广西杉木炭疽病病原鉴定及生物学特性测定
2022-07-14廖旺姣邹东霞罗辑吴耀军黄华艳
廖旺姣 邹东霞 罗辑 吴耀军 黄华艳




摘要:【目的】明確广西杉木炭疽病的病原种类及生物学特性,为杉木抗病育种及杉木炭疽病的防治技术研究提供科学依据。【方法】采集广西河池、百色、桂林、柳州、贺州和南宁市杉木种子园及林地炭疽病样品,采用常规组织和单孢分离法获得杉木炭疽病病原菌菌株,通过形态特征结合病原菌核糖体内转录间隔区(ITS)、几丁质合成酶(CHS1)、3-磷酸甘油醛脱氢酶(GPDH)、微管蛋白(TUB2)和肌动蛋白(ACT)多基因分子系统学分析,对获得的炭疽病菌菌株进行鉴定;采用平板培养法测定病原菌生物学特性。【结果】从杉木炭疽病样品中共分离获得60株炭疽菌属真菌,均具有致病性,但致病力存在差异。通过形态学结合病原菌多位点基因系统发育分析,确定广西杉木炭疽病病原菌种类为核果炭疽菌(Colletotrichum fructicola)和山茶炭疽菌(C. camelliae)。生物学特性测定结果显示,C. fructicola和C. camelliae菌丝生长及产孢最适温度分别为28和25 ℃;光照对C. fructicola菌丝生长无明显影响,但有利于C. camealliae菌丝生长,黑暗有利于2种病原菌产孢;pH 4时最适宜2种病原菌菌丝生长,pH 4最适宜C. fructicola产孢,pH 5最适宜C. camealliae产孢;2种病原菌对D-麦芽糖利用最好,乳糖和D-山梨醇最有利于C. fructicola产孢,可溶性淀粉和阿拉伯糖最有利于C. camelliae产孢;蛋白胨和酵母粉均有利于2种病原菌菌丝生长,酵母粉有利于C. fructicolaz产孢,酵母粉和牛肉膏有利于C. camelliae产孢。【结论】广西杉木炭疽病病原为核果炭疽菌(C. fructicola)和山茶炭疽菌(C. camelliae)。温度、pH和碳氮源对2种病原菌菌丝生长及产孢影响明显,光照对2种病原的影响略有不同。
关键词: 杉木;炭疽病;病原菌鉴定;生物学特性;广西
中图分类号: S763.15 文献标志码: A 文章编号:2095-1191(2022)04-1040-09
Identification and biological characteristics of the pathogens
of Chinese fir anthracnose in Guangxi
LIAO Wang-jiao, ZOU Dong-xia, LUO Ji, WU Yao-jun, HUANG Hua-yan*
(Guangxi Forestry Research Institute/Guangxi Engineering Research Center of Forest Pests’
Natural Enemies Breeding, Nanning, Guangxi 530002, China)
Abstract:【Objective】To identify and determine the biological characteristics of the pathogens causing Chinese fir anthracnose in Guangxi to provides a basis for their control. 【Method】Typical anthracnose samples were collected from Chinese fir seed orchards and woodlands in Hechi,Baise,Guilin,Liuzhou,Hezhou,and Nanning,in Guangxi,and thefungiwere isolated by tissue separation and purification. Their pathogenicity was tested according to Koch’s method and their identity determined through morphological studies and multi-gene phylogenetic analysis of their internal transcribed spacer(ITS),chitin synthelase 1(CHS1),glyceraldehyde 3-phosphoric acid dehydrogenase(GPDH),tubnlin2(TUB2), actin(ACT) sequences.The biological characteristics of Colletotrichum fructicola and C. camelliae were determined by the plate culture method. 【Result】A total of 60 strains of anthracnose fungi were isolated from collection sites and the all of the isolates could infect healthy Chinese fir leaves,but with different pathogenicities. The pathogens were identified as C. fructicola and C.camelliae from their phenotypic characteristics and multilocus sequence analysis. The study of their biological characteristics indicated that the optimum temperature for mycelial growth and sporulation of C. fructicola and C. camelliae was 28 and 25 ℃,respectively. Light had no obvious effect on the mycelial growth of C. fructicola,but was beneficial to the mycelial growth of C. camelliae. Complete darkness was beneficial to the spore production of the two pathogens. The optimum pH value for mycelial growth of the two pathogens was 4,and the optimum pH value for spore production was 4 and 5 for C. fructicola and C. camelliae,respectively. The better carbon sources for C. fructicola and C. camelliae was D-maltose for mycelium growth,whereas the optimal carbon source for spore germination was lactose and D-sorbitol for C. fructicola,and soluble starch and arabinose in C. camelliae. The best nitrogen sources for C. fructicola and C. camelliae were peptone and yeast powder during mycelium growth,and the optimal nitrogen for spore germination was yeast powder in C. fructicola,and yeast powder and beef extract in C. camelliae. 【Conclusion】The pathogens of Chinese fir anthracnose in Guangxi are identified as C. fructicola and C. camelliae. Temperature,pH,carbon and nitrogen sources have significant effects on mycelial growth and sporulation of C. fructicola and C. camelliae,while the effects of light on the two pathogens are slightly different.
