捕食性天敌叉角厉蝽生长发育、繁殖及各虫态形态特征观察
2022-07-14张曼高平赵航周辰彦梁晨汤永玉邢孔政吴国星高熹
张曼 高平 赵航 周辰彦 梁晨 汤永玉 邢孔政 吴国星 高熹



摘要:【目的】明確叉角厉蝽[Eocanthecona furcellata (Wolff)]从卵到成虫发育历期及繁殖能力,比较叉角厉蝽各虫态的主要形态鉴别特征,为叉角厉蝽的扩繁及生防潜能开发提供理论支持。【方法】在室内条件下,以云南元江采集并在室内扩繁6代以上的叉角厉蝽为研究对象,在培养皿中采用单头饲养的方法测定叉角厉蝽生长发育指标(卵孵化率、若虫存活率、卵到成虫的发育历期),在饲养盒中雌雄虫配对共培养测定其成虫繁殖力特征。运用显微照相系统比较叉角厉蝽各虫态的主要形态特征。【结果】叉角厉蝽从卵到成虫的平均发育历期为29.00±1.41 d,卵历期为 8.00±0.21 d,若虫期为19.76±0.12 d,1~5龄若虫平均发育历期为3.24~5.38 d,卵孵化率为92.91%,若虫存活率为80.21%,雌、雄成虫的平均寿命分别为35.40±1.96和37.40±2.65 d;叉角厉蝽雌成虫交配后,一生平均产卵次数为 4.73±2.01次,最多9次,最少2次,平均产卵前期为7.87±2.75 d,产卵期持续14.33±5.13 d,雌成虫与雄成虫交配单次平均产卵量为59.46±15.77粒,未交配雌成虫单次平均产卵量为29.34±15.31粒且为无效卵,最终无法孵化。叉角厉蝽卵为矮杯形或圆筒形,具卵盖,边缘有10~12根刺状精孔突,有金属光泽;1~5龄若虫体色红黑相间,若虫触角4节。1龄若虫触角第2和第3节末端有明显白色环,喙暗红色,接近体长,无翅芽;2龄若虫头部前段扁平突出,触角节间环变为红色,喙黑色;3龄若虫触角第2和第3节末端红色环明显,喙长为体长的一半;4龄若虫喙长为体长的一半,胸部刻点明显,出现翅芽,并延伸至胸部末端;5龄若虫喙暗红色,短于体长一半,中足和后足胫节中部出现白斑,翅芽延伸至腹部第3节;成虫体色黄褐与黑褐混杂相间,密布刻点,触角5节,喙黄褐色,最后一节黑色,喙长短于体长的一半,前胸背板侧角呈剑叉状突出,雄虫腹部近三角形,雌虫腹部卵圆形。【结论】明确了叉角厉蝽的发育历期和繁殖能力,证实叉角厉蝽无孤雌生殖现象。农业生产中可以喙的颜色、长度及翅芽发育作为叉角厉蝽主要形态鉴别特征。
关键词: 叉角厉蝽;发育历期;繁殖力;孤雌生殖;形态特征
中图分类号: S467.2 文献标志码: A 文章编号:2095-1191(2022)04-1078-10
Development, fecundity and morphological characteristics of the predatory Eocanthecona furcellata (Wolff)
ZHANG Man, GAO Ping, ZHAO Hang, ZHOU Chen-yan, LIANG Chen,
TANG Yong-yu, XING Kong-zheng, WU Guo-xing, GAO Xi*
(College of Plant Protection, Yunnan Agricultural University, Kunming,Yunnan 650201, China)
Abstract:【Objective】To determine the developmental duration and fecundity of Eocanthecona furcellata (Wolff) from egg to adult and to compare the main morphological distinguishing characteristics of the various insect states of E. furcellata, so as to provide theoretical support for propagation and development of biocontrol potential of E. furcellata. 【Method】Under laboratory conditions,E. furcellata collected in Yuanjiang County of Yunnan Province and propagated for more than six generations in the laboratory were taken as the research object. Growth and development indexes (egg hatching rate, nymph survival rate and development period from egg to adult) of the bugs were determined by single-head feeding in petri dish method, and the characteristics of adult fecundity were determined by paired co-culture of males and females in the feeding box. In addition, the main morphological characteristics of each insect state were compared using microscopic photography system. 【Result】The average developmental period from eggs to adults of E. furcellata was 29.00±1.41 d, the egg period was 8.00±0.21 d, and the nymph period was 19.76±0.12 d. The average developmental period from 1st to 5th instar was 3.24-5.38 d, the egg hatching rate was 92.91%, and the nymphs survive rate was 80.21%. The average lifespans of the female and male adults were 35.40±1.96 d and 37.40±2.65 d respectively. After mating, the number of times of egg production was 4.73±2.01 times, up to 9 times, at least 2 times and the average pre-oviposition stage was 7.87±2.75 d, and the spawning period lasted 14.33±5.13 d. The single fecundity of each female adult mating with a male adult was 59.46±15.77 eggs, and the single fecundity of the female adult without mating was 29.34±15.31 eggs, which were invalid eggs and could not be hatched. Eggs of E. furcellata were short cup-shaped or cylindric, with egg caps and 10~12 prickles on the edge and metallic luster; 1st to 5th instar nymphs were often red and black, antennae of the nymphs were 4 segments. The 1st instar nymphs had obvious white rings at the end of 2nd and 3rd segments of antennae and dark red beaks, its length was close to body length and it has wingless buds. The 2nd instar nymphs with flat protruding anterior head segment, antennal ring turned red, and beak was black. The 3rd instar nymphs had obvious red rings at the end of 2nd and 3rd segments of antennae and the beak length was half of body length. The beak of the 4th instar nymph was half as long as the body, and with marked punctures on chest, the wing buds extending to the end of the chest. The 5th instar nymphs with dark red beaks, and the beak length was shorter than half body length, middle and hind feet had white spots in middle part, wing buds reaching to the 3rd segment of abdomen. Adults’ body usually were yellowish-brown and dark-brown-mixed, with dense punctures, and their abdomen had 5 segments, and their beak was yellowish-brown and the last segment of black, beak length of half of the body length, the lateral angle of the anterior thoracodorsal plate was prominent as a sword fork, male abdomen nearly triangular and female abdomen was oval. 【Conclusion】The developmental period, fecundity and no parthenogenesis phenomenon of E. furcellata are clarified. The beak color, length and wing bud developmental stage are the main characteristics for distinguishing E. furcellata morphology.
