玉米和高粱褐色中脉基因的研究进展
2022-07-14李杰勤孟瑞瑞王丽华刘言龙王小雨
李杰勤, 孟瑞瑞, 王丽华, 刘言龙, 王小雨, 刘 萍
(安徽科技学院 农学院,安徽 凤阳 233100)
褐色中脉突变体(brown midrib,bmr)是指叶脉和茎杆木质部呈现棕灰或棕褐色的自然或化学突变体[1],一般在植株生长到4~6叶时在叶脉上可见,植株完全成熟后仅在茎秆木质部可见[2]。研究表明,叶片中脉颜色与木质素含量和组成有着密切关系[2],而总木质素含量和木质素单体的组成比例对细胞壁的可消化性影响较大[3]。因此,一般认为褐色中脉的颜色越深,其饲用品质也越好,褐色中脉突变体作物作为生物质能原料,其乙醇转化率也更高[4]。
木质素是存在于植物体内的天然有机大分子,其生物合成对植物生长发育有重要意义,在增强植物细胞壁的机械支持,提高植物体水分运输效率及增加对病菌微生物防御中起重要作用[5-7]。已经克隆出来的5个玉米褐色中脉突变体和3个高粱褐色中脉突变体均被证明与木质素的生物合成相关[5]。因此,研究褐色中脉突变体的木质素生物合成途径相关酶基因,能够更好地了解褐色中脉突变体的产生规律和木质素生物合成及调控机理。
本研究总结了玉米(Maize)和高粱(Sorghum)褐色中脉突变体的类型、木质素的生物合成途径及玉米和高粱褐色中脉基因研究进展,同时对褐色中脉突变体产生的原因进行讨论,以期为玉米和高粱育种应用及褐色中脉突变体的进一步研究提供理论基础。
1 玉米和高粱褐色中脉突变体的类型
1924年在玉米中发现了首个自然突变的褐色中脉突变体,随后在高粱和珍珠粟中也发现相似的突变体[8-9]。迄今为止,在玉米中共发现13个突变体[1,10-13]。根据Ali等[13]的等位性测验,可以将玉米褐色中脉(bm)突变体划分为6个不同的等位基因(表1)bm1、bm2、bm3、bm4、bm5和bm6。其中突变体bm1~bm6分别被定位到第5、1、4、9、5和2号染色体上[14-19]。它们均为自然突变,属于单基因隐性突变[13]。与各自野生型相比,玉米bm1、bm2、bm3、bm4、bm5不仅降低了木质素含量还改变了木质素的组成,而bm1、bm2、bm3、bm4、bm5只降低了木质素含量并没有改变木质素的组成(表1)。
Porter等[8]、Bittinger等[20]和Sattler等[21]用甲基磺酸乙酯(MES)和硫酸二乙酯(DES)对高粱进行化学诱变,共产生20个不同的突变体,通过等位性测验,Sattler等[21]将高粱褐色中脉突变体(bmr)划分为8种不同的等位基因(表1),其中bmr2、bmr6和bmr12分别被定位到第2、3和7号染色体上[22-23],而其它类型突变体并未被定位。高粱褐色中脉突变体与其野生型相比,木质素含量明显降低,其中高粱bmr2、bmr6和bmr12在降低了木质素含量的同时还改变了木质素的组成;其他类型仅木质素含量降低,但组成并未发生改变。
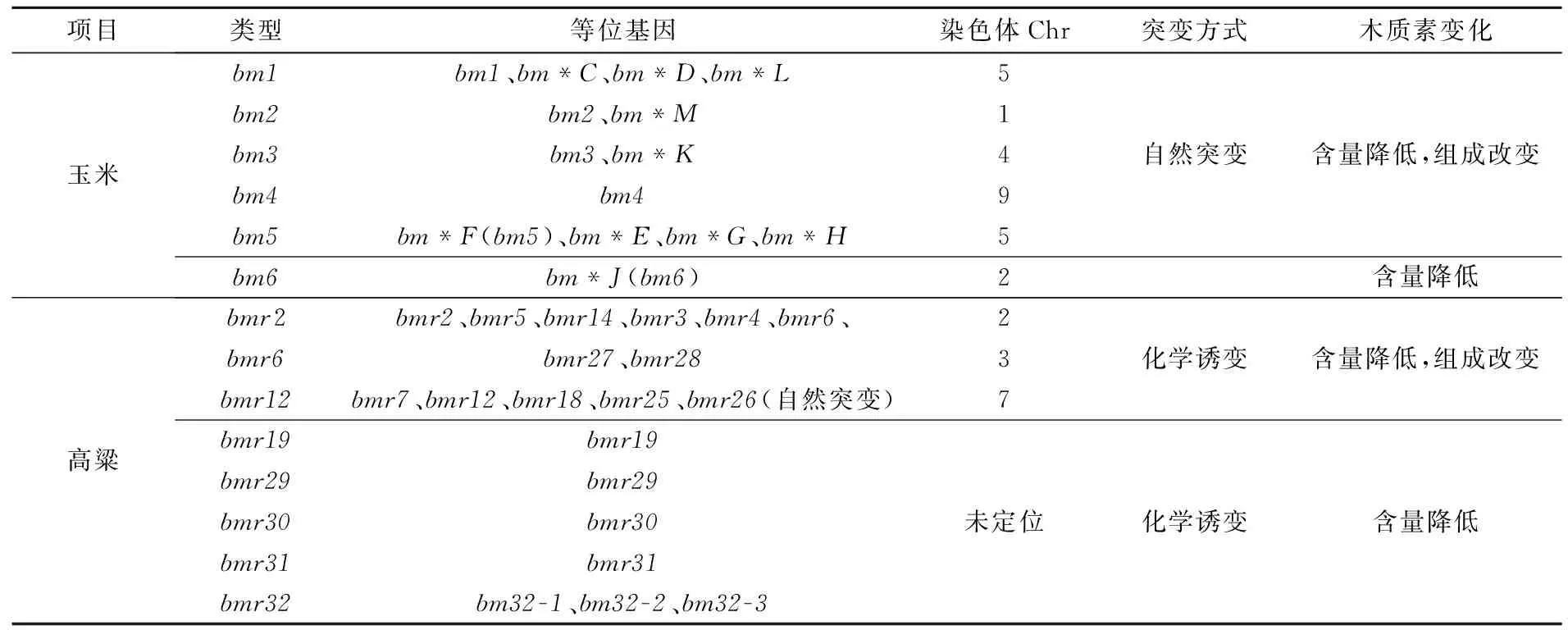
表1 高粱与玉米褐色中脉突变体类型
2 木质素类型及生物合成途径
