桉树伐桩分解过程中木质纤维素成分的变化特征
2022-07-20邓祥胜李明蔓刘海宇
邓祥胜,李明蔓,何 鹏,杨 梅,申 露,刘海宇,王 超,程 飞
(广西大学 林学院,广西 南宁 530004)
伐根是由林木采伐后剩余的地上部分伐桩和地下部分根系所构成[1]。伐根作为采伐剩余物,是人工林最重要的粗木质残体(Coarse woody debris,CWD),它既是林木自然更新过程萌芽生长和生物量积累的能量来源[2-3],也在水土保持、地力维持和碳吸存等方面具有重要的生态作用[4]。
桉树是我国南方重要的速生用材树种,桉树的种植为保证我国木材安全及天然林保护、林农脱贫致富、乡村振兴等作出了重要贡献。2018 年中国桉树人工林种植面积超过540 万hm2,其中广西桉树速丰林总面积达256 万hm2,约占广西森林总面积的1/6[5]。广西年森林采伐限额达到了3 681.8 万m3,居全国第一,约占全国的1/7,其中桉树采伐限额达2 600 万m3。可见广西每年因桉树采伐而产生的伐根储量巨大。高频率、短周期的轮伐经营和不合理的林地清理方式,导致了桉树人工林在长期连栽后地力下降。因此,伐根等采伐剩余物的分解和养分回归对于人工林立地的改善和可持续经营水平的提高是十分重要的。
一般而言,速生桉在采伐后数年内伐根从固有形态很快转变为无定性的粉末状而进入土壤,而从林地中消失,其分解速度远快于其他树种。这与我国南方高温多雨的气候条件有利于微生物的活动和物质的淋溶密切相关。伴随着形态从有形向无形的转化,伐根的化学结构成分也呈现剧烈的变化。伐根是复杂的木质生物材料,其主要结构成分是纤维素、半纤维素和木质素,及一定量的其他组分[6-7]。一般认为,木材力学性质大多与它的密度有显著相关性,如木材的强度和刚度随密度的增加而增大。木材组成细胞的细胞壁壁薄腔大,则木材密度小;相反,壁厚腔小,则木材密度大。因此,作为木材各种细胞的胞壁成分,纤维素、半纤维素和木质素便与木材的强度和刚性密切相关。纤维素是木材抗弯强度特别高的主要原因,而半纤维素和木质素起着支持纤维素骨架的作用,使木材具有理想的弹性和抗压强度。自然状态下,在微生物(尤其是褐腐菌和白腐菌)的作用下这些木质结构组分经过一系列水解、氧化、脱酸等反应[8]并发生降解,导致木质结构的物理和机械性能降低,甚至失效。可见,木质结构成分在伐根形态的维持中发挥着重要作用。同时,作为主要的碳源,在伐根分解过程中纤维素和木质素含量和比例的变化可通过影响微生物活动与人工林的碳排放过程相偶联。
目前仅有少量有关伐根的研究报道,且集中在伐根储量[9-10]、分解特征[9]、腐殖酸动态[11]以及微生物分解行为和机理[12-14]等方面,而对伐根分解过程中木质纤维素成分的如何变化知之甚少。本研究以不同腐烂等级桉树伐根的地上伐桩为研究对象,通过对伐桩分解过程中木质纤维素含量、纤维素相对结晶度、傅里叶变换红外光谱和纤维形态进行研究,旨在阐明伐桩分解过程中各木质纤维素组分的变化规律,为更好地理解伐根等粗木质残体的分解机制提供科学依据,为人工林地力的维持以及碳中和提供科学参考。
1 材料与方法
1.1 研究区概况
本研究伐桩样品来自广西南宁市南郊的广西国有七坡林场(22°13′47′′N,108°23′26′′E),该区域属南亚热带季风气候区,年均气温为21℃,极端最高气温40℃,最低气温-2℃。由于受季风的影响,旱雨季节明显,年降水量为1 200 ~1 500 mm,多集中在6—9 月,10 月至次年3 月为旱季。地貌属于低丘地形,平均海拔低于500 m,土壤以赤红壤为主,土层较深厚。