Key words:Eocanthecona furcellata(Wolff); developmental period; fecundity; parthenogenesis; morphological characteristics
Foundation items: Yunnan Basic Applied Research Plan Project (202001AT070139)
0 引言
【研究意义】捕食性天敌昆虫是自然界中天敌资源的重要成员,在生物防治中作为一类有效的自然调控因子,对害虫的种群抑制起着重要作用,同时也是农业生产中可以利用的重要天敌资源(李晓明和缪勇,2011;胡长效等,2020)。叉角厉蝽[Eocanthecona furcellata (Wolff)]隶属于半翅目(Hemiptera)蝽科(Pentatomidae)益蝽亚科(Asopinae)昆虫,其若虫和成虫对多数鳞翅目害虫幼虫、部分鞘翅目害虫幼虫和半翅目害虫等多种农业害虫具有较强的捕食能力(陈雪梅等,2021),是一种重要的捕食性天敌,因而受到国内外学者广泛关注。作为刺吸式捕食性天敌,叉角厉蝽不仅对多数农业害虫有较强的捕食能力,还拥有繁殖力强、产卵量大、害虫不会产生抗性、可操作性强、对环境无害等优点,具有广阔的生物防治应用前景。而昆虫生长发育特征受多种环境因素的交互影响,明确叉角厉蝽生长发育特征和繁殖力特性是开发叉角厉蝽生防潜力的前提条件,而研究叉角厉蝽不同虫态形态学特征则可为田间快速鉴定及田间统计提供理论支持。【前人研究进展】叉角厉蝽的世代发育分为卵、若虫、成虫3个时期,若虫有5个龄期,初孵若虫具有聚集的习性,成虫羽化7~10 d可观察到交尾(朱涤芳,1990);林长春等(1998)林间饲养叉角厉蝽雌雄成虫的体长分别为14.66~15.96和11.54~13.22 mm,体宽分别为6.32~6.51和4.89~5.70 mm,第3代无法完成发育;Lenin和Rajan(2016)用米蛾喂养叉角厉蝽,结果表明若虫总历期约16.00±0.64 d,雌雄成虫寿命分别为36.00±1.90和32.00±0.19 d;李慎磊等(2020)在对叉角厉蝽饲养技术的研究中发现,室温25~32 ℃、空气相对湿度60%~80%、光照周期14 L∶10 D的条件下,叉角厉蝽完成一个世代需要41.40 d,羽化为成虫发育成熟并交配产卵平均需要14.20 d,5龄若虫发育历期最长,但存活率相对较低。叉角厉蝽对多数鳞翅目、鞘翅目及半翅目害虫具有较强的捕食能力,其捕食量据其自身虫龄和密度、猎物大小和猎物密度而定,捕食量随自身虫龄和密度、猎物密度增大而增大(朱涤芳,1990;陈然等,2015;李文华等,2015)。据已有文献报道,叉角厉蝽捕食行为主要发生在若虫和成虫阶段,其中1龄若虫不捕食,仅靠吸食植物汁液即可完成发育,2龄开始捕食,随龄期的增长植食性逐渐减弱,捕食性逐渐增强(林长春等,1998;姚明勇等,2019)。2~3龄若虫每天可捕食1头米蛾(Corcyra cephalonica)幼虫,4~5龄若虫每天可捕食1~2头米蛾幼虫,成虫每天可捕食2~3头米蛾幼虫或1~2头大蜡螟(Galleria mellonella)幼虫(朱涤芳,1990)、3.75头绿额翠尺蛾(Pelagodes proquadraria)幼虫(谢钦铭等,2001)。草地贪夜蛾(Spodoptera frugiperda)是近年我国重要的入侵性害虫(宋洁蕾等,2019),范悦莉等(2019)研究表明,叉角厉蝽若虫对草地贪夜蛾3龄幼虫日最大捕食量、瞬时攻击率和处理时间分别为50.25头、0.6702 d和0.0199 d,其对草地贪夜蛾有较好的控害作用;唐敏等(2019)研究表明,叉角厉蝽对草地贪夜蛾幼虫具有较强的捕食能力,5龄若虫最强、雌成虫和雄成虫次之;李慎磊等(2020)田间试验结果表明,在玉米田草地贪夜蛾虫量约为20头/百株时,按照0.24头/m2释放叉角厉蝽5龄若虫,7 d后草地贪夜蛾虫口减退率达72.41%,且玉米拔节期释放叉角厉蝽效果好于喇叭口期和抽穗期。在饲养建立种群方面,张敏玲和卢传权(1996)用菜青虫和斜纹夜蛾饲养叉角厉蝽也可完成发育,且较喜食斜纹夜蛾,但不足的是不易得到大量的活体菜青虫和斜纹夜蛾以供叉角厉蝽繁殖;何旭诺等(2013)用甜菜夜蛾(S. exigua)幼虫和黄粉虫(Tenebrio molitor)幼虫饲喂叉角厉蝽,结果显示用甜菜夜蛾饲喂的叉角厉蝽若虫历期较短,而用黄粉虫幼虫饲喂的叉角厉蝽若虫存活率较高;Tuan等(2016)用小菜蛾(Pieris rapae)和斜纹夜蛾(S. litura)饲养叉角厉蝽,结果表明,小菜蛾饲养下叉角厉蝽的繁殖率和增长率显著高于用斜纹夜蛾饲养的叉角厉蝽;宫靖垚(2018)用研制的人工饲料饲喂叉角厉蝽能完成世代发育,但其发育历期、存活率、成虫体重、产卵量等指标大多不及用黄粉虫饲养的结果。微观上,Kumar等(2002)利用扫描电镜观察了叉角厉蝽卵粒的结构;高平等(2021)及Gao等(2021)利用扫描电镜观察了叉角厉蝽唾液腺的形态和超微结构,并对其唾液腺进行了转录组及差异分析,从基因层面研究唾液腺的特征与功能,从转录组水平预测唾液腺的潜在分泌蛋白;Zhao等(2021)观察了叉角厉蝽5龄若虫和雌雄成虫的触角超微结构。【本研究切入点】目前,国内外对叉角厉蝽生长发育方面的研究已有报道,但对各虫态形态特征系统性的描述仅见林长春等(1998)在福建省野外試验林中的调查研究,而对室内优化饲养条件下的叉角厉蝽生长发育、繁殖情况及各龄若虫的形态特征未见报道。【拟解决的关键问题】采用单头饲养法和雌雄虫配对共培养的方法,以云南元江采集并在室内扩繁6代以上的叉角厉蝽为研究对象,探明叉角厉蝽在室内恒定条件下的生长发育、繁殖特征、形态学特征以及是否存在孤雌生殖现象,并用显微照相系统观察叉角厉蝽不同虫态的形态学特征,为叉角厉蝽人工规模化养殖、释放技术的发展及其田间应用提供科学依据。