Key words: Cunninghamia lanceolata(Lamb.) Hook.; anthracnose; pathogen identification; biological characteristics; Guangxi
Foundation items:Guangxi Innovation Driven Development Project(Guike AA17204087-10); Guangxi Forestry Science and Technology Project (Guilingke〔2016〕No.4)
0 引言
【研究意義】杉木[Cunninghamia lanceolata (Lamb.)Hook.]是我国南方最重要的用材林树种之一。杉木生长快、产量高、材质好、用途广,其木材是优秀的建筑用材、装饰用材和纸浆用材(陈代喜和李富福,2013)。目前,广西杉木面积约150.38万ha,占全区人工林总面积的25.38%,是国家木材战略储备广西核心基地建设的重要树种(戴俊等,2020)。炭疽病是危害广西杉木的主要病害,主要危害幼树先年秋梢,致使针叶干枯或梢头枯死,严重时幼树整株死亡。2015—2020年,炭疽病在广西各杉木栽培区普遍发生,种子园和林地发病率30%~80%,严重者发病率可高达100%,极大地影响杉木健康生长,降低球果结实率,种子品质变差,甚至绝收,严重影响杉木种子的产量和品质。关于杉木炭疽病病原一直沿用胶孢炭疽菌(Colletotrichum gloeosporioides)(黄飞龙,1993),但该菌是复合种群,仅凭形态学特征很难区分复合种群内的合格种,而形态学结合多基因系统学分析能有效区分复合种群内的合格种(Weir et al.,2012)。目前,广西杉木炭疽病的研究主要集中在病害发生和防治等方面,尚缺乏病原菌的准确鉴定和生物学特性研究。准确鉴定杉木炭疽病病原并掌握其生物学特性是明确病害发生流行动态和制定有效防治措施的基础(孔前前等,2018;伍晓丽等,2021),同时,也是杉木种子园良种抗病选育的前提基础。因此,明确广西杉木炭疽病的病原种类及生物学特性,对杉木良种抗性选育及防治措施制定具有重要的指导意义。【前人研究进展】杉木炭疽病最早发现于20世纪70年代,而后李传道等(1980)和林业部南方森林植物检疫所等(1980a,1980b)对江西省杉木炭疽病进行调查研究,依据病原菌形态特征确定病原菌为胶孢炭疽菌,明确了发病规律。近年来报道的杉木炭疽病病原与此相同或不同,田龙艳等(2019)采用形态特征结合ITS等基因序列,确定粤北地区杉木炭疽病病原亦为胶孢炭疽菌;韩雨庭等(2021)采用形态特征结合ITS基因系统分析法明确云南省广南县杉木炭疽病病原为暹罗刺盘孢(C. siamense),该病原与前人所报道的胶孢炭疽菌不同。炭疽病的空间分布及病害调查方法已有研究报道,朱建华等(2004)报道林地炭疽病空间分布类型为聚集分布;伍南等(2012)利用遥感高光谱微分指数估测杉木炭疽病病情指数精度较高,且具有较高的实际应用价值,解决了传统的人力踏查耗时耗力,效率和准确性低,无法大面积实时监测的难题。病害防治方面的研究报道较多,传统的防治措施主要以营林为主,化学防治为辅,适地造林、合理施用化肥,促进杉木健康生长,提高抗病能力;病害发生严重时,使用百菌清和甲基托布津能有效防止病害的蔓延(肖斌和肖力,2017)。杉木炭疽病生物防治也有相关报道,路宗岩等(2013)发现一种地衣芽孢杆菌HY32对杉木炭疽病有较明显的防治效果;李红军等(2014)对HY32发酵条件进行优化,以期提高对杉木炭疽病的防治效果;李冬琴等(2015)从杉木根际土壤筛选到一株对炭疽菌有良好抑制作用的拮抗细菌AM53,而且研制出的相应菌剂在林间的防治效果良好。杉木炭疽病病原菌生物学特性研究报道较少,曾思海等(1998)报道了球果炭疽病菌(Colletotrichum sp.)生物学特性,适宜该菌的温度为22~30 ℃,以26 ℃为菌丝最适生长温度,24 ℃为分生孢子萌发最适温度;可生长的酸碱度为pH 5~8,以pH 7为最适酸碱度;分生孢子以在水滴和100%相对湿度中萌发率最高;菌丝在常见碳源蔗糖、常见氮源蛋白胨上生长良好。但该菌没有准确鉴定到种。【本研究切入点】前人依据形态学特征对广西杉木炭疽病病原进行鉴定,该方法很难区分近似种,病原菌分类准确性有待商榷。20世纪90年代至今未见采用形态学与多基因分子系统学分析对广西杉木炭疽菌进行鉴定的报道,病原菌生物学特性亦未见报道。【拟解决的关键问题】采用五点取样法和随机取样法分别对广西河池、百色、桂林、柳州、贺州和南宁市等杉木种子园和杉木林地炭疽病进行全面调查,采用常规组织和单孢分离法获得炭疽病病原菌,通过形态特征与多基因分子系统学分析相结合的方法对炭疽病菌菌株进行鉴定;采用平板培养法分析影响病原菌菌丝生长及产孢的温度、光照、pH、碳源和氮源等因子,明确广西杉木炭疽病病原菌的分类地位和生物学特性,为后续开展抗病育种及防治技术研究提供科学依据。
1 材料与方法
1. 1 试验材料