1.2 伐桩样品的采集
本研究所需的伐桩样品分别来自七坡林场1、3 和5 年生尾巨桉Eucalyptus urophylla×E. grandis1 代植苗林下,伐桩均为前一代尾巨桉萌芽林皆伐后产生,并在采伐后不久用10%的草甘膦水剂与水按体积比1∶1 配置成处理液对新鲜伐桩表面进行1 次处理,以抑制萌芽产生,加速伐桩死亡。在各桉树人工林下设置一个20 m×20 m 的样方,对样方内的伐桩进行调查和分级。根据闫恩荣等[15]对粗木质残体的5 级分类系统把伐桩分为5 个不同的腐烂等级,结果见表1。在实地调查过程中发现,同一样方中可能同时存在多个不同腐烂等级的伐桩,而同一伐桩上也可能同时存在2 种及以上不同腐烂等级,特别是分解程度高的伐桩较为普遍。因此若某一腐烂等级所占比例达到或超过该伐桩的50%,则将此伐桩归类为该腐烂等级。对于分解程度较轻的伐桩,用油锯从伐桩上端取5 cm 厚圆盘。对于有2 个及以上腐烂等级同时存在的深度分解伐桩,用油锯或刀具等将目标腐烂等级部分小心地从伐桩上取下。所有样品在清除土壤、植物组织等杂质后装入塑封袋中,做好标记并运回实验室。每个腐烂等级采集4 个样品,共20 个实验样品。所有样品在70℃烘至恒质量后用粉碎机粉碎,并分别过40 目和60 目筛网,得到两种粒径的木粉。

表1 森林生态系统粗木质残体的分类系统Table 1 Classification system of CWD in forest ecosystems
1.3 木质纤维素含量
根据Van Soest[16]的方法,利用纤维测定仪确定伐桩样品的木质纤维素含量。准确称取1.00 g木粉样品(通过40 目筛)置于烧杯中,并加入100 mL 中性洗涤剂(配方:分别取18.61 g 乙二胺四乙酸、6.81 g 四硼酸钠、30.00 g 十二烷基硫酸钠、10 mL 乙二醇乙醚和4.56 g 无水磷酸氢二钠配置成1 L 溶液),在5 ~10 min 内煮沸,并持续微沸60 min。煮沸完毕后,倒入已知质量的玻璃坩埚(W2)中进行抽滤,用沸水冲洗坩埚与残渣,洗至滤液中性。用20 mL 丙酮冲洗2 次,抽滤。将玻璃坩埚置于105℃烘箱中烘2 h 后,在干燥器中冷却30 min 称质量。中性洗涤纤维含量(NDF)的计算公式如下:

式中:W1为玻璃坩埚和中性洗涤纤维质量,g;W2为玻璃坩埚质量,g;W为试样质量,g。
酸性洗涤纤维含量的测定与中性洗涤纤维的测定相同,但用酸性洗涤剂(配方:取20.00 g十六烷基三甲基溴化铵,加入0.50 mol/L 硫酸溶液配制成1 L 溶液)替代中性洗涤液。酸性洗涤纤维含量(ADF)的计算公式如下:

式中:HL 为半纤维素含量,%;CL 为纤维素含量,%;NDF 为中性洗涤纤维含量,%;ADF 为酸性洗涤纤维含量,%;R为经72%硫酸处理后的残渣含量,%;AC 为灰分含量,%。
1.4 X 射线衍射分析
