江西省芝麻地方种质表型性状遗传多样分析
2022-07-14王郅琪孙建周红英颜小文梁俊超赵云燕饶月亮危文亮乐美旺
王郅琪 孙建 周红英 颜小文 梁俊超 赵云燕 饶月亮 危文亮 乐美旺




摘要:【目的】對江西省芝麻地方种质资源进行表型性状遗传多样分析,为江西芝麻种质资源的有效利用及开发提供理论参考。【方法】依托“第三次全国农作物种质资源普查与收集行动”从江西省39个县(市、区)收集到的132份芝麻地方种质为试验材料,对其23个表型性状(12个质量性状和11个数量性状)进行测定,并计算遗传多样性指数和变异系数。同时,对11个数量性状进行相关分析,并对供试种质进行聚类分析。【结果】23个表型性状的平均遗传多样性指数为1.2821,其中,12个质量性状的遗传多样性指数为0.3217~1.0307,平均0.7290;11个数量性状的变异系数为4.70%~43.58%,遗传多样性指数为1.6138~2.0627,平均1.8855。3种粒色类型芝麻种质中,以46份白芝麻种质表型性状的平均遗传多样性指数最高,为1.3602,74份黑芝麻种质和12份其他粒色(黄色、褐色、黄褐色、红褐色等)芝麻种质表型性状的平均遗传多样性指数分别为1.1782和1.0476。生育期与株高、始蒴高度、蒴果长、千粒重和蛋白含量呈极显著正相关(P<0.01,下同),但与含油量呈极显著负相关。供试的132份芝麻种质资源可分为四大类,不同类群种质的来源地存在地理位置的交错分布现象,其中,第Ⅰ类群可作为潜在选育芝麻高含油量新品种的亲本材料,第Ⅱ类群可作为潜在高产优良芝麻材料利用,第Ⅲ类群可作为潜在矮杆株型的亲本材料,第Ⅳ类群可作为潜在的高蛋白型亲本材料加以利用。【结论】江西芝麻地方种质资源的主要表型性状变异较大,遗传多样性较丰富,其中以46份白芝麻种质的遗传多样性更丰富。种质间亲缘关系与地理来源无必然联系。以高产高蛋白为育种目标时应选择株高较高,始蒴高度适中、蒴果较大、千粒重较高的材料为亲本,并据此进行后代选择。
关键词:芝麻;江西;地方种质资源;表型性状;遗传多样性;聚类分析
中图分类号:S565.302.4 文献标志码: A 文章编号:2095-1191(2022)04-1021-09
Genetic diversity analysis of phenotypic traits of sesame local germplasm in Jiangxi Province
WANG Zhi-qi1,2, SUN Jian1, ZHOU Hong-ying1, YAN Xiao-wen1, LIANG Jun-chao1,ZHAO Yun-yan1,2, RAO Yue-liang1, WEI Wen-liang2*, LE Mei-wang1*
(1 Crops Research Institute, Jiangxi Academy of Agricultural Sciences/Jiangxi Province Key Laboratory of Oil Crops Biology/Nanchang Branch of National Center of Oil Crops Improvement,Nanchang, Jiangxi 330200, China;
2 College of Agriculture, Yangtze University, Jingzhou, Hubei 434025, China)