供试植物材料:杉木炭疽病样品来源于广西河池、百色、桂林、柳州、贺州和南宁市杉木种子园和杉木林地;健康杉木枝条采自广西林业科学研究院杉木试验林。
培养基:马铃薯葡萄糖琼脂培养基(PDA):马铃薯200.00 g、葡萄糖20.00 g、琼脂粉20.00 g、水1000 mL。查彼培养基(Czapek):硝酸钠2.00 g、磷酸氢二钾1.00 g、氯化钾0.50 g、七水合硫酸镁0.50 g、硫酸铁0.01 g、蔗糖30.00 g、琼脂粉20.00 g、水1000 mL。
主要试剂及仪器:DNA提取试剂盒和PCR扩增试剂盒(2×Es Taq Master Mix)均购自天根生化科技(北京)有限公司;其他试剂均为国产分析纯。Biometra PC48型PCR仪(德国BIOMETRE公司);DYCP-31DN型电泳仪(北京六一仪器厂)。
1. 2 试验方法
1. 2. 1 病原菌分离纯化 病针叶冲洗干净后,依次浸泡75%酒精1~3 min,0.1%升汞4~5 min,无菌水冲洗3次。灭菌剪刀在病健交界处剪取约1.00 cm长组织,接种到PDA培养基上,置于28 ℃下培养5 d,组织块长出菌丝和分生孢子后,挑取单孢接种在新的PDA培养基上培养7 d,菌株保存至PDA斜面试管培养基,4 ℃冰箱保藏(方中达,2001)。
1. 2. 2 致病性测定 取杉木健康嫩枝,经75%酒精表面消毒和无菌水冲洗干净后用直径0.30 cm的卫生香在近针叶叶尖处烫一个直径约0.20 cm的伤口,解剖刀挑取0.30 cm病原菌菌饼接种到伤口上,以无菌PDA接种为对照,处理和对照各20张针叶,接种后置于底部铺有滤纸的白色透明塑料盒中,脱脂棉蘸灭菌水保湿。置于室温中培养,10 d后观察是否发病,统计发病率和测量病斑长度。取发病针叶进行病原菌再次分离,与接种菌进行比较是否相同,从而确定致病菌。
1. 2. 3 病原菌鉴定
1. 2. 3. 1 病原菌形态鉴定 纯化菌株置于PDA培养基25 ℃恒温、黑暗条件下培养,逐日观察。培养7 d后,记录菌株培养性状和菌落直径,镜检、拍摄和测量100个病原分生孢子和20个分生孢子附着胞大小(杨友联,2010)。
1. 2. 3. 2 病原菌基因序列测定及多基因系统分析
病原菌基因组DNA提取参照试剂盒使用说明操作,选择真菌核糖体基因转录间隔区(ITS,ITS4/ITS5)、几丁质合成酶(CHS1,CHS1-79F/CHS1-784R)、3-磷酸甘油醛脱氢酶(GPDH,GDF1/GDR1)、微管蛋白(TUB2,T1/β-tubulin2b)和肌动蛋白(ACT,ACT-512F/ ACT-783R)基因进行扩增,PCR扩增反应体系和条件、系统发育进化树构建等参考朱英芝等(2015)的方法。
1. 2. 4 病原菌生物学特性测定
1. 2. 4. 1 温度对菌丝生长和产孢的影响 从分离获得的杉木炭疽病病原菌中选择2株菌株作为代表菌株进行生物学特性测定。将直径为6 mm(下同)的病原菌菌饼接种至新的PDA培养基上,置于黑暗条件下分别于5、10、15、20、25、28、30和35 ℃培养,每处理6个重复,7 d后用十字交叉法测定菌落直径,10 d后用血球计数板测定产孢量(方中达,2001)。
1. 2. 4. 2 光照对菌丝生长和产孢的影响 取病原菌菌饼接至PDA培养基上,分别置于全光照(8 W,灯皿距离20 cm,下同)、全黑暗和12 h光暗交替25 ℃恒温培养,菌落直径及测定方法同1.2.4.1。
1. 2. 4. 3 不同pH对菌丝生长和产孢的影响 取病原菌菌饼接至不同pH的PDA(1 mol/L的HCl和NaOH调制)培养基上,25 ℃黑暗培养,菌落直径及测定方法同1.2.4.1。
1. 2. 4. 4 不同碳源对菌丝生长和产孢的影响 以不含蔗糖的查彼培养基为基础,分别配制D-麦芽糖、乳糖和阿拉伯糖等10种碳源培养基,取病原菌菌饼接种至上述含不同碳源的培养基上,菌落直径及测定方法同1.2.4.1。
1. 2. 4. 5 不同氮源对菌丝生长和产孢的影响 以不含NaNO3的查彼培养基为基础,分别配制L-赖氨酸、硫酸铵和蛋白胨等8种氮源培养基,取病原菌菌饼接种至上述含不同氮源的培养基上,菌落直径及测定方法同1.2.4.1。
1. 3 统计分析
试验数据采用Excel 2010整理,数据分析及差异显著性检验采用DPS 13.01中的Duncan’s新复极差法。
2 结果与分析
2. 1 病原菌分离及致病性测定结果
从采集自广西河池、百色、桂林、柳州、贺州和南宁市的杉木炭疽病样品中共分离获得60株菌株,分为两大类型,其中CSHC01、CSNN01、CSBS01、CSGL01和CSHZ01为第一种类型,共37株;CSHC02、CSNN02、CSLZ01、CSBS02和CSGL02为第二种类型,共23株。选择CSHC01和CSHC02为测试菌株,菌株接种健康杉木针叶3 d后,接种点褐变开始扩展,10 d后接种的针叶均感病,病斑扩展至针叶1/4~3/4,症状与自然感病症状相同(图1-A和图1-B);对照接种点变褐,但未扩展(图1-C)。感病针叶及对照针叶进行再次分离,接种处理针叶获得的菌株与原接种菌株一致,对照处理未获得任何菌株,依据柯赫氏法则,确定接种菌即为杉木炭疽病致病菌。