本研究中,纤维素相对结晶度被用来表征伐桩分解过程中纤维素结构的变化。取适量已过60目筛网的细粉样品,均匀平铺在有凹槽的玻璃片上,将玻璃片放入X 射线衍射仪(Rigaku D/MAX 2500V,日本)的扫描台,分析木材纤维素的相对结晶度。将扫描范围设置在10°~50°内,扫描速度为10° /min,获得X 射线衍射图。所得结果在Jade 6.0 软件中进行分析。采用Segal[17]法计算纤维素相对结晶度(Cellulose crystallinity index,CrI),计算公式如下:
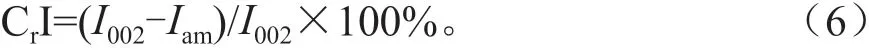
式中:CrI 为纤维素相对结晶度(%);I002为扫描曲线2θ=22.6°处衍射峰强度;Iam为2θ=18.5°处衍射峰强度。
1.5 傅里叶变换红外光谱分析
将干燥KBr 粉末与已过60 目筛网的木粉样品按照质量比为100∶1 的比例混合,然后将混合均匀的粉末在10 kPa 压力下持续1 min,压成直径为13 mm 的小圆盘。利用Thermo Fisher 红外光谱仪(Nicolet IS10,美国)对每一个样品进行扫描,获得光谱。扫描范围400 ~4 000 cm-1,仪器分辨率4 cm-1,扫描采样32 次。所得结果用Omnic 8.0软件分析,对原始光谱进行基线校正和归一化处理。
1.6 木材纤维形态的测定
将不同腐烂等级样品切成火柴杆大小的小木条,并取大小均匀的试样5 根放入试管中。加入冰醋酸和双氧水混合液(体积比1∶1),其中混合液没过试样,利用水浴加热法进行解析。当所有试样颜色变白时即可结束水浴,再用蒸馏水清洗试样,直到醋酸被完全清洗。将试样转移到250 mL锥形瓶中,加入100 mL 蒸馏水,再滴入适量的1%番红溶液震荡染色,用一次性胶头滴管吸取少量上清液制成简易玻片。采用数码显微成像系统(Leica Application Suite V4)对纤维长度、宽度和长宽比进行测定,每个试样随机测定纤维50 根。
1.7 数据处理和分析
运 用Excel 2010、SPSS 20.0、Origin 2020 和Canoco 5.0 软件对实验数据进行统计、方差分析、显著性检验、主成分分析和作图。方差分析之前对数据进行方差齐性检验。用SPSS 20.0 软件中的Levene 检验法检验不同腐烂等级各指标的数据方差是否具有齐性,若P>0.05,则说明指标间数据的方差具有齐性,则可以对数据进行方差分析(本文中不同腐烂等级指标之间的数据方差均具有齐性)。单因素方差分析时,若P<0.05,则特定指标在不同腐烂等级之间具有显著差异,之后进行Duncan 多重比较;若P>0.05,则无显著差异。表中数据为平均值±标准差。
2 结果与分析
2.1 木质纤维素的含量
桉树伐桩木质纤维素含量和比值在不同腐烂等级之间均存在明显差异(P<0.05,表2)。伐桩纤维素含量在分解前期(Ⅰ和Ⅱ)处于较高水平,然后迅速下降,在腐烂等级Ⅴ达到最低;半纤维素含量在伐桩分解过程中呈现降低—升高—降低的趋势,以腐烂等级Ⅲ达到最高;综纤维素含量的变化趋势与纤维素大体类似,其含量随腐烂等级的增加而降低,在分解前、中期(Ⅰ~Ⅲ)受到一定程度的半纤维素影响;腐烂等级Ⅲ木质素含量显著高于其他4 个腐烂等级,分解前期(Ⅰ和Ⅱ)木质素含量升高,在中期(Ⅲ)达到最高后在后期(Ⅳ和V)又明显下降。木质素/综纤维素在不同腐烂等级之间也差异显著(P<0.05),腐烂等级Ⅲ显著高于其他4 个等级,与木质素含量变化趋势相似。桉树伐桩相同腐烂等级的综纤维素、纤维素、半纤维素和木质素含量之间均有显著差异(P<0.05,表3),总体上各腐烂等级均表现为综纤维素>纤维素>木质素>半纤维素。