Abstract:【Objective】To analyze phenotypic traits and the genetic diversity characteristics of local sesame germplasm resources in Jiangxi Province, so as to provide theoretical basis for effective utilization of local sesame germplasm in Jiangxi Province. 【Method】With support from the Third National Campaign of Crop Germplasm Census and Collection, 132 local sesame germplasms from 39 counties in Jiangxi Province were collected,which were used as test materials.Twenty-three phenotypic traits (12 quality traits and 11 quantitative traits) were determined and genetic diversity index and coefficient of variation were computed. At the same time, the 11 quantitative traits were analyzed and cluster analysis were performed. 【Result】The results showed that among 23 phenotypic traits,the genetic diversity index of 12 quality traits ranged from 0.3217 to 1.0307,with an average of 0.7290. Meanwhile,the genetic diversity index of 11 quantitative traitsranged from 1.6138 to 2.0627,with an average of 1.8855. Among the three seed color types of sesame germplasm, 46 white sesame germplasm had the highest average genetic diversity index of phenotypic traits, which was 1.3602, 74 black sesame germplasm and 12 other seed color (yellow, brown, yellow-brown, red-brown, etc.) sesame germplasm. The average genetic diversity indexes of phenotypic traits were 1.1782 and 1.0476. Correlation analysis showed that the growth period was significantly positively correlated with plant height,first capsule height,mean capsule length,1000-seed weight and protein content(P<0.01, the same below),but it was extremely significantly negatively correlated with oil content. 