木质素是存在于植物体内的天然有机大分子,是一种由肉桂醇等单体聚合生成的酚类多聚物,木质素沉积在木质部导管和厚壁组织及韧皮部纤维中,在增强植物细胞壁的机械支持,提高植物体水分运输效率及增加对病菌微生物防御具有重要作用[5-7]。木质素的生物合成分主要分为两步,第一步是木质素单体的合成,第二步为木质素单体通过脱氢聚合合成高分子化合物[24]。木质素单体是由苯丙氨酸至少通过11种不同的反应途径衍生而成[25]。苯丙氨酸在苯丙氨酸裂解酶(PAL)作用下通过非氧化脱氨生成反式肉桂酸,肉桂酸-4-羟基化酶(C4H)在上一步的反应产物反式肉桂酸催化下生成对羟基香豆酸。对羟基香豆酸在香豆酸-3-羟化酶(C3H)、香豆酸-3-羟化酶(COMT)、阿魏酸-5-羟化酶(F5H)等酶作用下逐步生成咖啡酸、阿魏酸、5-羟基阿魏酸和芥子酸等羟基苯乙烯酸。4-香豆酸辅酶A连接酶(4CL)把这些不同结构的羟基苯乙烯酸乙酰化生成对应的羟基肉桂酰-CoA酯,各类羟基肉桂酰-CoA在肉桂酰CoA还原酶(CCR)作用下生成相应的肉桂醛,肉桂醇脱氢酶(CAD)在木质素单体合成的最后一步起作用,催化上一步产物肉桂醛生成相应的肉桂醇,肉桂醇会在漆酶和过氧化酶的作用下聚合生成各种木质素[26](图1)。高等植物的木质素主要可以分为3类:对-羟苯基木质素(H-木质素)、愈创木基木质素(G-木质素)和紫丁香基木质素(S-木质素)。H-木质素来源于松柏醇,G-木质素来源于芥子醇,而S-木质素来源于对羟基香豆醇[27]。木质素生物合成途径的变化导致木质素含量降低和组成成分改变,从而使玉米和高粱产生褐色中脉突变体。
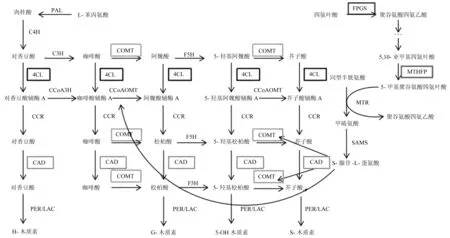
图1 木质素生物合成途径
3 玉米褐色中脉突变体(bm)基因克隆
玉米褐色中脉突变体bm1~bm6中,bm1~bm5已经被证明与木质素生物合成直接或间接相关。其中bm1编码CAD基因,bm2编码MTHFR基因,bm3编码COMT基因,bm4编码FPGS基因,而bm5编码4CL基因(表2)。
1995年,Vignols等[28]克隆了第一个玉米褐色中脉基因bm3,即编码咖啡酸-3-O-甲基转移酶的基因(COMT)。COMT是进化上保守的甲基转移酶家族的成员,其在木质素合成中的作用在单子叶植物和双子叶植物中均有记载[28]。在木质素生物合成途径中,COMT将S-腺苷-蛋氨酸(SAM)的甲基转移到5-羟基松柏醛和5-羟基松柏醇上,从而生成相应的芥子醛和芥子醇(图1)。Vignols等对玉米bm3两个等位基因突变体进行测序分析,发现其中一个等位基因是因为COMT基因内含子编码区的连接处插入了一个与B5元件同源的逆转录转座子,而造成COMT活性丧失,而另一个则是因为部分COMT基因的缺失[28]。

表2 玉米和高粱褐色中脉突变体基因克隆研究进展
随后,Halpin等[29]通过同样的方法,克隆了玉米褐色中脉基因bm1,即编码肉桂醇脱氢酶(CAD)基因,bm1类型的重要特征是茎杆木质部CAD活性的丧失,与野生型相比,bm1的总木质素含量下降约20%,木质素的组成也发生了变化。在木质素合成途径中,CAD以NADPH为辅助因子,催化肉桂醛生成相应的醇(表2)。bm1突变体中的对羟基苯基(H)、愈创木基(G)和丁香基(S)都显著减少,并增加了松柏醛和芥子醛以代替其相应的醇。bm1的蛋白提取物中,ZmCAD2蛋白显著降低,证明了玉米bm1编码玉米ZmCAD2。