1 材料与方法
1. 1 试验材料
供试叉角厉蝽于2018年采集自云南省玉溪市元江县(东经101°32′,北纬23°30′),由最初25头大小和龄期不等的虫源带回实验室用养虫笼(50 cm×50 cm×40 cm)饲养[温度(27±1)℃,相对湿度60%~80%,光周期14 L∶10 D],每日饲喂黄粉虫蛹,此后每年7~9月补充1次虫源,建立实验室种群并连续饲养6代以上。
1. 2 试验方法
1. 2. 1 叉角厉蝽各虫态的生长发育历期及繁殖力测定 在叉角厉蝽饲养笼中收集当日产的卵块(共9块),在显微镜下观察记录每卵块的卵粒数,然后将卵块置于干燥消毒的培养皿中,记录收集日期。将培养皿置于恒温培养箱内,每天观察并记录卵的孵化情况。卵孵化后,将1龄若虫置于消毒的培养皿中单独饲养,同时放入潮湿的棉球和黄粉虫蛹以提供所需水分和食物。每天观察记录若虫蜕皮及死亡情况并更换食物和棉球,直至成虫。
将当日羽化的叉角厉蝽雌、雄成虫配对置于塑料养虫盒(12.5 cm×8.0 cm×3.5 cm)中饲养,并提供潮湿棉球和新鲜黄粉虫蛹供其取食,每天更换。待雌成虫产卵后,记录成虫产卵日期、卵块数、卵粒数和成虫死亡时间。若雄虫死亡,则雌成虫继续饲养直至死亡,共设20个重复。
1. 2. 2 雌成虫是否存在孤雌生殖测定 根据日常饲养经验,有目的地挑选50头可能为雌性的叉角厉蝽5龄若虫于塑料养虫盒中进行单独饲养,而后将羽化的35头雌成虫继续单独饲养并每天观察记录其产卵日期、卵块数、卵粒数和死亡时间,将所产卵块置于培养皿(φ=60 mm)中饲养并观察其是否孵化。
1. 2. 3 若虫大小及形态特征观察 叉角厉蝽1龄若虫体长、体宽采用EZ-MET软件进行测量,2~5龄若虫在其蜕皮当天便用游标卡尺测量体长、体宽,用电子天平测量体重,若虫每一龄期和雌雄成虫各测量20头。在日产Olympus双目立体解剖镜下检视和观察各虫态的形态特征,重要的鉴别特征与拍照借助日产基恩士(KEY-CNCE VHX-5000)超景深显微照相系统完成。
1. 3 统计分析
试验数据采用Excel 2016进行整理,并用DPS v9.50数据处理系统进行统计分析。卵孵化率及若虫存活率经反正弦转换后进行方差分析,每个卵块作为1个重复;对各龄若虫的发育历期和存活率进行单因素方差分析(One-way ANOVA),并用Tukey法进行多重比较分析;卵孵化率、若虫存活率和雌雄成虫寿命采用独立样本t-检验进行差异显著性分析;使用GraphPad Prism v9.2绘图。
2 结果与分析
2. 1 叉角厉蝽的发育历期
在本试验条件下,叉角厉蝽从卵到成虫的平均发育历期为29.00±1.41 d,最长31 d,最短25 d。叉角厉蝽的卵历期为8.00±0.21 d,若虫期19.76±0.12 d。其中1龄与3龄、1龄与4龄间历期差异不显著(P>0.05,下同),其他各龄期间历期差异显著(P<0.05,下同),1~5龄若虫发育历期为3.24~5.38 d(表1)。
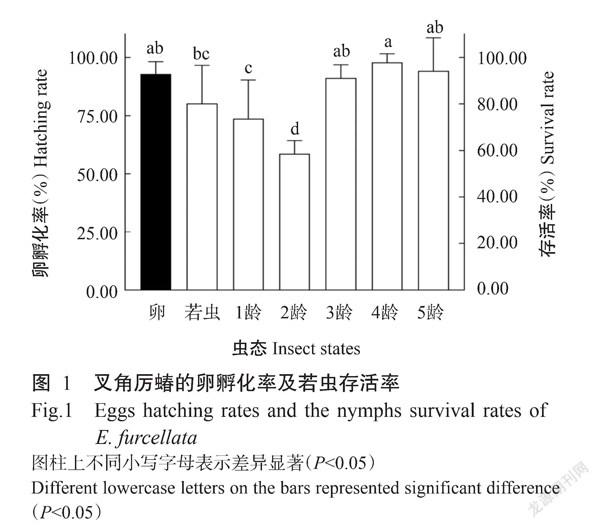
2. 2 叉角厉蝽卵孵化率及若虫存活率
叉角厉蝽的卵孵化率达92.91%,高于若虫存活率(80.21%),显著高于1龄和2龄若虫存活率。以4龄若虫的存活率最高,为97.82%,显著高于1龄和2龄若虫存活率,其他各龄若虫的存活率间无显著差异(图1)。
2. 3 叉角厉蝽成虫寿命及繁殖力
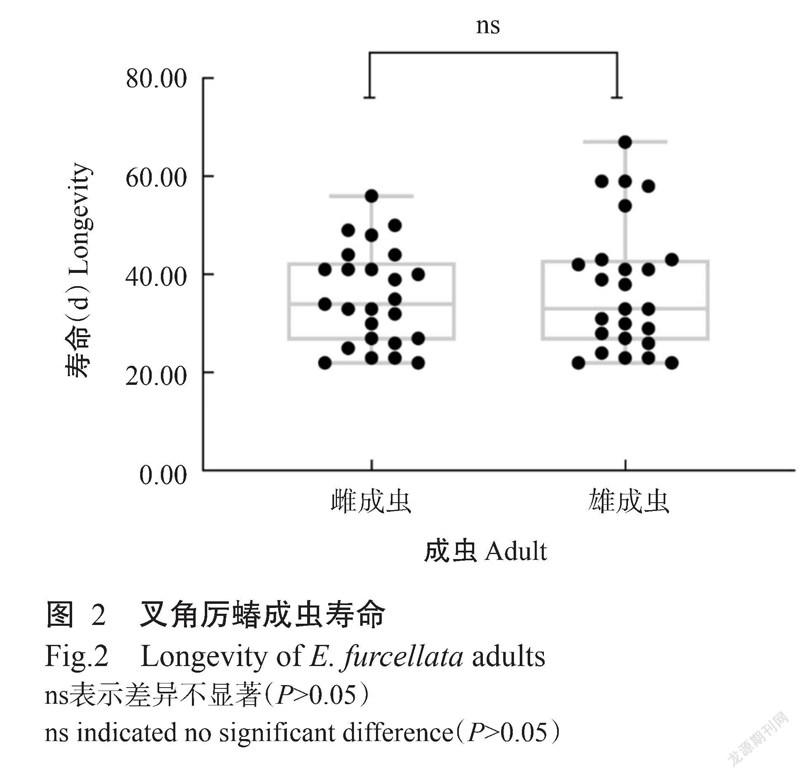
2. 3. 1 雌、雄成虫寿命 叉角厉蝽雌、雄成虫寿命间无显著差异(df=49,P=0.5475),雌、雄成虫的平均寿命分别为35.40±1.96和37.40±2.65 d(图2)。雌成虫寿命最长为56 d、最短22 d;雄成虫寿命最长为67 d,最短22 d。
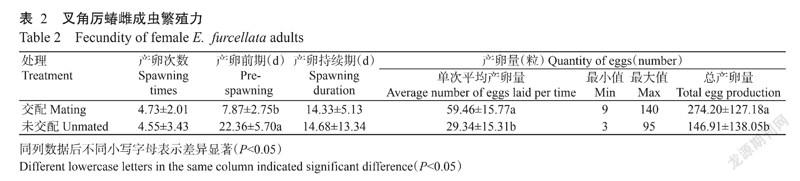
2. 3. 2 雌成虫繁殖力 叉角厉蝽雌成虫羽化后与同日龄雄成虫配对,最早第2 d观察到交尾,6日龄开始产卵,单次产卵量9~140粒,从配对后1个月期间每头雌成虫总产卵量70~585粒。雌成虫与雄成虫交配单次平均产卵量为59.46±15.77粒,总产卵量为274.20±127.18粒,一生产卵次数为4.73±2.01次,最多9次,最少2次;产卵前期为7.87±2.75 d,产卵期持续14.33±5.13 d(表2)。观察发现,雄成虫死亡一周后雌成虫仍继续产卵,但无法成功孵化,为无效卵。