132 sesame germplasm resources could be divided into four groups,and different origins of germplasms were distributed interlaced.Group I could be used as a potential parent material for breeding new sesame varieties with high oil content, group II could be used as a potential high yield and excellent sesame material, group III could be used as a potential dwarf plant type parent material, and group IV could be used as a potential high protein type parent material. 【Conclusion】Phenotypic variation of the main phenotypic traits of Jiangxi sesame germplasms is large. Genetic diversity of the germplasms is high, especially for 46 white sesame germplasms. The genetic relationship between germplasms is not necessarily related to the origins. In the breeding with high yield and high protein as the goal,the plant height can be biased. When aiming to breed high production and high protein content, the materials with higher first capsule height,larger mean capsule and higher thousand-seed weight should be chosen as the parents,and the offspring should be selected according to obtain excellent germplasm.
Key words: sesame; Jiangxi; local germplasm resources; phenotypic traits; genetic diversity; cluster analysis
Foundation items: National Modern Agricultural Industry Technology System (CARS-14); The Third National Action of Crop Germplasm Survey and Collection (111721301354052138)
0 引言
【研究意义】芝麻(Sesamum indicum L.)为胡麻科胡麻属的特色油料作物,在我国已有2000多年种植历史(吕伟等,2017)。芝麻营养价值丰富,其种子含油量高达50%以上,芝麻油中的不饱和脂肪酸(油酸和亚油酸)含量超过80%,蛋白含量在20%左右,且富含芝麻素、维生素E等天然抗氧化类物质(Nakano et al.,2003;Moazzami and Kamal-Eldin,2006)。芝麻的环境适应性广泛,在亚洲、非洲等70多个国家均有种植。我国是世界芝麻主产国之一,总产和单产均位于世界前列(Zhang et al.,2018)。