2. 2 病原菌形态特征
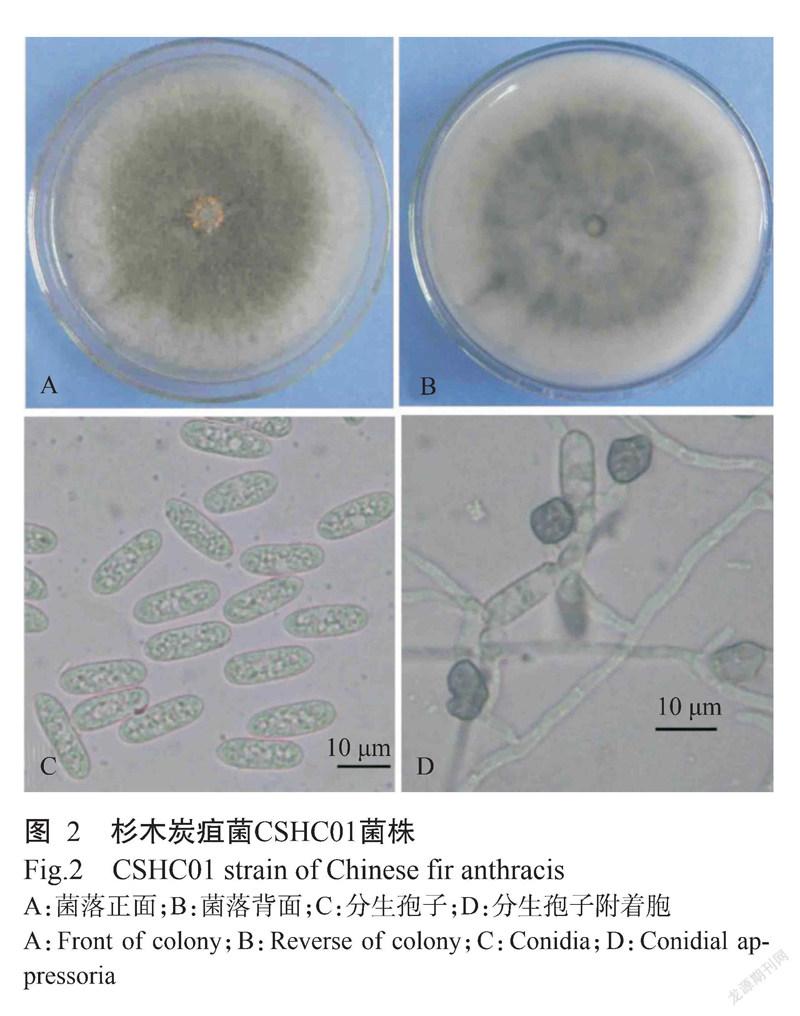
CSHC01菌株在PDA培养基上培养7 d,菌落平均直径为82.96 mm,平均生长速率11.85 mm/d,菌落圆形,气生菌丝绒毛状,灰色至灰黑色,边缘整齐,背面产生黑色色素(图2-A和图2-B)。分生孢子圆柱状,两端钝圆或一端稍尖,无色,单胞,光滑,具有1~2个油球,平均大小为(20.04±1.92)μm×(5.42±0.57)μm(图2-C)。分生孢子附著胞圆形或近椭圆形,浅褐色至褐色,少数呈不规则形,边缘完整,单个或多个,平均大小为(9.85±0.87)μm×(7.85±0.42)μm(图2-D),形态特征与核果炭疽菌(C. fructicola)相符(杨友联等,2014)。
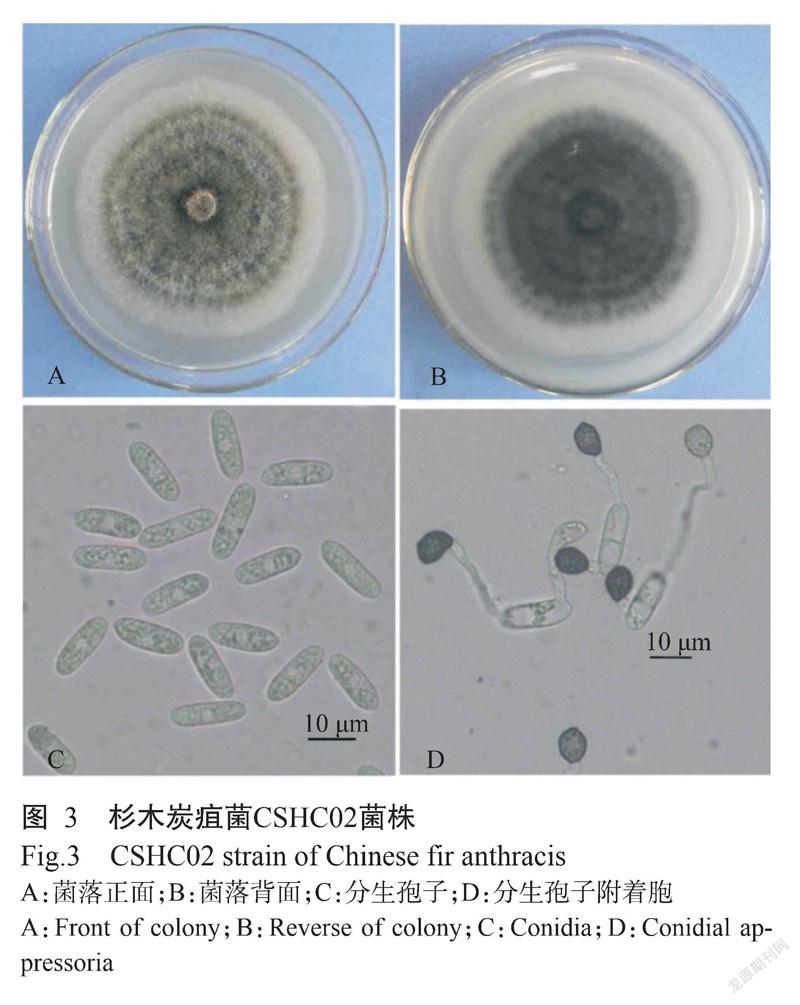
CSHC02菌株在PDA培养基上培养7 d,菌落平均直径为78.28 mm,平均生长速率11.18 mm/d,菌落近圆形,气生菌丝棉花状,白色至灰色,边缘整齐,背面灰色至黑色(图3-A和图3-B)。分生孢子圆柱状,两端钝圆或一端略粗,另一端稍尖,无色,单孢,光滑,平均大小为(15.90±1.07)μm×(5.17±0.35)μm(图3-C);附着胞近圆形,褐色至黑褐色,单个或多个,平均大小为(7.92±0.71)μm×(6.24±0.58)μm(图3-D)。形态特征与山茶炭疽菌(C. camelliae)相符(李河等,2017)。
2. 3 病原菌分子生物学鉴定结果
获得菌株的ITS、CHSI、GPDH、TUB2和ACT基因序列后,在GenBank数据库中对各基因序列分别进行比对,下载相关序列,用MEGA 5.0中的邻接法构建上述5个基因系统发育进化树。结果(图4)显示,CSBS01、CSGL01、CSHZ01、CSHC01和CSNN01菌株与C. fructicola菌株聚在同一分支,相似性达100%,CSLZ01、 CSGL02、CSNN02、CSHC02和CSBS02菌株与C. camelliae菌株聚在同一分支上,相似性达100%,遗传进化距离最近,能与炭疽菌其他种区分,各分支支持率均较高(图4)。分子生物学分析结果表明,广西杉木炭疽病病原菌为核果炭疽菌(C. fructicola)和山茶炭疽菌(C. camelliae)。
2. 4 病原菌生物学特性测定结果
2. 4. 1 温度对病原菌菌丝生长及产孢的影响 试验结果(表1)显示,C. fructicola菌丝生长温度为10~35 ℃,温度低于5 ℃菌丝不生长,最适生长温度为25~30 ℃,其中以28 ℃处理最好,菌落直径显著大于除25和30 ℃处理外的其他温度处理(P<0.05,下同),超過30 ℃菌丝生长急剧下降,至35 ℃时菌落直径仅为10.09 mm;C. fructicola在15~28 ℃均能产孢,其中以25 ℃产孢量最大,其次是20 ℃,2个温度处理间产孢量差异显著。C. camealliae菌丝生长温度为10~30 ℃,最适生长温度为28 ℃,其次是25 ℃,各温度处理间菌落直径差异显著;C. camealliae在15~30 ℃均能产孢,其中以25 ℃产孢量最大,其次是28 ℃。2株菌株菌丝生长温度范围基本相同,菌丝生长最适温度和产孢最适温度一致,分别为28和25 ℃。
2. 4. 2 光照对病原菌菌丝生长和产孢的影响 由表2可知,C. fructicola在3种光照条件下菌丝生长差异不明显,菌落直径差异不显著(P>0.05,下同);产孢量存在一定差异,以全黑暗处理产孢量最大,其次是全光照处理,两者间产孢量差异不显著,最差的是12 h光暗交替处理,产孢量与全光照处理差异显著。C. camealliae在全光照条件下生长最好,其次是全黑暗处理,两者间菌落直径差异不显著;产孢量以全黑暗处理最高,其次是12 h光暗交替处理,全光照处理最少,各处理间产孢量差异显著。
2. 4. 3 不同pH对病原菌菌丝生长及产孢的影响
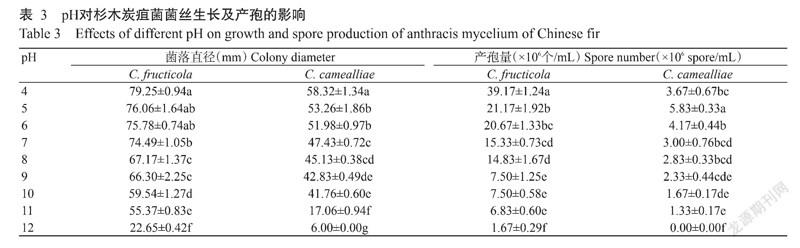
由表3可知,C. fructicola在pH 4~12范围内均能生长及产孢,其中以pH 4处理菌丝生长及产孢量最大,其次为pH 5和pH 6处理,菌丝生长及产孢量均随着pH升高而降低;C. camealliae菌丝生长及产孢pH范围较C. fructicola略小,pH 4处理菌丝生长最好,pH 5处理产孢量最大,其次为pH 6处理,此后产孢量随着pH升高而降低。
2. 4. 4 不同碳源对病原菌菌丝生长及产孢的影响
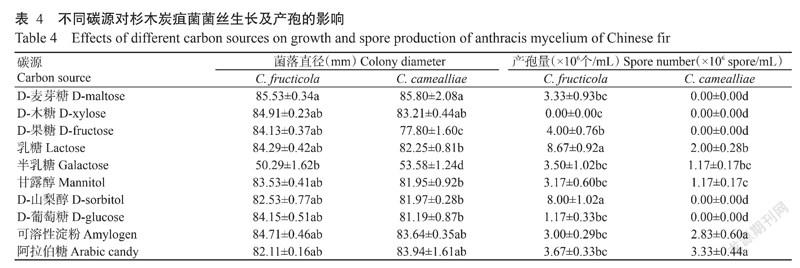