未交配雌成虫最早13日龄开始产卵,单次产卵量3~95粒,从羽化后2个月期间每头雌成虫总产卵量13~444粒。产卵次数为4.55±3.43次,最多12次,最少1次。产卵前期为22.36±5.70 d,显著长于交配雌成虫;产卵持续期为14.68±13.34 d,每雌成虫单次平均产卵量为29.34±15.31粒,每雌成虫一生总产卵量为146.91±138.05粒,单次平均产卵量和总产卵量均显著少于交配雌成虫(表2)。
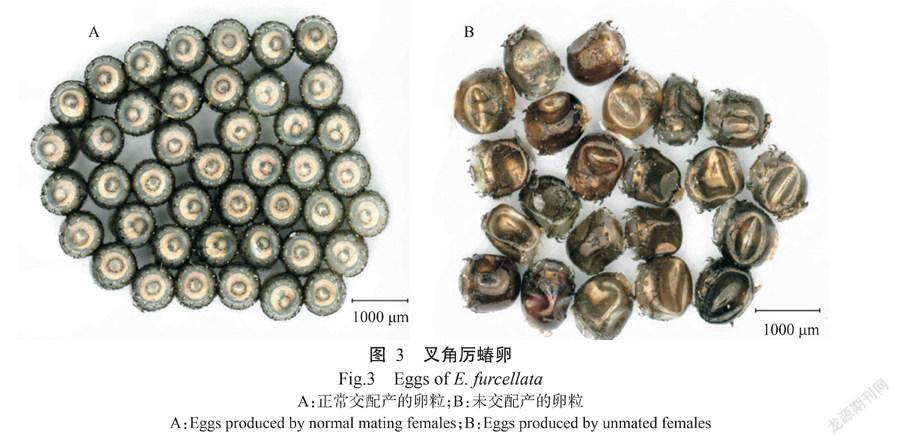
经2个月的观察发现,叉角厉蝽未交配雌成虫单头在室内恒定条件下饲养会出现产卵现象,所产的卵虽有色泽变化,但后期卵壳变瘪,均为无效卵,即无孵化能力(图3-B)。
2. 4 叉角厉蝽各虫态体型大小及形态特征
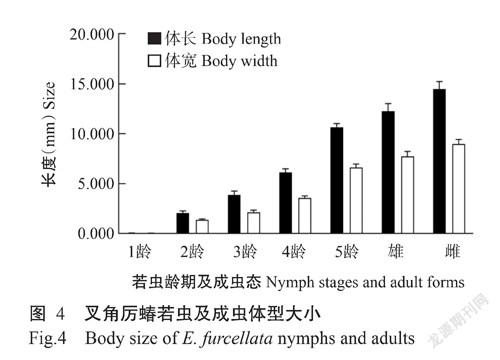
2. 4. 1 叉角厉蝽各虫态体型大小 叉角厉蝽若虫体型随着龄期的增长逐渐增大,1~5龄若虫体长分别为0.050±0.001、2.070±0.040、3.880±0.086、6.120±0.080和10.670±0.080 mm;雌雄成虫体长分别为 14.480±0.160和12.240±0.170 mm。1~5齡若虫体宽分别为0.030±0.001、1.370±0.020、2.140±0.050、 3.570±0.050和6.509±0.080 mm;雌雄成虫体宽分别为 8.960±0.100和7.720±0.110 mm(图4)。1龄和2龄若虫体重均值分别为0.28和0.65 μg,3~5龄若虫体重分别为5.45±0.42、18.10±1.18和111.95±6.62 mg,雌雄成虫体重分别为175.85±4.98和152.75±2.51 mg。
2. 4. 2 叉角厉蝽各虫态形态特征 卵:矮杯形或圆筒形,具卵盖,边缘有10~12根刺状精孔突;多数单层排列成块状,偶有双层排列;初产卵粒银灰色或银白色,后逐渐变为灰黑色至褐色,有金属光泽(图3-A)。
1龄若虫:扁圆形;初孵若虫红色,渐成红黑相间;触角4节,第2和第3节末端有明显白色环;喙暗红色,接近体长;胸部背面黑色,无刻点;足黑色,越往端部颜色越浅;腹部背面红色,有3个黑色横斑;无翅芽(图5-A和图6-A)。
2龄若虫:扁椭圆形;体色变深;头部前段扁平突出;触角4节,节间环变为红色;喙黑色,接近体长;胸部背面黑色,无刻点;足完全为黑色;腹部红色,有3个黑色横斑;无翅芽(图5-B和图6-B)。
3龄若虫:梨形;体色红(或橙红)黑相间;触角4节,第2和第3节末端红色环明显;喙黑色,为体长的一半;胸部背面黑色,出现刻点;足黑色,前足胫节略微膨大;腹部背面红色,有4个黑色横斑;无翅芽(图5-C和图6-C)。
4龄若虫:梨形;体色红(或橙红)黑相间;头部前段扁平突出明显;触角4节,节间环红色;喙黑色,为体长的一半;胸部第1节红色或红橙色,其余为黑色,刻点明显;腹部背面红色或橙红色,依次有2条黑色横线,4个黑色横斑;出现翅芽,并延伸至胸部末端(图5-D和图6-D)。
5龄若虫:梨形;体色红(或橙红)黑相间;喙暗红色,短于体长一半;胸部第1节中间为黑色,两侧为红色或红橙色,其余为黑色,刻点明显;腹部背面红色或红橙色,依次有1条黑色横线,4个黑色横斑;足黑色,中足和后足胫节中部出现白斑;翅芽延伸至腹部第3节(图5-E和图6-E)。
成虫:体色黄褐与黑褐混杂相间,密布刻点;头黑色,触角5节,第1节短,2~5节基部为黄色,黄色范围逐渐增大;喙黄褐色,最后一节黑色,短于体长的一半;前胸背板侧角呈剑叉状突出,黑色;小盾片基部黑褐色,基角具有黄斑,端部盾圓;雄虫腹部近三角形,雌虫腹部卵圆形;中足和后足的腿节基部浅黄褐色,端部黑色,胫节基部与端部黑色,中间浅黄褐色(图5-F和图5-G,图6-F和图6-G)。
3 讨论
叉角厉蝽是一种重要的捕食性天敌,在农业生产上具有较高的应用潜能,但实际应用并不广泛,研究并掌握叉角厉蝽在室内饲养的发育历期、繁殖力及其形态特征,对叉角厉蝽的人工规模化养殖和释放技术的发展具有重要意义。本研究结果表明,在(27±1)℃,相对湿度 60%~80%,光周期14 L∶10 D的恒定室内条件下,叉角厉蝽卵孵化率为92.91%,若虫存活率为80.21%,其中以4龄若虫的存活率最高,5龄若虫存活率其次,2龄若虫存活率最低。林长春等(1998)研究结果显示,叉角厉蝽5龄若虫的存活率最低,其研究是在南亚热带气候的野外试验林中进行,温度和湿度等环境条件并不稳定,而本研究是在室内恒定的环境条件下进行,造成2项研究结果的差异可能与试验地的气候条件及种群自身遗传等因素相关。温度是影响昆虫生长发育的重要因素。张敏玲和卢传权(1996)对叉角厉蝽饲养条件的研究表明,室温低于19 ℃条件下长时间饲养的叉角厉蝽成虫既不产卵也不交配,不低于25 ℃条件下叉角厉蝽才能正常生长、发育和繁殖。已报道文献中多数试验饲养温度为(27±1)℃,在此温度条件下,姚明勇等(2019,2020)探究了光周期和性比对叉角厉蝽生长发育、繁殖及成虫寿命的影响;姚琼等(2021)探究了叉角厉蝽章鱼胺受体的基因表达及化学农药对其表达的影响;以及叉角厉蝽对草地贪夜蛾(范悦莉等,2019;唐敏等,2019)、粘虫(廖贤斌等,2020)的捕食功能和对斜纹夜蛾的搜索效率的影响(陈雪梅等,2021)。因此,本研究设置温度为(27±1)℃,得到1~5龄若虫的平均发育历期为3~6 d,若虫历期为19.76 d,优于朱涤芳(1990)在温度为25 ℃条件下饲养的21.60 d。