其中,江西省是我国四大芝麻主产区之一,种植历史悠久,全省各市均有种植,民间蕴含着丰富的地方种质资源,但由于种植结构的变化和育成品种的不断推广利用,江西芝麻地方种质资源面临急剧丢失的风险,亟待开展种质资源抢救性收集工作。种质资源鉴定和评价是种质利用的重要基础,因此,对江西省芝麻地方种质表型性状遗传多样分析进行分析,对丰富地方种质资源及提高芝麻育种效率具有重要意义。【前人研究进展】我国芝麻地方种质资源十分丰富,关于芝麻种质资源的研究主要集中在产量性状、品质性状、抗病性和耐渍性的鉴定。俞信英等(2015)基于27个数量性状和55个质量性状对浙江省沿海地区19份芝麻地方种质资源进行遗传多样性分析,结果发现27个数量性状的变异系数为5.6%~93.7%,55個质量性状的遗传多样性指数为0~1.55%,且19份种质资源的聚类结果与胚轴长度、株高、果轴长度、千粒重、单株产量等性状有关,而与地域来源无关。吕伟等(2019)对189份山西芝麻种质资源的18个表型性状进行遗传多样性及聚类分析,结果发现山西芝麻种质的表型性状存在明显的遗传变异,质量性状以主茎茸毛量的遗传多样性指数最高(1.15),数量性状以节间长度的遗传多样性指数最高(2.09)。杨学乐等(2019)采用主成分分析和聚类分析方法,基于12个质量性状和9个数量性状对收集的63份湖南省种质资源进行遗传多样性分析,结果发现湖南省芝麻种质资源具有丰富的遗传多样性;茎秆绒毛是遗传多样性指数最高的质量性状,而空稍尖长度是变异系数最高的数量性状。崔宏亮等(2020)对135个新疆芝麻品种的10个主要农艺性状进行分析,结果发现有7个农艺性状变异系数大于10%,说明新疆种质资源的资源类型和遗传变异较丰富。崔彦琴等(2020)采用聚类分析方法,基于31个表型性状对300份河北省芝麻种质资源进行遗传多样性分析,结果发现22个质量性状的遗传多样性指数为1.0986~5.6941,存在丰富的遗传变异;300份种质资源可聚为六大类群。杨航等(2020)对73份贵州地方芝麻种质资源的8个品质性状进行分析,结果发现贵州芝麻种质资源平均含油量约50%,不饱和脂肪酸含量在85%以上,其中亚油酸含量较高,具有开发利用价值。【本研究切入点】虽然江西省蕴含着丰富的地方种质资源,但鲜见有关江西省芝麻地方种质表型性状遗传多样分析的研究报道。【拟解决的关键问题】依托2017—2019年“全国第三次农作物种质资源普查与收集行动”共收集江西全省所有县市的芝麻地方种质132份,对其主要表型性状和不同粒色类型芝麻种质的遗传多样性进行分析,挖掘优异种质,为江西芝麻种质资源的有效利用提供可靠数据及理论参考。
1 材料与方法
1. 1 试验材料
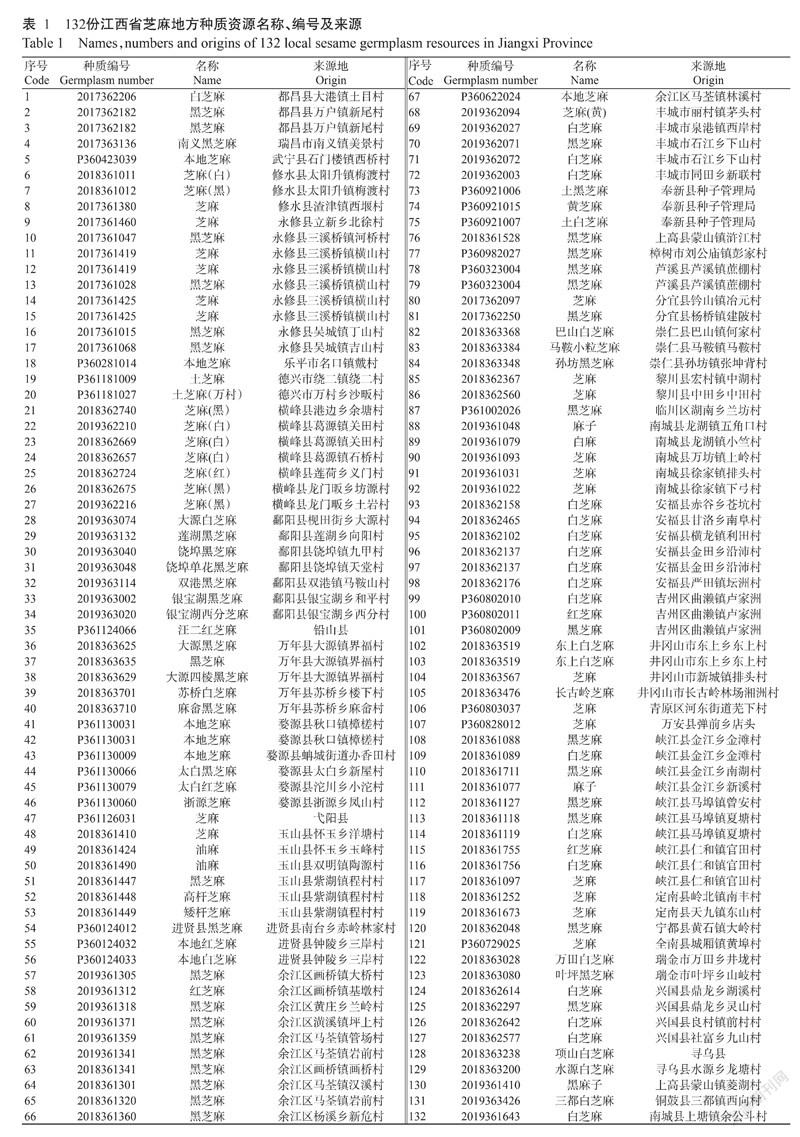
供试材料为“全国第三次农作物种质资源普查与收集行动”所收集的132份江西省芝麻地方种质资源,分别来自九江、南昌、上饶、景德镇、鹰潭、宜春、萍乡、新余、抚州、吉安、赣州等11个地市的39个县(市、区),其中,白芝麻46份,黑芝麻74份,黄色、褐色、黄褐色和红褐色等其他粒色芝麻共计12份,种质信息详见表1。
1. 2 试验设计
试验于2020年6—10月在江西省农业科学院作物研究所南昌横岗试验田基地进行。采取顺序区组设计,每份种质种植4行,行长1.6 m,行距0.4 m,株距0.15 m,进行日常田间栽培管理。
1. 3 试验方法
参考《芝麻种质资源描述规范化和数据标准化》(张秀荣和冯祥运,2006)的描述标准对23个表型性状进行观察记载和数据采集,包括12个质量性状(株型、基部叶缘、花旁蜜腺、每叶腋蒴果数、蒴果棱数、成熟主茎颜色、蒴果颜色、粒色、叶茸毛稀密和长短、茎秆绒毛稀密和长短),以及11个数量性状(生育期、株高、始蒴高度、空稍尖长度、蒴果长、蒴果宽、蒴果厚、每蒴粒数、千粒重、蛋白含量和含油量)。每份种质选取10个典型单株进行数据测定和采集。参照刘艳阳等(2014)的方法,利用近红外分析仪测定含油量和蛋白质含量。
