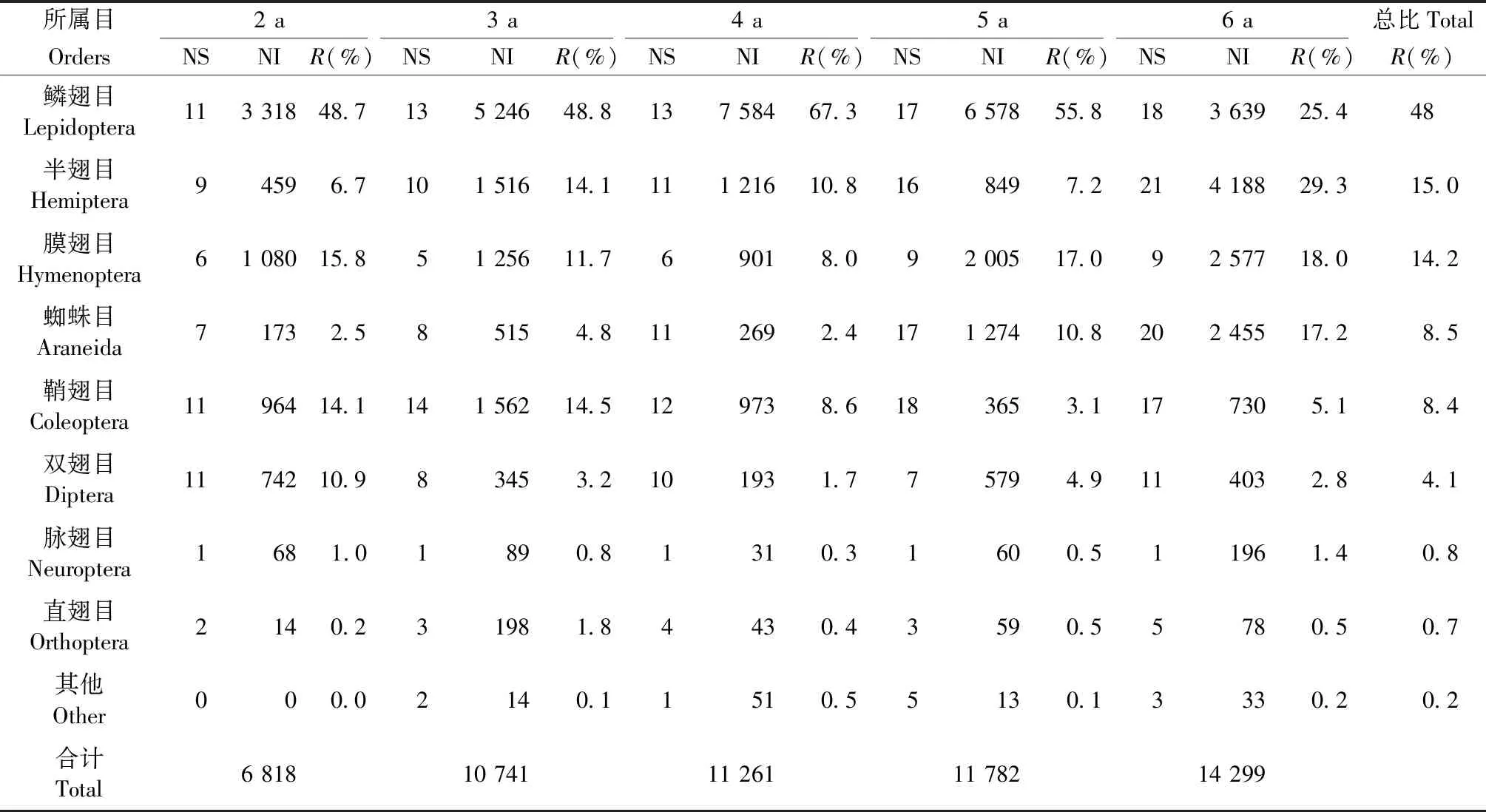
表1 节肢动物群落基本组成①Tab.1 The basic composition of arthropod community
2.2 转基因107杨亚群落组成
2.2.1 不同年份转基因107杨亚群落组成 分析转基因107杨亚群落组成,包括害虫、天敌和中性节肢动物物种数和个体数。
1)转基因107杨害虫亚群落组成 转基因107杨害虫亚群落物种及个体数如图1、2所示。由图1可见,在同一年份,转基因107杨与对照107杨的物种数无显著差异,随年份变化物种数表现为一定增长趋势。由图2可见,同一年份的转基因107杨各株系的害虫亚群落个体数量均显著低于对照107杨(P<0.05)。调查数据显示: 主要靶标害虫杨扇舟蛾、美国白蛾数量为对照107杨林分较多,各株系总体低于对照。个体数量随林龄变化有一定波动,各株系间杨扇舟蛾个体数量在林龄2 、4、5年相对较多,3、6年相对较少; 美国白蛾个体数量在林龄2、5年相对较多,林龄6 年明显减少。