食物也极大地影响着昆虫的生长发育。何旭诺等(2013)应用桔小实蝇、黄粉虫、斜纹夜蛾和甜菜夜蛾4种活体昆虫的幼虫饲养叉角厉蝽,结果表明用黄粉虫幼虫饲养的叉角厉蝽若虫存活率最高,产卵前期也最短。宫靖垚(2018)研究结果显示,人工饲料饲养的叉角厉蝽若虫发育历期、存活率、成虫体重和产卵量等指标大多不及用黄粉虫饲养的叉角厉蝽若虫。前人曾尝试利用菜青虫饲养叉角厉蝽(张敏玲和卢传权,1996),但菜青虫在饲养过程中易受病毒感染,饲养技术不成熟,难以获得充足的活体菜青虫供叉角厉蝽扩繁,故相比于其他饲料而言,黄粉虫的饲养技术更成熟,并已发展成为一种产业化养殖,可得到充足的虫源供叉角厉蝽扩繁。黄粉虫的幼虫、蛹、成虫均含有丰富的蛋白质和氨基酸,是常用的饲料昆虫和食品蛋白添加剂(蒋素容等,2012;解晓菲等,2020),其幼虫在发育后期具有自相残杀的习性(高红莉等,2006),且活动能力较蛹更强,在每次蜕皮后,表皮逐渐硬化(刘守柱等,2009),其蛹则脆弱且肉质多汁,活动能力弱,故黄粉虫蛹较幼虫而言更有利于叉角厉蝽的刺吸式口器取食。姚明勇等(2020)以斜纹夜蛾幼虫为食物,光周期14 L∶10 D条件下叉角厉蝽若虫的存活率为64.3%,本研究以黄粉虫蛹为食物,在相同的光周期条件下叉角厉蝽若虫的存活率为80.21%,可见黄粉虫蛹饲养更有利于该虫的种群扩繁。
孤雌生殖是一种在特殊情况下对遗传资源应急保护的生殖方式(徐立军等,2016),并且孤雌生殖现象广泛存在于昆虫界中,很多研究均表明孤雌生殖可适应恶劣的环境,保证种群得以繁衍和扩大种群分布(王成业,2011;孙玺文等,2018)。在农业生产上,产雌孤雌生殖的松毛赤眼蜂可以单性生殖产生可育的雌性后代,并且具有良好的防治效能和较强的野外定殖能力(李媛媛,2019)。研究发现,产雌孤雌生殖品系的松毛虫赤眼蜂与正常两性生殖的松毛虫赤眼蜂对玉米螟的防治效果及对玉米的增产效果无显著差异,可产生相同的防治效应(张海燕,2009),这为具有孤雌生殖能力的天敌昆虫在农业生产上应用提供了依据。本研究发现叉角厉蝽未交配雌成虫有产卵现象,故进一步探究其是否存在孤雌生殖现象,而经观察,未交配的叉角厉蝽雌成虫所产卵无法正常孵化,无孤雌生殖现象。王小芬(2012)在室内改变南方小花蝽生存的环境温度、湿度以及食物和性比,以探究在胁迫条件下南方小花蝽是否存在孤雌生殖现象,并结合微卫星标记技术验证南方小花蝽田间自然种群是否存在孤雌生殖现象,结果发现南方小花蝽无孤雌生殖现象。南方小花蝽与叉角厉蝽同为半翅目捕食性天敌,其繁殖特性具有相似性,目前只有少量文献报道蝽类昆虫存在孤雌生殖现象,而且昆虫孤雌生殖现象是昆虫长期生存斗争的适应性结果(王成业,2011)。林长春等(1998)报道在林间连续饲养叉角厉蝽3代,第3代叉角厉蝽若虫不能完成发育,后期卵也不能正常孵化,故本研究推测自然条件下叉角厉蝽也无存在孤雌生殖现象的可能。此外,本研究还分析比较了未交配雌成虫与交配雌成虫间产卵前期、单次产卵量及一生总产卵量的差异,其中产卵前期未交配雌成虫显著长于交配雌成虫,总产卵量显著少于交配雌成虫,说明在一般情况下,两性生殖中雌雄虫交配可刺激雌成虫产卵和加速产卵速率,从而导致总产卵量的增加(张美翠,2015)。
全面了解叉角厉蝽的生长发育、繁殖及形态特征等有利于其更好地应用到害虫生物防治中。本研究供试虫体均是在室内长期饲养的稳定种群,而野外定殖的虫体在生长环境上有所差别,这对其发育进程有一定影响,因此,在后期研究中可通过比较野外定殖群体与室内群体的发育历期、繁殖力、存活率及寿命等生物学指标以组建生命表来评估和掌握叉角厉蝽室内群体与野外群体的发育差异,及时向室内补充野外群体,防止室内叉角厉蝽种群退化(朱艳娟等,2021)。此外,探究叉角厉蝽室内继代饲养种群是否退化及退化规律可指导改善室内饲养环境,提升室内饲养品质,推进人工规模化养殖和释放技术的发展,从而将其有效应用于农业生产,为绿色农业服务。
4 结论
明确了叉角厉蝽的发育历期和繁殖能力,证实叉角厉蝽无孤雌生殖现象。农业生产中可以喙的颜色、长度及翅芽发育作为叉角厉蝽主要形态鉴别特征。
参考文献:
陈然,梁广文,张拯研,曾嵘,冼继东. 2015. 叉角厉蝽对斜纹夜蛾的捕食功能反应[J]. 环境昆虫学报,37(2):401-406. [Chen R,Liang G W,Zhang Z Y,Zeng R,Xian J D. 2015. The functional response of Cahtheconidea furcellata (Hemiptera:Asopinae) to Spodoptera litura (Lepidoptera:Noctuidae)[J]. Journal of Environmental Entomology,37(2):401-406.] doi:10.3969/j.issn.1674-0858. 2015.02.26.