试验结果(表4)显示,C. fructicola在供试的10种碳源培养基上均能生长,其中以D-麦芽糖处理生长最好,其次是D-木糖、可溶性淀粉、乳糖、D-葡萄糖、D-果糖、甘露醇、D-山梨醇和阿拉伯糖处理,9种碳源处理间菌落直径差异不显著,在半乳糖处理上生长最差;C. fructicola可在除D-木糖外的9种碳源培养基上产孢,以乳糖和D-山梨醇处理产孢量最大。C. camealliae亦可在供试的10种碳源培养基生长,生长最佳碳源与C. fructicola相同,均为D-麦芽糖,其次是D-木糖、可溶性淀粉和阿拉伯糖,生长最差的碳源与C. fructicola相同,为半乳糖,与其他碳源处理菌落直径差异显著;C. camealliae只在乳糖、半乳糖、甘露醇、可溶性淀粉和阿拉伯糖培养基上产孢,其中在阿拉伯糖和可溶性淀粉培养基上产孢量最大,两者的产孢量与其他碳源处理差异显著。
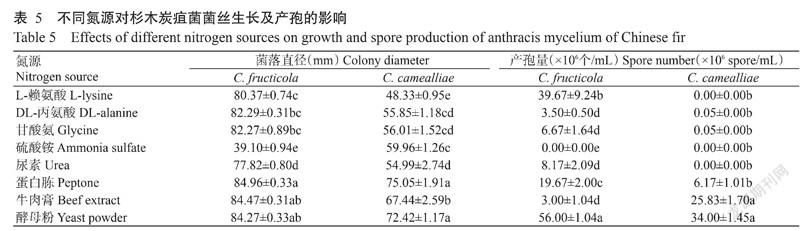
2. 4. 5 不同氮源对病原菌菌丝生长及产孢的影响
试验结果(表5)表明,C. fructicola在供试的8种氮源培养基上均能生长,其中以蛋白胨处理生长最好,其次是牛肉膏和酵母粉处理,3种有机氮源处理的菌落直径差异不显著,说明有机氮源有利于该菌的菌丝生长;C. fructicola在除硫酸铵处理外的7种氮源培养基上均能产孢,其中以酵母粉处理的产孢量最大,其次是L-赖氨酸处理,在牛肉膏处理的产孢量最少。C. camealliae与C. fructicola相似,可在供试的8种氮源培养基上生长,其中以蛋白胨和酵母粉处理生长最好,其次是牛肉膏处理;C. camealliae只在DL-丙氨酸、甘酸氨、蛋白胨、牛肉膏和酵母粉培养基上产孢,以酵母粉处理的产孢量最大,其次是牛肉膏处理,DL-丙氨酸和甘酸氨处理的产孢量最少。
3 讨论
3. 1 广西杉木炭疽病病原
本研究确定广西杉木炭疽病病原为C. fructicola和C. camelliae,且以C. fructicola为优势病原。与李传道等(1980)和林业部南方森林植物检疫所等(1980a,1980b)报道的C. gloeosporioides不同,也与韩雨庭等(2021)报道的C. siamense不同。C. fructicola寄主范围较广,可危害小粒咖啡(Coffea arabica)、沙梨(Pyrus pyrifolia)、深波叶补血草(Limonium sinense)、葡萄(Vitis vinifera)、草莓(Fragaria ananassa)、茶树(Camellia sinensis)、棉花(Gossypium hirsutum)、油茶(C. oleifera)和土沉香(Aquilaria sinensis)等植物(Weir et al.,2012;杨友联等,2014;李河等,2017;廖旺姣等,2018)。C. camelliae寄主范围较C. fructicola窄,目前已报道的寄主为油茶、茶树和金花茶(C. nitidissima)(谢玲等,2009;李河等,2017;王雪等,2020),报道的3种寄主均为山茶科植物。
C. fructicola和C. camelliae是油茶炭疽病重要病原(吴耀军和奚福生,2010;李河等,2017),此次在广西杉木上发现,说明杉木是这2种病原菌的新记录寄主。在河池、百色、南宁和桂林市的杉木上2種病原菌均有发现,在贺州的杉木上只发现C. fructicola,在柳州的杉木上只发现C. camelliae,可能与采集样品范围有关。百色、河池、柳州和贺州市是杉木主要栽培区,同时是广西油茶的重要栽培区,桂林和南宁市也分布有大量的油茶林。杉木炭疽病与油茶炭疽病病原是否有关联有待进一步研究。
3. 2 广西杉木炭疽病病原C. fructicola和C. camealliae生物学特性
对广西杉木炭疽病病原生物学特性测定结果表明,C. fructicola最适生长温度为25~30 ℃,其中以28 ℃最好,最佳产孢温度为25 ℃。C. fructicola菌丝生长及产孢最佳温度与刘倩丽等(2014)报道的海南檀香炭疽病菌(C. fructicola)最适生长温度及分生孢子最适萌发温度为30 ℃相比偏低;与宋丽丽等(2019)报道草莓炭疽病菌(C. fructicola)最适生长温度范围一致,但最佳生长温度较草莓炭疽病菌(30 ℃)略低。可能与广西杉木生长的环境温凉有关(王献溥等,2004)。C. fructicola适宜生长的pH为4~12,与檀香炭疽病菌菌丝生长适宜的pH 4~11(刘倩丽等,2014)略宽;C. fructicola最适宜生长的pH为4,比檀香炭疽病菌最适pH为6(刘倩丽等,2014)和草莓炭疽病菌最佳pH为6~7(宋丽丽等,2019)不同,C. fructicola适合偏酸条件下生长;光照对C. fructicola生长无明显影响,与刘倩丽等(2014)报道光照有利于檀香炭疽病菌菌丝生长的结论存在差异;黑暗有利于产孢,两者结论一致;C. fructicola在可溶性淀粉等碳源上生长较好,与檀香炭疽病菌(刘倩丽等,2014)的研究结果一致;C. fructicola在蛋白胨氮源培养基上生长良好,与檀香炭疽病菌(刘倩丽等,2014)的研究结论一致;本研究中,C. fructicola在硫酸铵氮源培养基上生长较差,而檀香炭疽病菌(刘倩丽等,2014)却生长良好,可能与病原菌的不同寄主来源有关。