表2 不同腐烂等级桉树伐桩木质纤维素的含量†Table 2 Lignocellulosic content of Eucalyptus stumps of different decay classes

表3 同一腐烂等级桉树伐桩木质纤维素的含量Table 3 Lignocellulosic content of Eucalyptus stumps of same decay classes %
2.2 纤维素相对结晶度
纤维素相对结晶度随着纤维素结构的变化而变化[18],因此可以较好地反映伐桩分解过程中木质结构的变化。桉树伐桩X 射线衍射典型特征晶面主要出现在14.5°、16.5°、22.6°和34.6°位置(图1),其可以观察到纤维素相对结晶度随着腐烂等级的增加而减少(图1 和表4)。不同腐烂等级之间纤维素相对结晶度差异显著(P<0.05),腐烂等级Ⅰ和Ⅱ显著高于其他3 个等级,以腐烂等级Ⅴ最低(22.32%)。

表4 不同腐烂等级桉树伐桩纤维素相对结晶度Table 4 CrI of Eucalyptus stumps of different decay classes
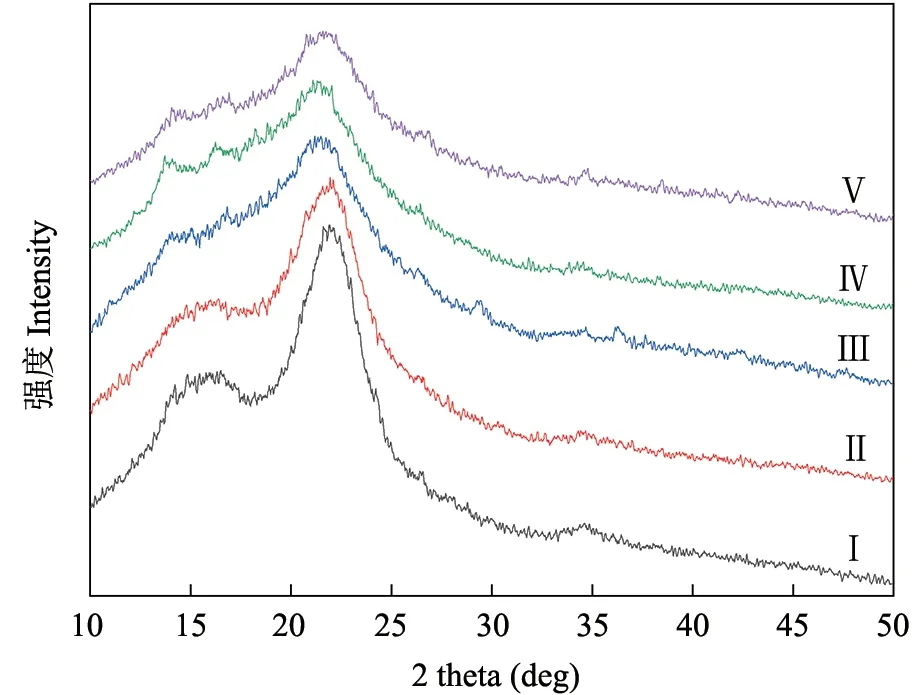
图1 不同腐烂等级桉树伐桩X 射线衍射Fig. 1 X-ray diffraction pattern of Eucalyptus stumps of different decay classes
2.3 木质纤维素红外光谱特征
不同腐烂等级的桉树伐桩傅里叶变换红外光谱如图2 所示。在光谱指纹区800 ~1 800 cm-1范围可以观察到木质纤维素成分特征官能团的振动,因此可以对该区域的红外光谱进行分析,官能团振动类型和对应波长、化学成分见表5[19-20]。不同腐烂等级在1 600、1 330、1 120、1 050 cm-1处振动强度变化不明显,故不作为分析依据,被排除掉。相对于腐烂等级Ⅰ,其他4 个腐烂等级的光谱在1 740、1 506、1 460、1 420、1 370、1 160、898 cm-1处均有明显变化,他们分别对应表5 中半纤维素、木质素、木质素/半纤维素、纤维素、纤维素/半纤维素、纤维素/半纤维素、纤维素等相关官能团及其振动类型。本研究中,谱带在1 506 cm-1处是木质素苯环芳香骨架振动引起,谱带在1 740、1 370、1 160、898 cm-1处是碳水化合物(纤维素和半纤维素)特定官能团振动引起。

表5 不同腐烂等级桉树伐桩红外光谱官能团类型和化学成分Table 5 Functional groups of infrared spectroscopy and chemical components of Eucalyptus stumps of different decay classes
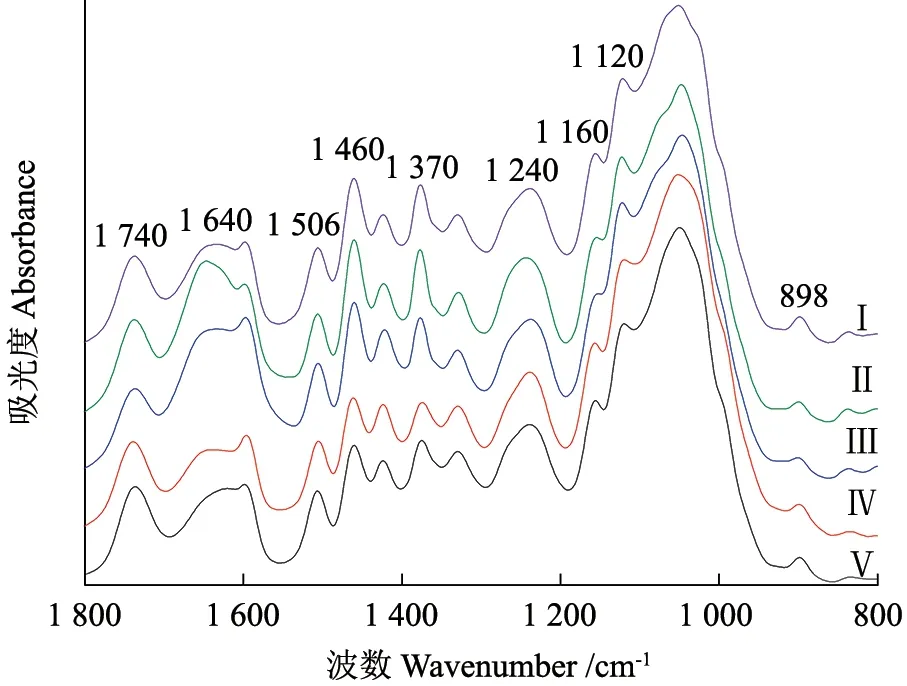
图2 不同腐烂等级桉树伐桩红外光谱Fig. 2 FTIR spectra of Eucalyptus stumps of different decay classes