陈雪梅,谷星慧,冼繼东,张立猛,范悦莉,黄智华,周文兵,赵进龙,张翠萍. 2021. 叉角厉蝽对烟草上斜纹夜蛾搜索效率影响因子的研究[J]. 环境昆虫学报,43(1):224-232. [Chen X M,Gu X H,Xian J D,Zhang L M,Fan Y L,Huang Z H,Zhou W B,Zhao J L,Zhang C P. 2021. A study on influence factors of search efficiency of Eocanthecona furcellata (Hemiptera) on Spodoptera litura (Le-pidoptera:Noctuidae) in tobacco[J]. Journal of Environmental Entomology,43(1):224-232.] doi:10.3969/j.issn. 1674-0858.2021.01.23.
范悦莉,谷星慧,冼继东,陈雪梅,陆永跃,张立猛,周文兵. 2019. 叉角厉蝽对草地贪夜蛾的捕食功能反应[J]. 环境昆虫学报,41(6):1175-1180. [Fan Y L,Gu X H,Xian J D,Chen X M,Lu Y Y,Zhang L M,Zhou W B. 2019. Functional response of Eocanthecona furcellate (Hemiptera:Pentatomidae) to Spodoptera frugiperda (Lepidoptera:Noctuidae)[J]. Journal of Environmental Entomo-logy,41(6):1175-1180.] doi:10.3969/j.issn.1674-0858. 2019.06.06.
高红莉,周文宗,张硌,李洪涛. 2006. 饲料种类和饲养密度对黄粉虫幼虫生长发育的影响[J]. 生态学报,26(10):3258-3264. [Gao H L,Zhou W Z,Zhang G,Li H T. 2006. Effect of different fodders and breeding densities on the larva growth and development of Tenebrio molitor L.[J]. Acta Ecologica Sinica,26(10):3258-3264.] doi:10.3321/j.issn:1000-0933.2006.10.014.
高平,廖贤斌,李丽芳,兰明先,赵航,陈斌,吴国星,高熹. 2021. 叉角厉蝽唾液腺转录组及差异分析[J]. 云南农业大学学报(自然科学),36(1):29-38. [Gao P,Liao X B,Li L F,Lan M X,Zhao H,Chen B,Wu G X,Gao X. 2021. Transcriptome and difference analysis of Eocanthecona furcellata (Wolff)[J]. Journal of Yunnan Agricultural University (Natural Science Edition),36(1):29-38.] doi:10.12101/j.issn.1004-390X(n).201911011.
宫靖垚. 2018. 叉角厉蝽人工饲料研制及其捕食效能评价[D]. 广州:华南农业大学. [Gong J Y. 2018. The artificial diet for Eocanthecona furcellate (Wolff) and its pre-datory efficiency[D]. Guangzhou:South China Agricultural University.]
何旭诺,冼继东,陈然,张拯研,曾嵘. 2013. 4种昆虫饲料对叉角厉蝽生长发育和繁殖的影响[J]. 环境昆虫学报,35(6):799-803. [He X N,Xian J D,Chen R,Zhang Z Y,Zeng R. 2013. Effects of four insect feed on development and reproduction of Cantheconidea furcellata (Hemiptera:Asopinae)[J]. Journal of Environmental Entomology,35(6):799-803.] doi:10.3969/j.issn.1674-0858. 2013.06.16.
胡长效,强承魁,王胜永. 2020. 微小花蝽对梨瘿蚊的室内捕食作用[J]. 江苏农业学报,36(1):57-62. [Hu C X,Qiang C K,Wang S Y. 2020. Predation of Orius minutes on Dasumeira pyri in the laboratory[J]. Jiangsu Journal of Agricultural Sciences,36(1):57-62.] doi:10.3969/j.issn.1000-4440.2020.01.008.