综合以上分析,转基因107杨对害虫亚群落的物种数影响不显著,但能有效控制靶标害虫的个体数量,表现出持续抗虫效果。
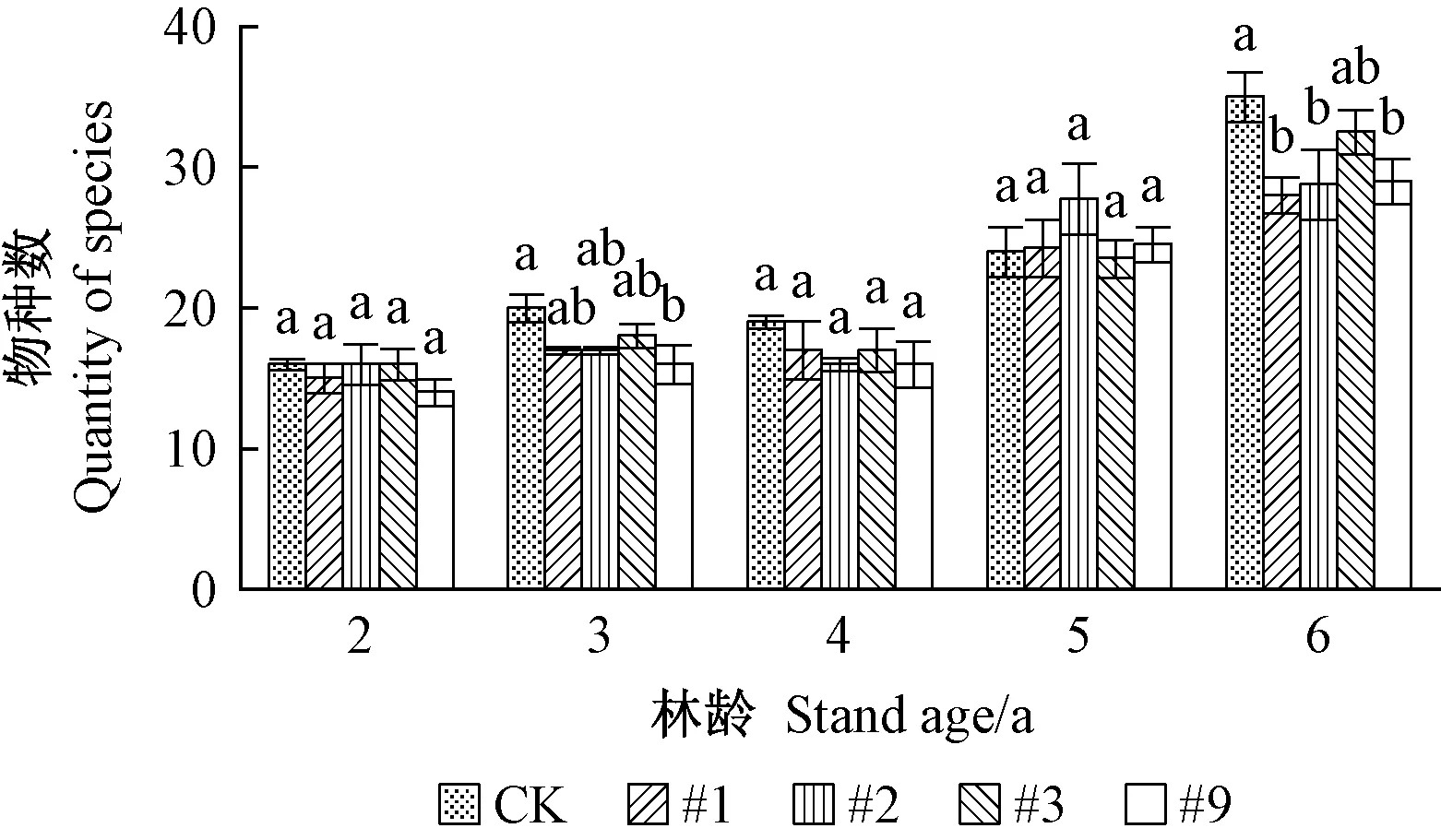
图1 转基因107杨害虫亚群落物种数Fig. 1 Quantity of species of pest sub-community in transgenic poplar 107同一林龄不同小写字母代表同一指标不同株系在0.05水平下差异显著,下同。Different lowercase letters in the same age mean that different strains of the same index have significant differences at the level of 0.05, the same below.