2014年,Tang等[16]利用候选基因法克隆出bm2基因,酵母互补实验证明bm2编码亚甲基四氢叶酸还原酶(MTHFR),参与关键甲基供体S-腺苷-L-蛋氨酸(SAM)的形成,SAM是COMT和CCOAOMT的甲基供体,把MTHFP和木质素生物合成联系起来。与其野生型相比,bm2突变体积累的木质素含量降低,组成也发生改变,降低的木质素主要为G-木质素和S-木质素。
2015年,Li等[17]利用物理定位法克隆了bm4基因,通过酵母互补实验证明bm4编码功能性FPGS,bm4突变体导致木质素含量下降和木质素中丁香基(S)和愈创木基(G)比值升高。玉米bm4突变体的发现证明了FPGS在木质素生物合成中的作用。
Haney等[14]鉴定出bm5基因,Mechin等[18]对bm5进行化学分析,发现玉米bm5突变体中G-木质素含量显著下降,H-木质素含量明显提高,而对S-木质素的生物合成影响不大。随后,Xiong等[30]通过MutMap基因定位法,确定bm5基因座编码一个木质素单体生物合成途径关键酶之一:4-香豆酸:辅酶A连接酶(Zm4CL1)。bm5突变体测序分析发现,4CL活性的降低是由于转座子的插入。Zm4CL1基因突变主要影响玉米木质化组织中G-木质素的生物合成和可溶性阿魏酰衍生物的积累,但不影响玉米的生长发育。此外,bm5突变体的中性洗涤纤维消化率和细胞壁糖化效率均显著提高。bm5突变体的抗逆性降低表明Zm4CL1是细胞壁工程的一个优良靶点,对该基因的遗传操作将有利于农作物秸秆和秸秆的利用[30]。
4 高粱褐色中脉突变体(bmr)基因克隆
在已知的8个不同类型的高粱褐色中脉突变体中,bmr2、bmr6和bmr12已被鉴定,并参与木质素的生物合成途径。它们分别编码木质素单体生物合成途径中4-香豆酸:辅酶A连接酶(4CL)、肉桂醇脱氢酶(CAD)和咖啡酸O-甲基转移酶(COMT)[22,31-32](表2)。
2003年Bout等[31]利用候选基因法从高粱中克隆了首个褐色中脉基因,结果显示,bmr12、bmr18和bmr26都为编码COMT的等位基因上出现点突变而使COMT基因失活。在木质素生物合成途径中,COMT优先催化木质素合成途径中的5-羟基松柏醛和5-羟基松柏醇甲基化生产芥子醛和芥子醇,参与S-木质素的形成。Palmer等[33]研究发现,COMT活性降低降低了S-木质素的含量并增加了5-OH木质素含量。
2010年,Sattler等[34]发现bmr6突变体中木质素3种亚基:对羟基苯基(H)、愈创木基(G)和丁香基(S)均减少。随后Li等[17]克隆出bmr6基因,基因互补实验证明bmr6编码肉桂酸脱氢酶(CAD2),CAD是乙醇脱氢酶超家族的一员,在木质素生物合成途径中,利用NADPH作为辅助因子将对羟基肉桂醛还原生成相应的醇[32,35]。与野生型相比,bmr6突变体中木质素含量明显下降,但醛和多糖含量升高,表明bmr6细胞壁中醛和多糖取代了木质素单体(香豆醇、松柏醇和芥子醇)[17]。