C. camealliae最适生长温度为28 ℃,与王雪等(2019)报道的贵州茶褐枯病病原菌(C. camelliae)最适生长温度结论一致;C. camealliae菌丝生长最适pH为4,与茶褐枯病病原菌最适pH为4.5~6.0(王雪等,2019)存在差异,本研究的C. camealliae适宜在偏弱酸条件下生长;C. camealliae在蛋白胨氮源培养基上生长最好,茶褐枯病病原菌在蛋白胨培养基上生长一般(王雪等,2019),可能与菌株不同寄主来源有关。
本研究的C. fructicola和C. camealliae从杉木种子园分离获得,研究发现这2种炭疽菌与不同寄主来源的菌株的生物学特性存在一定差异,可能与菌株的遗传特性有关,有待进一步探究。
4 结论
采用形态学结合病原菌的ITS、CHS1、GPDH、TUB2和ACT基因分子系统学分析,确定广西杉木炭疽病病原菌为核果炭疽菌(C. fructicola)和山茶炭疽菌(C. camelliae),以C. fructicola分离率较高,这是国内杉木炭疽病新病原首次报道,杉木是这2种炭疽菌的新寄主。温度、pH、碳氮源对C. fructicola和C. camelliae菌丝生长及产孢影响明显,光照对C. fructicola菌丝生长无明显影响,但对C. camelliae有影响,黑暗条件均利于产孢。
参考文献:
陈代喜,李富福. 2013. 广西林木良种基地建设与育苗新技术[M]. 南宁:广西科学技术出版社:41-44. [Chen D X,Li F F. 2013. Construction of Guangxi forest seed base and new seedling technology[M]. Nanning:Guangxi Science and Technology Press:41-44.]
戴俊,陈代喜,黄开勇,董利军,黄鹏艳,罗启亮,谭文靖,王晓波,唐红亮. 2020. 广西杉木第二代良种林分生长量调查分析[J]. 广西林业科学,49(2):237-240. [Dai J,Chen D X,Huang K Y,Dong L J,Huang P Y,Luo Q L,Tan W J,Wang X B,Tang H L. 2020. Growths investigation and analysis on second generation improved variety stands of Cunninghamia lanceolata in Guangxi[J]. Guangxi Fore-stry Science,49(2):237-240.] doi:10.19692/j.cnki.gfs. 2020.02.014.
方中达. 2001. 植病研究方法[M]. 第3版. 北京:中国农业出版社:122-125. [Fang Z D. 2001. Plant disease research method[M]. The 3rd Edition. Beijing:China Agriculture Press:122-125.]
韩雨庭,唐婕,尼玛此姆,马焕成,伍建榕. 2021. 广南县杉木三种侵染性病害病原鉴定[J]. 中国森林病虫,40(5):1-8. [Han Y T,Tang J,Nima C M,Ma H C,Wu J R. 2021. Pathogen identification of three infectious diseases of Chinese fir in Guangnan County[J]. Forest Pest and Disea-se,40(5):1-8.] doi:10.19688/j.cnki.issn1671-0886.2021 0016.
黄飞龙. 1993. 关于广西杉木林病害发生和防治的探讨[J]. 广西林业科技,22(3):103-107. [Huang F L. 1993. Discussion on the occurrence and prevention of diseases in the fir forests in Guangxi[J]. Guangxi Forestry Science and Technology,22(3):103-107.] doi:10.19692/j.cnki.gfs.1993.03002.
孔前前,阮柳,劉东霞,刘振宇,张雨竹,秦丰,袁军海,马占鸿,王海光. 2018. 河北苜蓿镰孢菌根腐病病原生物学特性研究[J]. 中国农业大学学报,23(8):59-76. [Kong Q Q,Ruan L,Liu D X,Liu Z Y,Zhang Y Z,Qin F,Yuan J H,Ma Z H,Wang H G. 2018. Biological characteristics of Fusarium causing alfalfa root rot in Hebei Province[J]. Journal of China Agricultral University,23(8):59-76.] doi:10.11841/j.issn. 1007-4333.2018.08.07.