蒋素容,蔡艳,周宇爝. 2012. 不同飼料对黄粉虫幼虫水分及粗蛋白和粗脂肪含量的影响[J]. 四川农业大学学报,30(2):243-247. [Jiang S R,Cai Y,Zhou Y J. 2012. Effects of different feedstuffs on the content of water,crude protein and fat in Tenebrio molitor L.[J]. Journal of Sichuan Agricultural University,30(2):243-247.] doi:10.3969/j.issn.1000-2650.2012.02.023.
李慎磊,简国新,邓伟林,谷小红,刁碧群,林小军,管云. 2020. 叉角厉蝽人工饲养技术及其防治草地贪夜蛾的初步研究[J]. 中国植保导刊,40(3):56-60. [Li S L,Jian G X,Deng W L,Gu X H,Diao B Q,Lin X J,Guan Y. 2020. A preliminary study of Eocanthecona furcellata (Wolff) artificial breeding technology and its control of Spodoptera frugiperda (Lepidoptera:Noctuidae)[J]. China Plant Protection,40(3):56-60.] doi:10.3969/j.issn.1672-6820.2020.03.011.
李文华,贾彩娟,陈惠平,付浪,温健,陈科伟. 2015. 叉角厉蝽对黄野螟幼虫的捕食功能反应[J]. 环境昆虫学报,37(4):843-848. [Li W H,Jia C J,Chen H P,Fu L,Wen J,Chen K W. 2015. Functional response of Eocanthecona furcellate(Wolff) to the larvae of Heortia vitessoides (Moore)[J]. Journal of Environmental Entomology,37(4):843-848.] doi:10.3969/j.issn.1674-0858.2015.04.21.
李晓明,缪勇. 2011. 半翅目捕食性天敌昆虫评价研究[J]. 安徽农业科学,39(17):10297-10298. [Li X M,Miao Y. 2011. Study on evaluation of Hemiptera predatory insects[J]. Journal of Anhui Agricultural Sciences,39(17):10297-10298.] doi:10.13989/j.cnki.0517-6611.2011.17.147.
李媛媛. 2019. 松毛虫赤眼蜂产雌孤雌生殖表型及体内Wolbachia含量影响因素分析[D]. 辽宁:沈阳农业大学. [Li Y Y. 2019. Analysis of the factors on thelytokous-phenotype and Wolbachia titer in Trichogramma dendrolimi Matsumura(Hymenoptera:Trichogrammatidae)[D]. Liaoning:Shenyang Agricultural University.] doi:10.27327/ d.cnki.gshnu.2019.000346.
廖贤斌,高平,赵航,谢永辉,陈雅琼,王志江,陈斌,吴国星,高熹. 2020. 叉角厉蝽成虫对粘虫幼虫的捕食功能反应[J]. 南方农业学报,51(8):1992-1997. [Liao X B,Gao P,Zhao H,Xie Y H,Chen Y Q,Wang Z J,Chen B,Wu G X,Gao X. 2020. Predatory function response of adult of Eocanthecona furcellata(Wolff) to the larvae of Mythimna seperata (Walker)[J]. Journal of Southern Agriculture,51(8):1992-1997.] doi:10.3969/j.issn.2095-1191.2020.08.026.
林长春,王浩杰,任华东,洪长福. 1998. 叉角厉蝽生物学特性研究[J]. 林业科学研究,11(1):92-96. [Lin C C,Wang H J,Ren H D,Hong C F. 1998. Studies on the biological characteristics of Cantheconidea furcellata (Wolff) (Hemiptera Pentatomidae,Asopinae)[J]. Forest Research,11(1):92-96.] doi:10.13275/j.cnki.lykxyj.1998.01.015.
刘守柱,薛超彬,罗万春. 2009. 黄粉虫幼虫体壁硬化过程中酚氧化酶活性的变化[J]. 昆虫学报,52(9):941-945. [Liu S Z,Xue C B,Luo W C. 2009. Phenoloxidase activi-ty during cuticle sclerotization in larvae of Tenebrio molitor(Coleoptera:Tenebrionidae)[J]. Acta Entomologica Sinica,52(9):941-945.] doi:10.16380/j.kcxb.2009.09. 013.
宋潔蕾,李艳丽,李亚红,黄碧芬,康云昌,马庭矗,张澍雷,桂富荣. 2019. 不同杀虫剂对草地贪夜蛾的室内毒杀效果 及毒力测定[J]. 南方农业学报,50(7):1489-1495. [Song J L,Li Y L,Li Y H,Huang B F,Kang Y C,Ma T C,Zhang S L,Gui F R. 2019. Measurements of indoor toxic effects and virulence of different insecticides against Spodoptera frugiperda[J]. Journal of Southern Agriculture,50(7):1489-1495.] doi:10.3969/j.issn.2095-1191. 2019.07.12.
孙玺文,张廷伟,刘长仲. 2018. 配对时间对二斑叶螨种群繁殖的影响[J]. 甘肃农业大学学报,53(2):69-73. [Sun X W,Zhang T W,Liu C Z. 2018. Effects of paring time on the population reproduction of Tetranychus urticae[J]. Journal of Gansu Agricultural University,53(2):69-73.] doi:10.3969/j.issn.1003-4315.2018.02.011.
唐敏,邝昭琅,李子园,陆永跃,陈科伟,刘光华. 2019. 叉角厉蝽对草地贪夜蛾幼虫的捕食功能反应[J]. 环境昆虫学报,41(5):979-985. [Tang M,Kuang Z L,Li Z Y,Lu Y Y,Chen K W,Liu G H. 2019. Functional response of Eocanthecona furcellate(Wolff) to the larvae of Spodoptera frugiperda (J. E. Smith)[J]. Journal of Environmental Entomology,41(5):979-985.] doi:10.3969/j.issn.1674- 0858.2019.05.8.
王成业. 2011. 昆虫孤雌生殖起源的遗传机制和进化意义[J]. 动物学研究,32(6):689-695. [Wang C Y. 2011. Genetic mechanism and evolutionary significance of the origin of parthenogenetic insects[J]. Zoological Research,32(6):689-695.] doi:10.3724/SP.J.1141.2011.06689.
王小芬. 2012. 南方小花蝽孤雌生殖的微卫星标记验证[D]. 武汉:华中农业大学. [Wang X F. 2012. Parthenogenesis of Orius similis tesred by using microsatellites[D]. Wuhan:Huazhong Agricultural University.]
谢钦铭,梁广文,罗诗,朱君林. 2001. 叉角厉蝽对绿额翠尺蛾幼虫的捕食作用的初步研究[J]. 江西科学,19(1):21-23. [Xie Q M,Liang G W,Luo S,Zhu J L. 2001. Observation of the predacious function of a bug Cantheconidae furcellata on litchi looper Thalassodes proquadria[J]. Jianxi Science,19(1):21-23.] doi:10.13990/j.issn 1001-3679.2001.01.00.