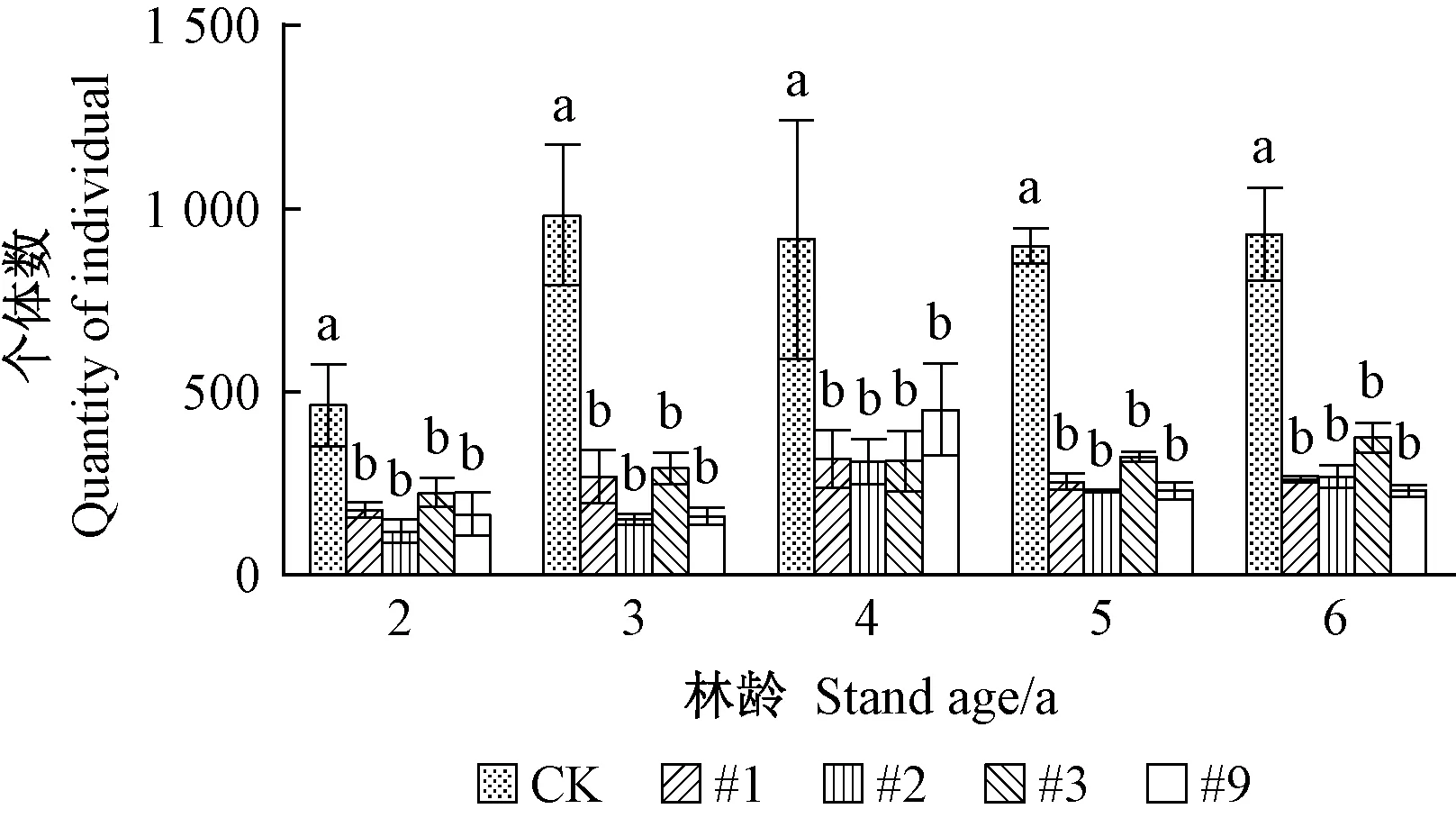
图2 转基因107杨害虫亚群落个体数Fig. 2 Quantity of individual of pest sub-communityin transgenic poplar 107
2)转基因107杨天敌亚群落组成 转基因107杨天敌亚群落物种及个体数见图3、4。由图3可见,同一年份转基因107杨天敌亚群落物种数与对照107杨无显著性差异(P>0.05),物种数在林龄2、4年相对稳定,5、6年增多。由图4可见,同一年份转基因107杨天敌亚群落个体数与对照107杨差异不显著(P>0.05),个体数量在林龄6年相对较多,与其余年份相比呈上升趋势,主要是由于瓢虫(Coccinellidae)类及蜘蛛类节肢动物类群个体数量增多如异色瓢虫(Harmoniaaxyridis)个体数量在林龄3、6年相对较多,2、4、5 年相对较少,狼蛛(Tarantula)个体数量随年份变化呈现明显增长趋势。上述结果表明转基因107杨在抑制靶标害虫的同时,天敌亚群落物种及个体数量没有减少,后期呈现一定增加趋势。
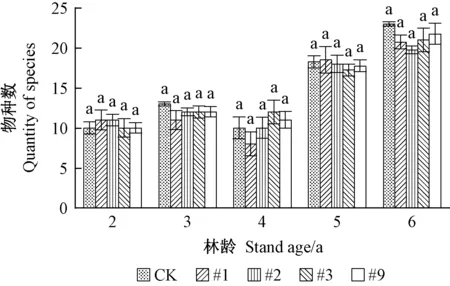
图3 转基因107杨天敌亚群落物种数Fig. 3 Quantity of species of natural enemy sub-community in transgenic poplar 107
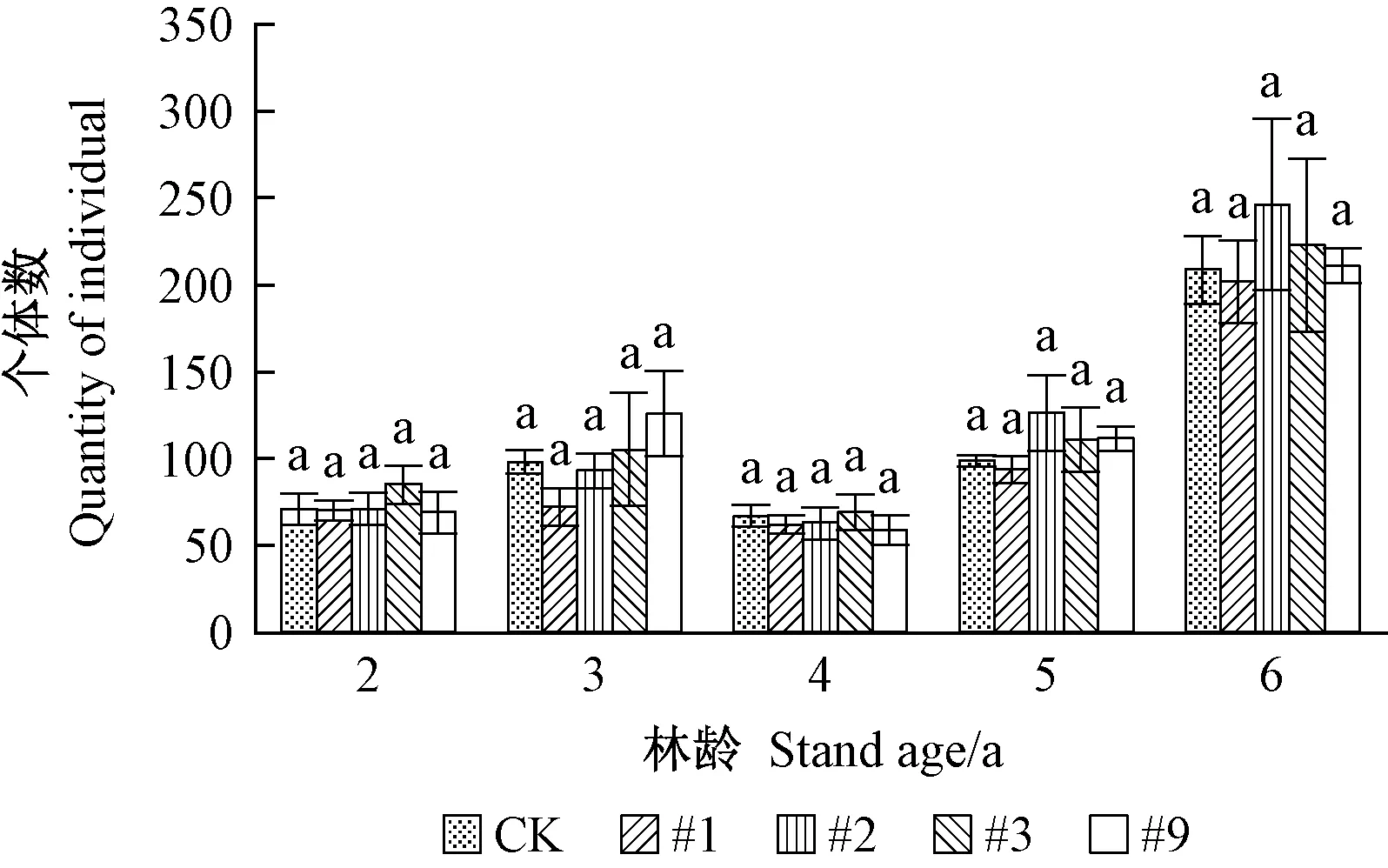