木质素与纤维素、半纤维素的特征吸收峰的峰高比值可用来判定木质纤维素相对含量的高低[21-22]。不同腐烂等级桉树伐桩红外光谱特征峰相对强度变化如表6 所示。通过木质官能团峰高比值来判定木质素、纤维素和半纤维素相对含量高低时,采用I1506/I1740特征峰比值表征木质素相对含量,采用I1740/I1506、I1370/I1506、I1160/I1506和I898/I1506特征峰比值表征纤维素和半纤维素相对含量。由表6 可 知,I1740/I1506、I1370/I1506、I1160/I1506和I1506/I1740特征峰比值在不同腐烂等级之间具有显著差异(P<0.05)。总体来看,随着伐桩腐烂等级的增加,I1740/I1506、I1370/I1506和I1160/I1506均呈现先减小后增大的趋势,I1506/I1740则随着腐烂等级的增加呈先增大后减小的趋势,而I898/I1506各腐烂等级间没有显著差异。可见,在整个伐桩分解过程中其木质纤维素相对含量均在减少。分解后期(Ⅳ和Ⅴ),I1740/I1506、I1370/I1506和I1160/I1506有所增加,而I1506/I1740则减小,表明此时木质素相比半纤维素和纤维素有着更快的分解速率。

表6 不同腐烂等级桉树伐桩红外光谱特征峰相对强度Table 6 Relative intensity of characteristic peaks of infrared spectrum for Eucalyptus stumps of different decay classes
2.4 纤维形态特性
不同腐烂等级桉树伐桩的纤维长度、宽度和长宽比均具有显著差异(P<0.05,表7)。腐烂等级Ⅰ的纤维长度和宽度显著高于其他腐烂等级,并随腐烂等级的增加而减小,纤维长度以腐烂等级Ⅳ和Ⅴ最小,而宽度以腐烂等级Ⅴ最小。纤维长宽比随腐烂等级的增加总体呈减小的趋势,但以腐烂等级Ⅴ最大。

表7 不同腐烂等级桉树伐桩纤维形态特征Table 7 Fiber morphological characteristics of Eucalyptus stumps of different decay classes
2.5 木质纤维素因子与伐桩腐烂等级之间的关系
主成分分析的4 个轴对伐桩木质纤维素因子与腐烂等级之间的关系的总解释度高达95.10%,其中前两个主成分共解释总变异的69.42%,第一主成分解释量占45.14%,第二主成分占24.28%,前
两轴对伐桩木质纤维素因子与腐烂等级之间的关系有较强的解释能力(图3)。第一主成分与相对结晶度、纤维素、纤维长度和纤维宽度的相关性较强,而木质素、半纤维素和纤维素长宽比与第二主成分的相关性较强。其中腐烂等级Ⅰ和Ⅱ与相对结晶度、纤维素、综纤维素、纤维素长度和纤维素宽度呈正相关,与木质素/综纤维素呈一定程度的负相关;腐烂等级Ⅲ和Ⅳ与木质素、半纤维素呈正相关,与纤维素长宽比呈负相关;腐烂等级Ⅴ与纤维素长宽比呈正相关,与木质素和半纤维素呈负相关。从图3 也可以看出,不同腐烂等级桉树伐桩根据纤维素因子的数量特征可以被分成3 组,即腐烂等级Ⅰ和Ⅱ的桉树伐桩,其木质纤维素因子数量特征更为相似,可分为一组;腐烂等级Ⅲ和Ⅳ的桉树伐桩可成为一组;腐烂等级Ⅴ则单独为一组。

图3 伐桩木质纤维素因子与腐烂等级的主成分分析Fig. 3 Principal component analysis of lignocellulosic factors and decay classes