解晓菲,黄勇,夏鹏亮,任晓红,王瑞. 2020. 探索以黄粉虫蛹为食物繁育蠋蝽的可行性[J]. 湖北农业科学,59(14):85-87. [Xie X F,Huang Y,Xia P L,Ren X H,Wang R. 2020. Explore the feasibility of breeding Arma chinensis with Tenebrio molitor as food[J]. Hubei Agricultural Scien-ces,59(14):85-87.] doi:10.14088/j.cnki.issn0439-8114. 2020.14.015.
徐立军,嵇保中,刘曙雯,丁芳,王亚召. 2016. 白蚁的孤雌生殖[J]. 中国森林病虫,35(6):32-36. [Xu L J,Ji B Z,Liu S W,Ding F,Wang Y Z. 2016. Termites parthenogene-sis[J]. Forest Pest and Disease,35(6):32-36.] doi:10. 3969/j.issn.1671-0886.2016.06.008.
姚明勇,王岚,周吕,黄敏,陈文龙. 2019. 性比对叉角厉蝽成虫寿命和繁殖力的影响[J]. 山地农业生物学报,38(3):78-81. [Yao M Y,Wang L,Zhou L,Huang M,Chen W L. 2019. Effects of different sex ratios on longevity and fecundity of adults Cantheconidea furcellata (Hemiptera:Asopinae)[J]. Journal of Mountain Agriculture and Bio-logy,38(3):78-81.] doi:10.15958/j.cnki.sdnyswxb.2019. 03.014.
姚明勇,周吕,王岚,陈文龙. 2020. 光周期对叉角厉蝽生长发育及繁殖的影响[J]. 西南师范大学学报(自然科学版),45(3):109-114. [Yao M Y,Zhou L,Wang L,Chen W L. 2020. Effects of different photoperiods on development and reproduction of Eocanthecona furcellata[J]. Journal of Southwest China Normal University(Natural Science Edition),45(3):109-114.] doi:10.13718/j.cnki.xsxb.2020. 03.017.
姚琼,全林发,徐淑,董易之,李文景,池艳艳,陈炳旭. 2021. 叉角厉蝽2个章鱼胺受体的基因克隆及化学农药对其表达的影响[J]. 生物技术通报,37(10):152-161. [Yao Q,Quan L F,Xu S,Dong Y Z,Li W J,Chi Y Y,Chen B X. 2021. Gene cloning of 2 octopamine receptors from Eocanthecona furcellata and effects of chemical pesticide on its expression[J]. Biotechnology Bulletin,37(10):152-161.] doi:10.13560/j.cnki.biotech.bull.1985.2021-0064.
张海燕. 2009. 赤眼蜂种间Wolbachia转染与生防潜力研究[D]. 辽宁:沈阳农业大学. [Zhang H Y. 2009. Wolbachia transfection and biological control capability of Trichogramma spp.[D]. Liaoning:Shenyang Agricultural University.]
张美翠. 2015. 扶桑綿粉蚧不同生殖方式的比较研究[D]. 金华:浙江师范大学. [Zhang M C. 2015. Comparative study of reproductive mode of Phenacoccus solenopsis Tinsley[D]. Jinhua:Zhejiang Normal University.]
张敏玲,卢传权. 1996. 叉角厉蝽的饲养[J]. 昆虫天敌,18(1):74-77. [Zhang M L,Lu C Q. 1996. Rearing of Eocanthecona furcellata[J]. Natural Enemies of Insects,18(1):74-77.]
朱涤芳. 1990. 叉角厉蝽生物学特性研究[J]. 昆虫天敌,12(2):71-74. [Zhu D F. 1990. Studies on the biological characteristics of Cantheconidea furcellata (Wolff) (Hemiptera Pentatomidae,Asopinae)[J]. Natural Enemies of Insects,12(2):71-74.]
朱艳娟,殷焱芳,王震,陈红印,刘晨曦. 2021. 蠋蝽在饲养中的品质退化观察[J]. 中国生物防治学报,38(1):159-165. [Zhu Y J,Yin Y F,Wang Z,Chen H Y,Liu C X. 2021. A note of degeneration of predaceous Arma chinensis (Heteroptera:Pentatomidae) during the rearing process[J]. Chinese Journal of Biological Control,38(1):159-165.] doi:10.16409/j.cnki.2095-039x.2021.03.005.
Gao P,Liao X B,Wu G X,Li L F,Lan M X,Hussain M,Chen B,Tang G W,Xie Y H,Gao X. 2021. Gross morphology and ultrastructure of the salivary glands of the stink bug predator Eocanthecona furcellata(Wolff)[J]. Microscopy Research and Technique,84(2):246-252. doi:10.1002/jemt.23581.
Kumar V,Morrison M N,Babu A M,Thiagarajan V. 2002. Egg shell architecture of the stink bug,Eocanthecona furcellata (Wolff):Ultrastructure of micropylar processes and egg burster[J]. International Journal of Tropical Insect Science,22(1):67-73. doi:10.1017/S174275840001 5071.
Lenin E A,Rajan S J. 2016. Biology of predatory bug Eocanthecona furcellata(Wolff)(Hemiptera:Pentatomidae) on Corcyra cephalonica stainton[J]. Journal of Entomology and Zoology Studies,4(3):338-340.
Tuan S J,Yeh C C,Atlihan R,Chin H. 2016. Linking life table and predation rate for biological control:A comparative study of Eocanthecona furcellata (Hemiptera: Pentatomidae) fed on Spodoptera litura (Lepidoptera:Noctui-dae) and Plutella xylostella(Lepidoptera:Plutellidae)[J]. Journal of Economic Entomology,109(1):13-24. doi:10. 1093/jee/tov265.
Zhao H,Liang C,Gao P,Xie Y H,Wang Z J,Wu G X,Tang G W,Chen B,Gao X. 2021. Observation of the fine structure of antennal sensilla of the stink bug,Eocanthe-cona furcellata(Hemiptera:Pentatomidae)[J]. Micron,150:103143. doi:10.1016/j.micron.2021.103143.
收稿日期:2021-11-17
基金項目:云南省基础研究计划项目(202001AT070139)
通讯作者:高熹(1980-),https://orcid.org.000-0003-0990-3452,博士,副教授,主要从事害虫生物防治研究工作,E-mail:chonchon@163.com
第一作者:张曼(1998-),https://orcid.org/0000-0002-1501-4638,研究方向为害虫生物防治,E-mail:zh.ms@foxmail.com