图4 转基因107杨天敌亚群落个体数Fig. 4 Quantity of individuals of natural enemysub-community in transgenic poplar 107
3)转基因107杨中性节肢动物亚群落组成 转基因107杨中性节肢动物亚群落物种及个体数见图5、6。由图5可见,同一年份转基因107杨与对照107杨物种数不存在差异显著性(P>0.05),随年份变化物种数略有下降。由图6可见,同一年份转基因107杨与对照107杨中性节肢动物个体数量无显著差异性(P>0.05)。随年份的变化个体数量产生波动,林龄5、6 年个体数量与其余各年份相比呈明显增长趋势,其他年份表现为林龄3年与2、4 年相比个体数量增多,2、4年波动较小变化不明显。例如家蚁(Monamoriumpharaonis)、家蝇(Muscadomestica)在林龄2、4年个体数量相对稳定,变化不大,林龄5、6年与2、4年相比个体数量呈增长趋势。综上所述,转基因107杨对中性节肢动物群落物种及个体数量无显著影响,随年份变化个体数量表现为一定程度的增加。
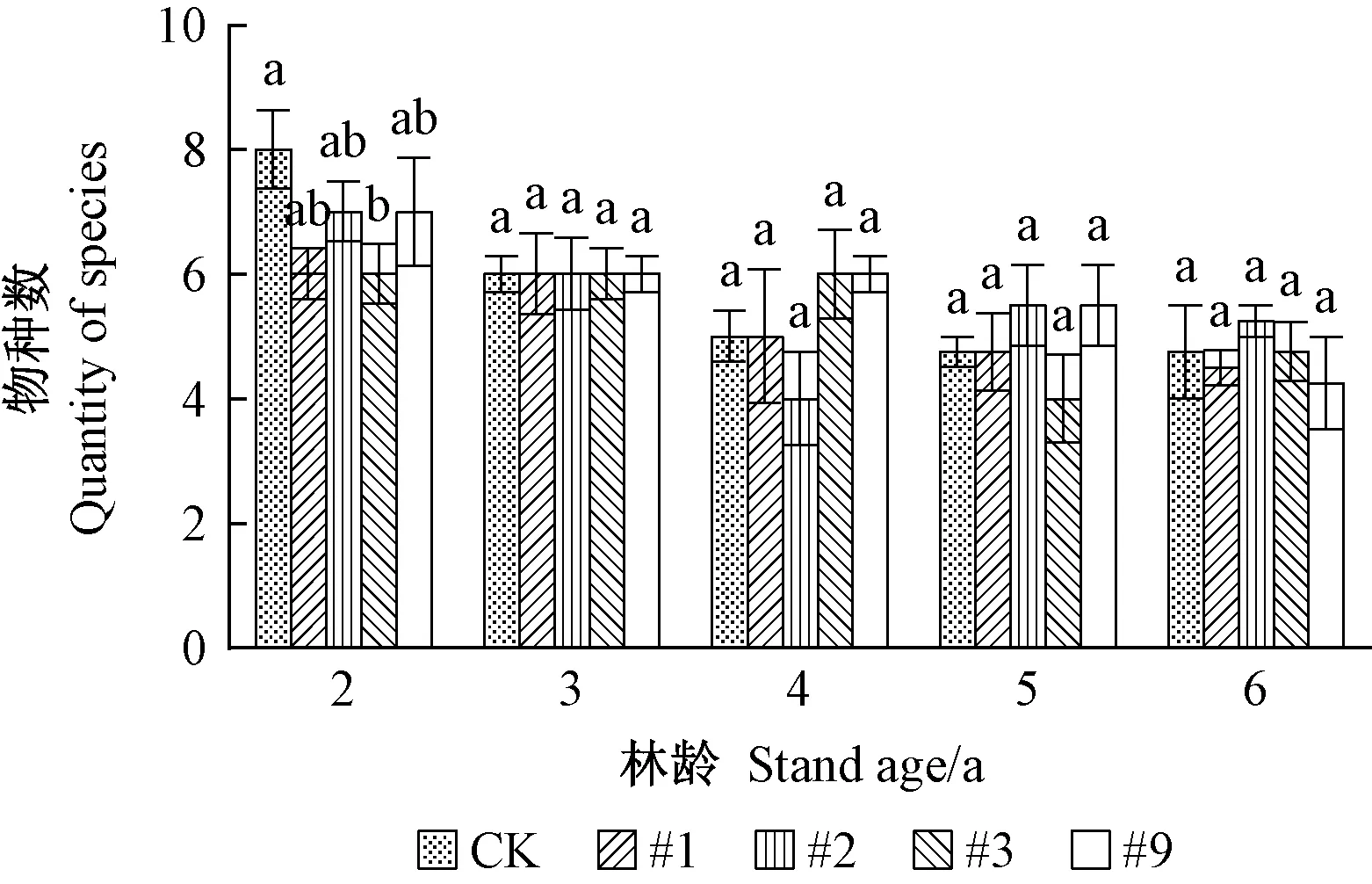
图5 转基因107杨中性节肢动物亚群落物种数Fig. 5 Quantity of species of neutral arthropod sub-community in transgenic poplar 107