李传道,朱熙櫵,石峰云. 1980. 杉木炭疽病的研究——Ⅱ. 病原菌的鉴定[J]. 南京林业大学学报(自然科学版),(3):28-34. [Li C D,Zhu X Q,Shi F Y. 1980. Studies on the anthracnose of Chinese fir Ⅱ. identification of the causal fungus[J]. Journal of Nanjing Forestry University(Natural Science Edition),(3):28-34.]
李冬琴,谭益民,路宗岩,周国英. 2015. 杉木炭疽病拮抗菌AM53菌剂的研制及其林间防效[J]. 福建农林大学学报(自然科学版),44(2):154-158. [Li D Q,Tan Y M,Lu Z Y,Zhou G Y. 2015. Development and forest control of antagonistic bacteria AM53 against Colletotrichum gloeosporioides on Chinese fir[J]. Journal of Fujian Agriculture and Forestry University(Natural Science Edition),44(2):154-158.] doi:10.13323/j.cnki.j.fafu(nat.sci.).2015. 02.008.
李河,李杨,蒋仕强,刘君昂,周国英. 2017. 湖南省油茶炭疽病病原鉴定[J]. 林业科学,53(8):43-53. [Li H,Li Y,Jiang S Q,Liu J A,Zhou G Y. 2017. Pathogen of oil-tea trees anthracnose caused by Colletotrichum spp. in Hunan Province[J]. Scientia Silvae Sinicae,53(8):43-53.] doi:10.11707/j.1001-7488. 20170806.
李红军,周国英,路宗岩,谭益民,段爱国. 2014. 杉木炭疽病拮抗菌HY32的发酵条件优化[J]. 林业科学,50(12):94-100. [Li H J,Zhou G Y,Lu Z Y,Tan Y M,Duan A G. 2014. Optimization of fermentation medium formula and culture conditions for the antagonistic bacterium HY32 against Colletotrichum gloeosporioides[J]. Scientia Silvae Sinicae,50(12):94-100.] doi:10.11707/j.1001-7488. 20141213.
廖旺姣,邹东霞,黄华艳,罗辑,赵程劼,吴耀军. 2018. 土沉香幼苗炭疽病病原分离与鉴定[J]. 南方农业学报,49(1):74-78. [Liao W J,Zou D X,Huang H Y,Luo J,Zhao C J,Wu Y J. 2018. Isolation and identification for pathogen of anthracnose in Aquilaria sinensis(Lour.) Gilg.[J]. Journal of Southern Agriculture,49(1):74-78.] doi:10.3969/j.issn.2095-1191.2018.01.12.
林业部南方森林植物检疫所,南京林产工业学院,江西省进贤县北岭林场. 1980a. 杉木炭疽病的研究——Ⅰ. 症状和病原的探讨[J]. 南京林业大学学报(自然科学版),(2):31-38. [South-China Forest-plant Quarantina Service,Nanjing Technological Colloge of Forest Products,Beiling Forestry Station,Jinxian County,Jiangxi Provin-ce. 1980a. Studies on the anthracnose of Chinese firⅠ.Symptoms and the causal agent[J]. Journal of Nanjing Fores-try University (Natural Science Edition),(2):31-38.]
林业部南方森林植物检疫所,南京林产工业学院,江西省进贤县北岭林场. 1980b. 杉木炭疽病的研究——Ⅲ. 病害流行[J]. 南京林产工业学院学报,(4):16-22. [South-China Forest-plant Quarantina Service,Nanjing Technological College of Forest Products,Beiling Forestry Station,Jinxian County,Jiangxi Province. 1980b. Studies on the anthracnose of Chinese fir Ⅲ. Epiphytotic of the disease[J]. Journal of Nanjing Academy of Forestry Industry,(4):16-22.]
刘倩丽,周国英,刘成锋,田媛媛,倪先鹏,刘君昂. 2014. 檀香炭疽病病原鉴定及其生物学特性研究[J]. 热带作物学报,35(11):2266-2273. [Liu Q L,Zhou G Y,Liu C F,Tian Y Y,Ni X P,Liu J A. 2014. Identification and biological characteristics of Collectotrichum fructicola on Santalum album L.[J]. Chinese Journal of Tropical Crops,35(11):2266-2273.] doi:10.3969/j.issn.1000-2561.2014. 11.028.
路宗岩,周国英,陈玉华,闫法领,闫瑞坤,伍南. 2013. 杉木炭疽病拮抗菌HY32的筛选及其应用[J]. 生物技术通报,(3):181-185. [Lu Z Y,Zhou G Y,Chen Y H,Yan F L,Yan R K,Wu N. 2013. Screening and application of the antagonistic bacterium HY32 against Colletotrichum gloeosporioides[J]. Biotechnology Bulletin,(3):181-185.]
宋丽丽,张丽勍,高清華,段可. 2019. 草莓果生刺盘孢菌的生物学特性及致病性测定[J]. 上海农业学报,35(6):88-96. [Song L L,Zhang L Q,Gao Q H,Duan K. 2019. Bio-logical characteristics ang pathogenicity of strawberry Collectotrichum fructicola[J]. Acta Agriculturae Shanghai,35(6):88-96.] doi:10.15955/j.issn.1000-3924.2019. 06.16.
田龙艳,练涛,柯思恺,秦长生,徐金柱,赵丹阳,邱华龙,杨华,靳秀芳,李南林. 2019. 粤北地区杉木真菌性病害种类[J]. 林业环境与科学,35(4):90-96. [Tian L Y,Lian T,Ke S K,Qin C S,Xu J Z,Zhao D Y,Qiu H L,Yang H,Jin X F,Li N L. 2019. The fungal diseases of Chinese fir in Northern Guangdong[J]. Forestry and Environmental Scicence,35(4):90-96.] doi:10.3969/j.issn.1006-4427.2019.04.015.
王献溥,郭柯,田新智. 2004. 广西杉木林的分类、分布和演替[J]. 植物资源与环境学报,13(1):43-47. [Wang X P,Guo K,Tian X Z. 2004. Classification,distribution and succession of Cunninghamia lanceolata(Lamb.) HK. fore-sts in Guangxi,China[J]. Journal of Plant Resources and Environment,13(1):43-47.]