2012年,Saballos等[22]克隆出bmr2基因,实验证明bmr2编码木质素生物合成途径中的4-香马酸:辅酶A连接酶(4CL)。与野生型相比,高粱bmr2降低了木质素的含量,同时木质素组成的也发生了变化。bmr2突变体的木质素显示愈创木基和丁香基残基均减少。
到目前为止,已经鉴定出8个褐色中脉基因,5个在玉米中,3个在高粱中。其中bm1和bmr6,bm3和bmr12,bm5和bmr2代表了玉米和高粱的3个同源基因座,分别编码CAD、COMT和4CL基因(表2),并参与木质素的生物合成。而bm2和bm4通过影响COMT和CCoAOMT合成所需的甲基供体SAM而被并入木质素生物合成途径中(图1)。
5 褐色中脉突变体研究过程中存在的问题和展望
随着研究的不断深入,褐色中脉突变体相关基因逐步被克隆出来。但现在仍然有许多难题亟待解决。比如,褐色中脉突变体产生褐色中脉的原因尚未清楚。最初的研究发现,褐色中脉突变体的色素沉积着局限于木质化组织,而木质化组织不易处理。主要是因为木质素生物合成途径十分复杂,过程中涉及十几种酶系参与反应。从理论上来说,合成途径中任何一步受到干扰都有可能影响木质素的产生。但早期生化分析表明,褐色中脉的色素沉积不是由于类胡萝卜素、花青素、黄酮、单宁或黄酮醇的积累[1]。有趣的是,当木质素生物合成发生突变或受到RNA干扰技术的影响时,能够在维管植物(从双子叶植物到裸子植物)中观察到木质化组织的异常褐色染色[36-40]。这些结果表明,木质化组织的颜色改变,是由于木质素生物合成途径中几个不同步骤的中断引起的,并不是针对于维管植物的特定的某个步骤的变化[27,36-40]。
此外,木质素组分的特殊变化不容易与褐色色素沉积联系起来,因为这些突变体和转基因株系在木质素生物合成途径的不同步骤中受损,从而导致木质素组成成分发生变化[27,36-40]。肉桂醇脱氢酶(CAD)活性受损的突变体或转基因系是例外,其中木质化组织呈现出棕色是由于肉桂醛取代肉桂醇并入木质素中[27,37-39,41]。对于在木质素生物合成的其它步骤中受损的突变体和转基因株系,有人认为颜色的改变是由于将香豆醇、松柏醇或芥子醇以外的酚类化合物并入木质素。虽然颜色变化的确切原因尚不清楚,但褐色中脉突变体似乎是C4禾草中木质素生物合成受损的一个良好标志。
褐色中脉作为木质素生物合成途径反应重要标识,对研究植物木质素生物合成及调控的相关基因具有重要意义,而CRISPR/Cas9系统作为一种快速发展的基因编辑技术体系,具有制备简单、靶向性强、成本低和脱靶率低等诸多优势,成功应用到了微生物、植物和动物中[42]。在植物中,CRISPR/Cas9系统可以改造植物基因组序列、定点修饰目标基因、验证基因功能[43-47]。目前为止,除4CL(bm5和bmr2)、COMT(bm3和bmr12)、CAD(bm1和bmr6)、MTHFR(bm2)和FPGS(bm4)之外,木质素生物合成途径其他酶相关的褐色中脉突变体未见报道,利用CRISPR/Cas9技术对木质素生物合成途径中的其他基因进行定向敲除,是用来研究褐色中脉突变体与植物木质素生物合成及调控相关基因的一种新思路。