图6 转基因107杨中性节肢动物亚群落个体数Fig. 6 Quantity of individuals of neutral arthropod sub-community in transgenic poplar 107
2.2.2 转基因107杨亚群落组成双因素方差分析转基因107杨亚群落组成双因素方差分析见表2。转基因107杨不同株系对害虫亚群落物种及个体数影响显著(P<0.05),对天敌与中性节肢动物亚群落物种、个体数量影响均不显著(P>0.05)。年份对各亚类群落物种、个体数影响均显著(P<0.05),株系与年份交互影响不显著(P>0.05)。上述分析表明,转基因107杨对害虫亚群落个体数量影响较大,对天敌及中性节肢动物亚群落无显著影响,年份间各亚类群落物种、个体数变化较大。
2.3 转基因107杨节肢动物群落特征指数
转基因107杨节肢动物群落特征指数见表3。同一年份转基因107杨节肢动物群落多样性、均匀度指数均显著高于对照107杨,优势度指数显著低于对照107杨(P<0.05),各株系间差异不显著(P>0.05)。整体来看,#2株系随年份变化波动较小,多样性、均匀度指数较高,优势度指数较低,群落结构较为稳定。转基因107杨节肢动物群落特征指数随年份的变化表现为林龄2、4年波动较小,5、6年多样性、均匀度指数上升,优势度指数降低。综合以上分析表明,转基因107杨对群落特征指数未产生显著不利影响,随年份变化表现出更为稳定的趋势。
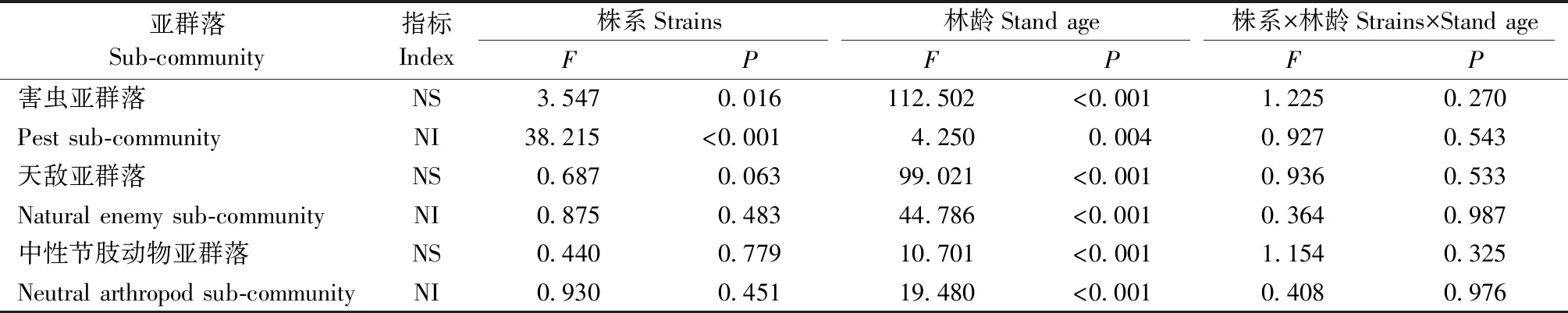