王雪,王勇,尹桥秀,李冬雪,任亚峰,江仕龙,王德炉,宋宝安,陈卓. 2020. 贵州省余庆县茶褐枯病病原菌的鉴定[J]. 植物保护,46(2):101-106. [Wang X,Wang Y,Yin Q X,Li D X,Ren Y F,Jiang S L,Wang D L,Song B A,Chen Z. 2020. Identification of the pathogen of tea brown blight in Yuqing County,Guizhou Province[J]. Plant Protection,46(2):101-106.] doi:10.16688/j.zwbh. 2019225.
王雪,尹桥秀,李冬雪,任亚峰,包兴涛,武娴,王德炉,陈卓. 2019. 茶褐枯病病原菌(Colletotrichum camelliae)生物学特性研究[J]. 中国植保导刊,39(6):5-11. [Wang X,Yin Q X,Li D X,Ren Y F,Bao X T,Wu X,Wang D L,Chen Z. 2019. Biological characteristics of Colletotrichum camelliae causing tea brown blight disease[J]. China Plant Protection,39(6):5-11.] doi:10.3969/j.issn.1672-6820. 2019.06.001.
伍南,刘君昂,周国英,闫瑞坤,董文统. 2012. 基于高光谱微分指数的杉木炭疽病病情指数反演[J]. 林业科学,48(8):94-98. [Wu N,Liu J A,Zhou G Y,Yan R K,Dong W T. 2012. Inversion anthracnose disease indices of Chinese fir basedon hyperspectral derivative indices[J]. Scien-tia Silvae Sinicae,48(8):94-98.]
伍晓丽,陈大霞,刘飞,王钰,李隆云. 2021. 黄连根腐病腐霉属病原菌鉴定[J]. 西南大学学报(自然科学版),43(6):37-43. [Wu X L,Chen D X,Liu F,Wang Y,Li L Y. 2021. Identification of the Pythium pathogen responsible for root rot of Coptis chinensis[J]. Journal of Southwest University (Natural Science Edition),43(6):37-43.] doi:10.13718/j.cnki.xdzk.2021.06.005.
吴耀军,奚福生. 2010. 中国油茶油桐病虫害彩色原生态图鉴[M]. 南宁:广西科学技术出版社:1-2. [Wu Y J,Xi F S. 2010. Illustrated handbook of pests and diseases of Camellia oleifera Abel in China[M]. Nanning:Guangxi Scien-ce and Technology Press:1-2.]
肖斌,肖力. 2017. 杉木病虫害种类调查及防治措施浅析[J]. 生物灾害科学,40(2):104-107. [Xiao B,Xiao L. 2017. A brief analysis of main species and prevention against diseases and pests of Cunninghamia lanceolata[J]. Biological Disaster Science,40(2):104-107.] doi:10.3969/j.issn. 2095-3704.2017.02.024.
谢玲,黄思良,岑贞陆,黄连冬,刘宇宁. 2009. 广西金花茶炭疽病的病原鉴定及其寄主抗性评价(英文)[J]. 广西农业科学,40(8):998-1003. [Xie L,Huang S L,Cen Z L,Huang L D,Liu Y N. 2009. Identification of causal organism of yellow camellia anthracnose in Guangxi and evaluation of camellia species for host resistance[J]. Guangxi Agricultural Sciences,40(8):998-1003.]
楊友联. 2010. 中国贵州、云南、广西炭疽菌属真菌多基因分子系统学分析研究[D]. 武汉:华中农业大学. [Yang Y L. 2010. Multi-locus phylogeny of Colletotrichum species in Guizhou,Yunnan and Guangxi,China[D]. Wuhan:Huazhong Agricultural University.]
杨友联,刘永翔,刘作易. 2014. 棉花炭疽病的病原鉴定[J]. 贵州农业科学,42(8):86-91. [Yang Y L,Liu Y X,Liu Z Y. 2014. Identification of Colletotrichum species associated with cotton anthracnose[J]. Guizhou Agricultural Sciences,42(8):86-91.] doi:10.3969/j.issn.1001-3601.2014.08.024.
曾思海,黄天章,詹学齐,王远浩. 1998. 杉木球果炭疽病病原菌及其生物学特性的研究[J]. 福建林业科技,25(2):6-10. [Zeng S H,Huang T Z,Zhan X Q,Wang Y H.1998. Studies on the pathogenic fungi of cone anthracnose of Cunninghamia lanceolata and on their biological characteristics[J]. Journal of Fujian Forestry Science and Technology,25(2):6-10.] doi:10.13428/j.cnki.fjlk.1998.02.002.
朱建华,陈红梅,傅瑞树,叶剑雄,杨爱民. 2004. 杉木炭疽病空间格局及变化研究J]. 中国生态农业学报,12(3):169-171. [Zhu J H,Chen H M,Fu R S,Ye J X,Yang A M. 2004. Spatial pattern and variance of the Chinese fir anthracnose Glomerella cingualta[J]. Chinese Journal of Eco-Agriculture,12(3):169-171.]
朱英芝,廖旺姣,邹东霞,吴耀军,邓艳. 2015. 广西油茶炭疽病病原菌鉴定及生物学特性[J]. 植物保护学报,42(3):382-389. [Zhu Y Z,Liao W J,Zou D X,Wu Y J,Deng Y. 2015. Identification and biological characteristics of pathogen from Camellia oleifera anthracnose in Guangxi[J]. Journal of Plant Protection,42(3):382-389.] doi:10.13802/j.cnki.zwbhxb.2015.03015.
Weir B S,Johnston P R,Damm U. 2012. The Colletotrichum gloeosporioides species complex[J]. Studies in Mycology,73:115-180. doi:10.3114/sim0011.
收稿日期:2021-09-06
基金项目:广西创新驱动发展专项(桂科AA17204087-10);广西林业科技项目(桂林科字〔2016〕第4号)
通讯作者:黄华艳(1974-),https://orcid.org/0000-0003-2436-862X,正高级工程师,主要从事林木病虫害防治研究工作,E-mail:258939303@qq.com
第一作者:廖旺姣(1980-),https://orcid.org/0000-0002-1378-1358,高级工程师,主要从事林木病害防治研究工作,E-mail:liaowangjiao@126.com