1. 4 统计分析
遗传多样性指数计算采用Shannon-Weaver信息指数法(Keylock,2005),计算公式:H′=-ΣPi×lnPi ,其中Pi为某一性状第i级别内材料份数占总份数的百分比。对数量性状进行分级,先计算参试材料的总体平均数([X])和标准差(s),然后划分为10级,即第一级[Xi<([X]-2s)]到第十级[Xi>([X] +2s)],每0.5 s为一级,每一级的相对频率用于计算遗传多样性指数(董玉琛等,2003;任小平等,2007;孙建等,2009)。平均遗传多样性指数为各性状遗传多样性指数的平均值。利用Excel 2010进行数据整理分析、SPSS 19.0进行相关分析及聚类分析(孔凡洲等,2012)。
2 结果与分析
2. 1 芝麻种质资源表型性状的遗传多样性分析结果
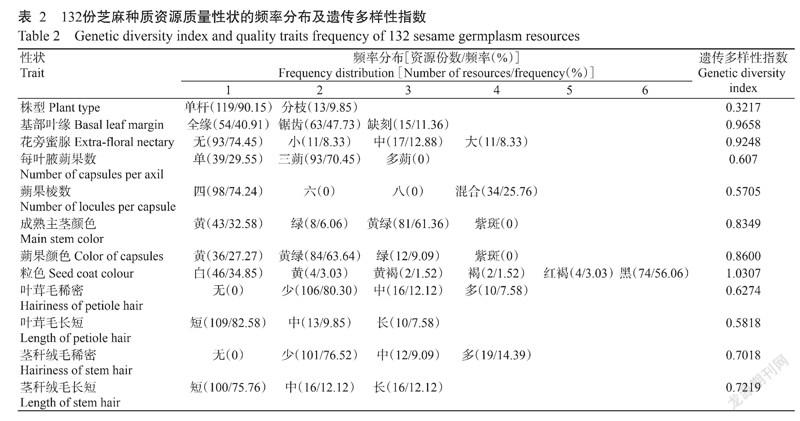
132份江西地方芝麻种质资源的12个质量性状的频率分布如表2所示。90.15%的种质株型为单杆;47.73%的种质基部叶缘为锯齿;74.45%的种质无花旁蜜腺;每叶腋蒴果数以三蒴居多,占比70.45%;74.24%的种质蒴果棱数为四棱;61.36%的种质成熟主茎颜色为黄绿色;蒴果颜色以黄绿色居多(63.64%),56.06%的种质粒色为黑色;对于叶茸毛稀密和长短,80.30%种质为少量茸毛,82.58%种质为短绒毛;对于茎秆绒毛稀密和长短,76.52%种质为少量绒毛,75.76%种质为短绒毛。12个质量性状的遗传多样性指数分析结果(表2)显示,12个质量性状的遗传多样性指数为0.3217~1.0307,平均0.7290,其中,粒色的遗传多样性指数最高(1.0307),株型的遗传多样性指数最低(0.3217)。
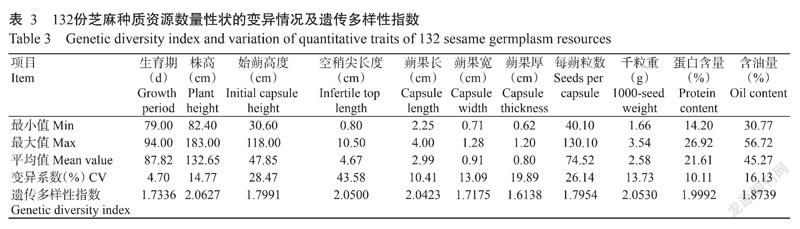
对132份芝麻种质资源11个数量性状的变异范围、变异系数和遗传多样性指数进行分析,结果(表3)显示,数量性状的变异系数为4.70%~43.58%,遗传变异较明显,其中,始蒴高度、空稍尖长度和每蒴粒数的变异系数较大,均在20%以上,说明这些性状的遗传稳定性较差;11个数量性状的遗传多样性指数为1.6138~2.0627,平均1.8855,其中,以株高的遗传多样性指数最高(2.0627),蒴果厚的遗传多样性指数最低(1.6138)。
综合来看,12个质量性状的平均遗传多样性指数为0.7290,11个数量性状的平均遗传多样性指数为1.8855。23个表型性状的平均遗传多样性指数为1.2821,表明132份江西省芝麻地方种质资源的表型性状遗传多样性较丰富,且数量性状遗传多样性比质量性状更丰富。
2. 2 不同粒色种质变异情况及遗传多样性分析结果
供试的132份芝麻种质资源粒色十分丰富,包括白、黑、黄、褐、黄褐、红褐色等多种类型,根据粒色可大体分为白芝麻、黑芝麻和其他粒色芝麻3种类型,其中,白芝麻种质46份,占34.85%;黑芝麻种质74份,占56.06%;其他粒色(黄色、红褐色、褐色、黄褐色等)芝麻种质12份,占9.09%。通过分析3种粒色類型芝麻种质11个数量性状的变异情况及遗传多样性指数(表4)可知,黑芝麻种质的生育期、空稍尖长度、蒴果长和蛋白含量4个性状的平均值均高于白芝麻种质及其他粒色芝麻种质,且其变异系数范围(3.41%~40.40%)最小,说明黑芝麻种质资源的数量性状间差异相对较小。白芝麻种质的蒴果宽度、蒴果厚度、每蒴粒数、千粒重和含油量的平均值均高于黑芝麻种质及其他粒色芝麻种质,各数量性状的遗传多样性指数(1.5996~2.1106)较白芝麻种质及其他粒色芝麻种质高,说明白芝麻种质数量性状的遗传多样性较丰富。其他粒色芝麻种质的株高和始蒴高度的变异系数范围(5.30%~57.78%)均大于白芝麻种质和黑芝麻种质,说明其他粒色芝麻种质间差异相对较大,可能与混合有不同粒色芝麻种质资源有关。