表2 转基因107杨亚群落组成双因素方差分析①Tab.2 Two-factor analysis of variance for sub-community composition in transgenic 107 poplar
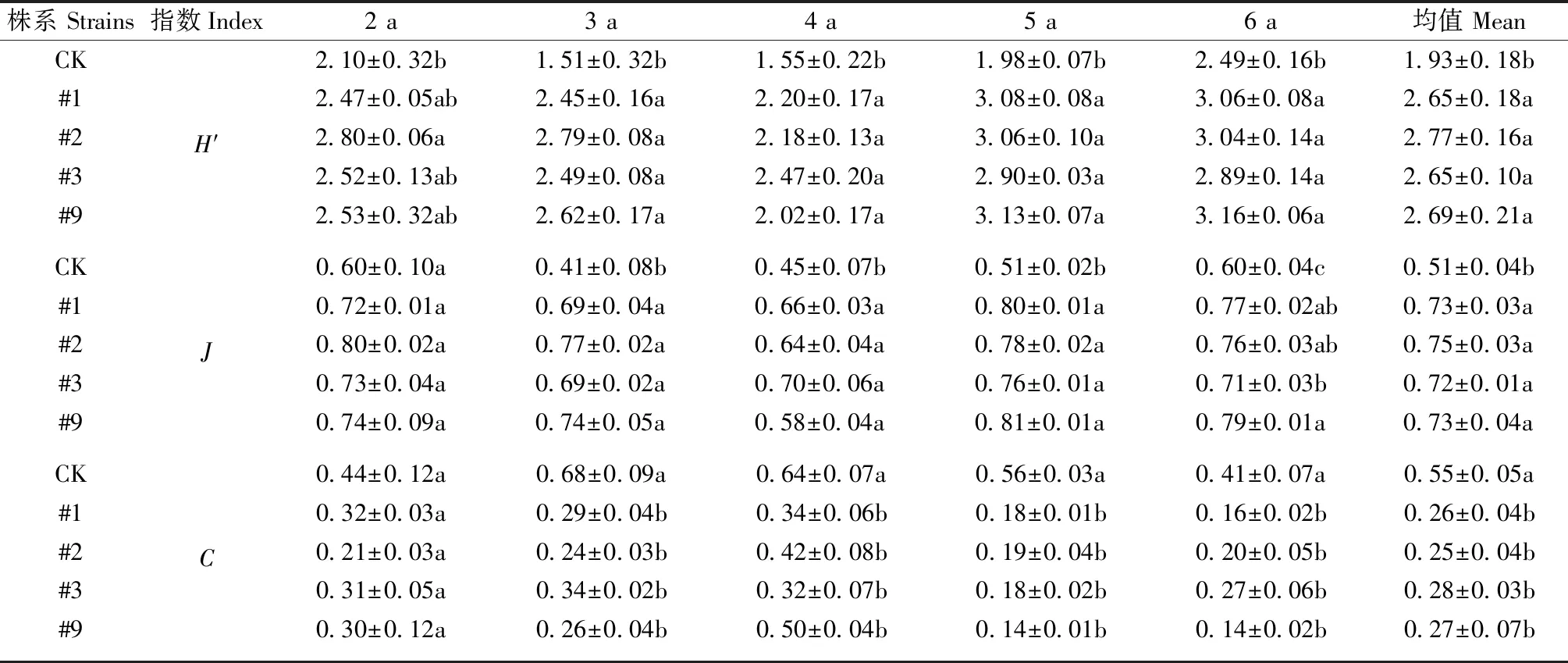
表3 转基因107杨节肢动物群落特征指数①Tab.3 Community characteristic index of transgenic 107 poplar arthropods
转基因107杨节肢动物群落特征指数双因素方差分析表见表4。可见,转基因107杨群落特征指数受不同株系及林龄影响显著,株系与林龄的交互作用对多样性指数影响显著(P<0.05),而对均匀度及优势度指数无显著影响(P>0.05)。
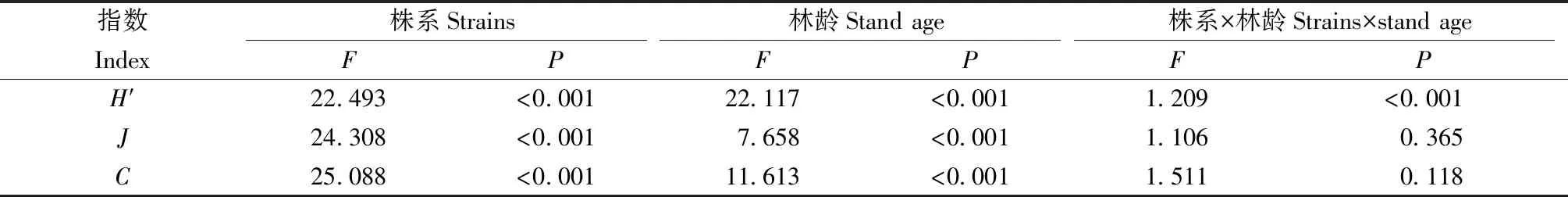
表4 转基因107杨节肢动物群落特征指数双因素方差分析表Tab.4 Two-factor analysis of variance of index in transgenic 107 poplar arthropod community characteristics
2.4 转基因107杨稳定性分析
转基因107杨节肢动物群落稳定性结果见表5。可知: 转基因107杨Ss/Si,Sp/Sb,Sa/Sb指标均显著高于对照107杨(P<0.05),表明转基因107杨物种数多,个体数量少,转基因107杨在种间数量上制约作用强于对照107杨,转基因107杨Sn/Sa,Sn/Sp指标与对照相比差异不显著(P>0.05),表明转基因107杨与对照107杨捕食性天敌物种数与个体数无明显差异。