3 讨 论
3.1 桉树伐桩木质纤维素结构组分
纤维素、半纤维素和木质素是伐桩的主要组分。本研究中,伐桩分解前期(腐烂等级I 和II)半纤维素优先被降解,这与其他褐腐木材降解结果类似[21,23-26]。在植物细胞壁中,纤维素微纤丝主要嵌入半纤维素、果胶多糖和木质素中[6,27]。半纤维素的降解先于纤维素的原因在于纤维素微纤丝被半纤维素包裹,而半纤维素是以糖苷键连接的无水单糖聚合物(主要是木糖和甘露糖)[28],因此微生物优先分解半纤维素中的木糖和甘露糖[29]。当纤维素外围的半纤维素包膜被降解,便成为微生物入侵纤维素的入口,进一步导致纤维素降解。纤维素由纤维二糖单体组成,其结构简单,容易被褐腐菌优先降解[30-32],导致腐烂等级Ⅰ~Ⅲ的综纤维素持续减少。木质素和木质素/综纤维素与其呈相反的变化,除了与在该阶段纤维素和半纤维素被分解有关外,还与木质素的相对增加(富集)有关。木质素具有比纤维素相对更为复杂的结构,分解较慢而引发富集,Fravolini 等[33]发现纤维素的分解率是木质素的10 倍。崔福星等[34]也发现随着落叶松倒木腐烂等级的增加,纤维素和半纤维素含量在逐渐减少,而木质素则呈现积累状态。其他有关粗木质残体分解的研究也能得到类似的结果[31,35-38]。由于微生物胞外酶优先分解纤维素,导致木质素累积,被认为是森林土壤稳定碳源的重要形成机制[31]。白腐菌在伐桩分解中后期逐渐占优势,其对木质细胞壁成分分解不具有选择性,可以分解纤维素、半纤维素、木质素等,因此造成腐烂等级Ⅲ~Ⅴ的木质纤维素含量均减少,致使伐桩很快失去了形态支撑。但需要注意的是,白腐菌通过分泌一系列胞外过氧化物酶和漆酶对木质素有更强的分解能力[39-40]。
3.2 桉树伐桩纤维素相对结晶度
纤维素结构可分为结晶区和非结晶区,结晶区有较高的密度和较低的化学反应活性[41]。纤维素相对结晶度是指纤维素的结晶区质量(或体积)占纤维素整体质量(或体积)的百分数。相对结晶度随着纤维素两个结构区的占比以及整体质量(或体积)的变化而变化,从而影响到木材的抗拉强度、抗弯强度和稳定性。一般来说,纤维素结晶区占比越高,相对结晶度越大,抗拉强度、抗弯强度和稳定性也越高。本研究中,伐桩纤维素相对结晶度随着腐烂等级的增加而降低,表明随着腐烂的进行,纤维素结晶区占比逐渐降低,伐桩木质结构稳定性下降,形态逐渐从有形向无形转化。
伐桩分解前期(腐烂等级Ⅰ和Ⅱ)半纤维素的优先降解对纤维素影响较小,纤维素相对结晶度仅仅减少了0.49%,而这种减少主要源于微生物对纤维非结晶区的攻击。有研究表明,相比纤维素结晶区,真菌更容易侵染纤维素非结晶区,而使之成为真菌侵染结晶纤维素的切入点[42]。
随着腐烂进一步加深,相对结晶度显著下降到22.32%(腐烂等级Ⅴ),表明纤维素结晶区遭受到强烈降解,这与白腐菌和褐腐菌向微环境中分泌大量有机酸(如草酸)促进芬顿化学反应有关[40]。综合化学成分和相对结晶度结果可以推测,微生物首先侵袭伐桩细胞壁半纤维素和纤维素非结晶区,再以此为突破口进一步深度侵袭细胞壁,导致细胞壁严重损坏。
3.3 桉树伐桩木质纤维素红外光谱