在林龄5、6 年,#3株系稳定性有所下降,与其他株系差异显著,其余年份各株系间差异不显著,#2株系随年份变化保持较高稳定性。综上所述,转基因107杨稳定性高于对照107杨。
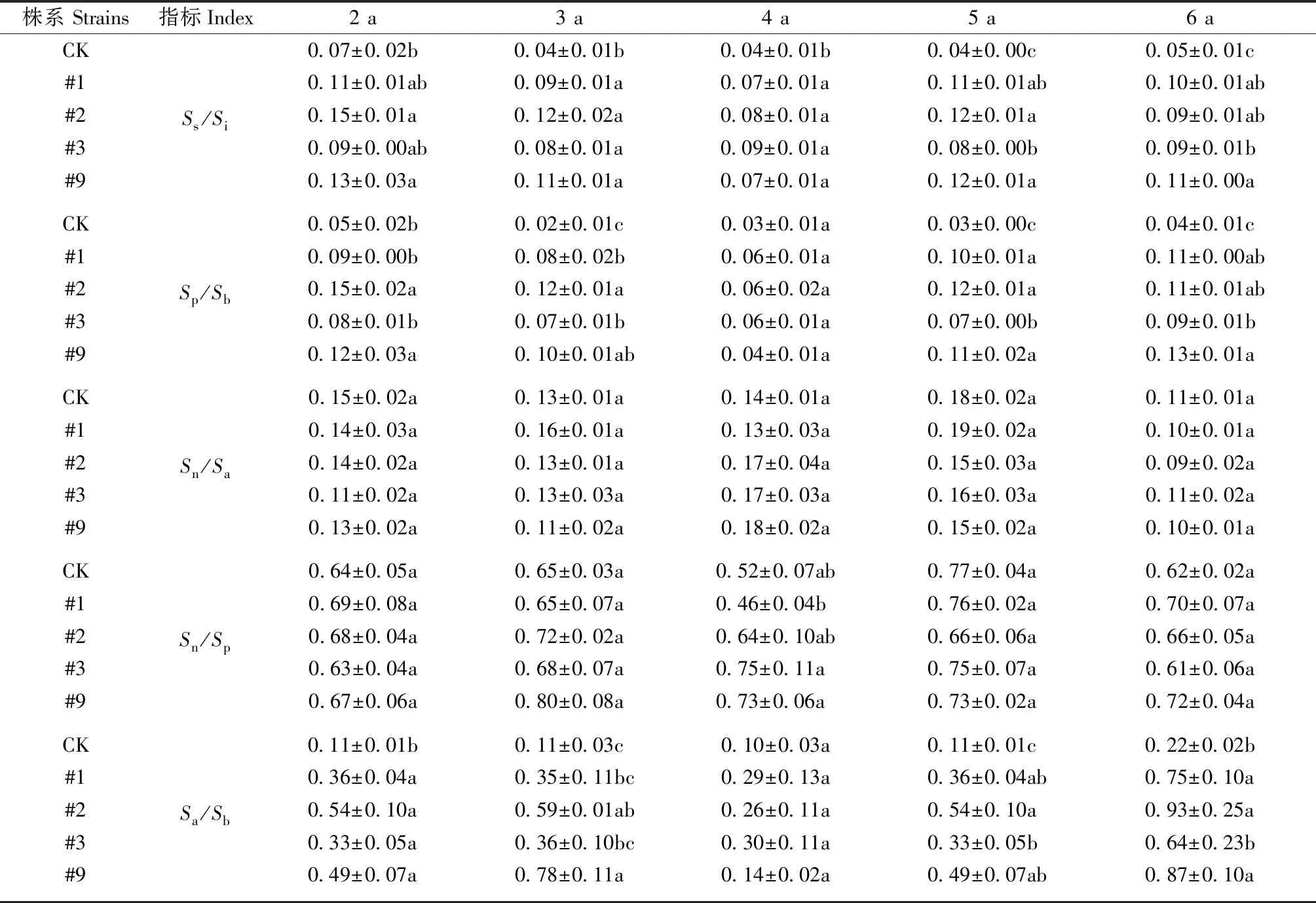
表5 转基因107杨稳定性分析①Tab.5 Stability analysis of transgenic 107 poplars
3 讨论
利用不同方式进行转基因杨树繁殖均具有较高抗虫性,嫁接繁殖还可提高其环境安全性(陈盼飞, 2016),因此本研究以嫁接转基因107杨为对象,通过研究转基因107杨对节肢动物群落多样性及稳定性的影响,为其生态安全性评价提供依据。转入基因能否稳定存在与表达是其能否高效抗虫及保持较高环境安全性的重要因素,且转基因杨树对靶标害虫的抗虫效果是其应用的关键,目前研究表明转不同类型的抗虫基因均对靶标害虫保持较高抗虫性(胡建军等, 2010),且毒蛋白的表达量越高,抗虫效果越好(夏兰芹等, 2000)。本研究表明转抗虫基因杨能有效抑制靶标害虫,对靶标害虫具有持续的抗虫效果,原因可能是由于转入基因中的毒蛋白在植株体内稳定持续高效表达。转基因107杨在抑制靶标害虫的同时,对非靶标生物的影响同样是生态安全性评价的重要内容,任何一个物种的变化都会对其他物种产生直接或间接的影响,进一步对群落产生影响,因此转基因杨树在抑制靶标害虫的同时,对非靶标害虫的影响同样不容忽视。