Yilgor 等[43]和Zhang 等[44]认为傅里叶变换红外光谱峰值变化是物质成分变化的良好反映,它的峰值强度可以与纤维素、半纤维素和木质素的改性、退化以及原子之间键的强度联系起来。红外光谱中,碳水化合物和木质素特征峰的强度比(碳水化合物/木质素)的降低表明真菌分解纤维素和半纤维素的速率比木质素要快[45]。本研究中,腐烂等级Ⅰ~Ⅲ碳水化合物(I1370、I1160和I898)和木质素(I1506)的比值I1370/I1506、I1160/I1506、I898/I1506均呈变小的趋势,说明在这个过程中纤维素和半纤维素相对木质素被优先分解,这与本研究的化学成分分析结果相一致。在此过程中,腐烂等级I 和II中半纤维素(1 740 cm-1)和木质素(1 506 cm-1)的 比 值I1740/I1506明 显 比I1370/I1506、I1160/I1506、I898/I1506降低更多,说明在这个过程中半纤维素又较纤维素优先分解,也与本研究的化学成分分析结果一致。值得注意的是,I1740/I1506在腐烂等级Ⅰ~Ⅲ中呈先减小后增大的趋势,是由于1 740 cm-1处峰高呈先减小后增大引起,1 740 cm-1处峰高增加是褐腐菌和白腐菌向微环境释放出的酸与细胞壁中的羟基发生了醚化反应,使木质素分解形成羰基,说明木质素结构在腐烂等级Ⅱ时就开始受到褐腐菌的侵蚀破坏。腐烂等级Ⅲ~Ⅴ的I1740/I1506、I1370/I1506、I1160/I1506、I898/I1506比值均呈增大的趋势。然而,纤维素和半纤维素峰高却呈降低的趋势,说明二者在该阶段存在持续分解。因此上述比值增大主要是因为在该阶段木质素的分解较其他两种组分更加强烈,同时也表明木质素的强烈降解开始发生在腐烂等级Ⅲ,这与本研究的化学分析结果相一致。木质素相对含量的迅速减少与白腐菌可以分解纤维素和半纤维素但对木质素有更强分解能力的特征相一致。
3.4 桉树伐桩纤维形态特征
木材纤维形态特征是木材物理和力学性质的决定因素。木材纤维长度和宽度体现木质材性的优劣。纤维宽度越大,细胞壁越薄,纤维间的接触面积越大,木质的交织力和粘结性就越强,木质强度就越高。纤维长宽比越小,则说明木质纤维相互交织的数量越少,交织能力越弱,也说明木质强度和硬度减小。本研究中,伐桩纤维长度、宽度和长宽比均随着腐烂等级的增加而减小,表明伐桩木质强度、力学等物理性质随分解而逐渐下降,从而导致伐桩逐渐失去其固有的形态支撑。
综上,桉树伐桩木质组分在不同腐烂等级之间具有差异,分解初期主要是半纤维素和纤维素等易分解物质的降解,而在分解后期所有木质组分均被分解。此外,本研究还指出伐桩的分解与微生物的活动密切相关,然而本研究并未对不同腐烂等级伐桩木质组分与微生物多样性和群落结构之间的关系进行探讨,该方面的工作还有待进一步加强。
4 结 论
1)根据木质纤维素含量的变化,桉树伐桩分解过程可以分为两个阶段。第一阶段是腐烂等级Ⅰ~Ⅲ,综纤维素选择性被分解,其中半纤维素优先分解,木质素被富集;第二阶段是腐烂等级Ⅲ~Ⅴ,木质素和综纤维素同时被分解。
2)腐烂等级Ⅰ和Ⅱ纤维素尚未发生分解,但非结晶区遭到侵袭而引起相对结晶度下降。随着腐烂程度的增加,木质纤维素含量随之减少,桉树伐桩的相对结晶度也显著减少。
3)随着腐烂等级的增加,综纤维素与木质素红外光谱特征峰的峰高比值呈现先减少后增加的变化趋势。
4)伐桩纤维长度、宽度和长宽比总体上均随着腐烂等级的增加而减小。
5)主成分分析显示,由于腐烂等级Ⅰ和Ⅱ的伐桩其木质纤维素因子较为相似被分为一组,而腐烂等级Ⅲ和Ⅳ的伐桩则被分为另一组,腐烂等级Ⅴ的伐桩单独为一组。