本研究发现同一林龄转基因107杨害虫亚群落个体数量低于对照,天敌及中性节肢动物亚群落物种及个体数量无明显差异,随林龄的变化部分天敌与中性昆虫个体数量表现出一定程度增多,表明转基因107杨对天敌及中性节肢动物亚群落种类与数量未造成不利影响,与Zhang等(2011)研究结果相似,原因可能是由于转入的抗虫基因具有选择性,在抑制靶标害虫的同时对非靶标生物不具有抑制或致死效应。
节肢动物群落的多样性、均匀度指数越高,优势度指数低,表明群落结构越复杂,物种分布越均匀,群落结构更稳定。本研究结果显示转基因107杨节肢动物群落多样性、均匀度指数高于对照,优势度指数低于对照,表明转基因107杨对节肢动物群落未产生显著负面影响,这与他人(姜文虎等, 2018; Zuoetal., 2018; Zhangetal., 2011)的研究结果相似。转基因107杨群落特征指数随年份变化产生波动,年份对其影响较大,差异显著,可能是由于调查期间选取的调查时间、天气、温度等其他因素的影响。对转基因107杨稳定性进行分析,转基因107杨稳定性显著高于对照107杨,表明转基因107杨相对稳定,这与刘洁(2015)的研究一致。
4 结论
转基因107杨对主要靶标害虫具有持续的抗虫效果,对天敌及中性节肢动物群落未产生不利影响,生态安全性相对较高。温度、湿度等气候因素、毒蛋白的表达及其他各种因素同样会对天敌、害虫及中性昆虫的种类与数量产生影响,今后的研究中需结合其他各方面因素进行长期监测,为进一步评价转基因107杨生态安全性提供依据。