比较3种粒色类型芝麻种质23个表型性状的遗传多样性指数,结果发现46份白芝麻种质23个表型性状的平均遗传多样性指数为1.3602,74份黑芝麻种质的平均遗传多样性指数为1.1782,12份其他粒色芝麻种质的平均遗传多样性指数为1.0476,可见白芝麻种质表型性状的遗传多样性最丰富。
2. 3 芝麻种质资源数量性状的相关分析结果
对芝麻种质资源的11个数量性状进行相关分析,结果如表5所示。生育期与株高、始蒴高度、蒴果长、千粒重和蛋白含量呈极显著正相关(P<0.01,下同),但与含油量呈极显著负相关;株高与始蒴高度、蒴果长度、千粒重和蛋白含量呈极显著正相关,与含油量呈显著负相关(P<0.05,下同),其中始蒴高度与株高的相关性最大。始蒴高度和含油量呈显著负相关。空稍尖长度和蒴果长呈极显著正相关,与蒴果厚呈显著负相关。蒴果长与千粒重呈极显著正相关,与蒴果宽、蒴果厚和含油量呈极显著负相关。蒴果宽与蒴果厚、每蒴粒数和含油量呈极显著正相关,与千粒重呈显著负相关。蒴果厚度与每蒴粒数和含油量呈极显著正相关,与千粒重呈极显著负相关。每蒴粒数与千粒重呈极显著负相关。蛋白含量与含油量呈极显著负相关。
2. 4 芝麻种质资源的聚类分析结果
采用欧式距离Ward聚类法对132份江西省地方芝麻种质资源进行聚类分析,将其分为四大类(图1),分别计算各类群的数量性状,结果如表6所示。第I类群包含42份种质,占供试种质数量的31.82%,该类群芝麻种质的质量性状:株型均为单杆,基部叶缘多为锯齿,花旁蜜腺以无蜜腺为主,每叶腋蒴果数以三蒴居多,蒴果棱数均为混合棱数,成熟主茎颜色以黄色为主,蒴果颜色以黄色和黄绿色为主,叶茸毛稀密以少量茸毛居多,叶茸毛长短以短绒毛为主,茎秆绒毛稀密以少量绒毛居多,茎秆绒毛长短以短绒毛居多。该类群种质的数量性状:生育期最短,株高适中,始蒴高度适中,空稍尖长度适中,蒴果长最短,蒴果宽、蒴果厚、每蒴粒数和含油量均最高,千粒重最小,可作为潜在选育芝麻高含油量新品种的亲本材料。第II类群包括40份材料,占供试种质数量的30.30%。该类群芝麻种质中质量性状的特点表现为株型、花旁蜜腺、每叶腋蒴果数、叶茸毛稀密、叶茸毛长短、茎秆绒毛稀密、茎秆绒毛长短和每叶腋蒴果数8个性状与第I类群一致,而基部叶缘表现全缘型和锯齿型数量相当;蒴果棱数以四棱为主;成熟主茎颜色以黄绿为主;蒴果颜色中以黄绿色为主。该类群的数量性状:生育期适中,株高适中,始蒴部位较高,空稍尖长度较长,蒴果的长,宽,厚均较长,每蒴粒数适中,千粒重相对较高、蛋白含量和含油量均较高,该类种质整体属于潜在的高产型种质资源,综合性状表现良好,可作为潜在高产优良芝麻材料利用。第III类群包括17份种质,占供试种质数量的12.88%。该类群芝麻种质的质量性状与第II类群基本一致,其数量性状:株高最矮,始蒴高度最低,空稍尖长度最长,每蒴粒数最少,千粒重中等,蛋白含量和含油量适中,可作为潜在的矮秆株型亲本材料。第IV类群的材料包含33份种质,占供试种质数量的25.00%。该类种质的质量性状表现最为独特,其中,最特殊的是株型,分单杆和分枝两种,基部叶缘多为全缘和锯齿,花旁蜜腺多为中等和大型,每叶腋蒴果数多为单蒴,蒴果棱数均为四棱,其数量性状:生育期最长,株高最高,始蒴高度最高,空稍尖长度最短,蒴果长度最长,蒴果宽度和厚度最低,每蒴粒数适中,千粒重最高,蛋白含量最高,含油量最低,该类种质可作为潜在的高蛋白型的亲本材料加以利用。
3 讨论
不同来源芝麻种质资源的表型性状各具特色,是丰富芝麻优异种质资源多样性的关键,也是挖掘优异种质、选育新品种的基础。通过分析供试种质资源的变异系数和遗传多样性指数可评价种质资源的遗传多样性丰富程度。本研究对132份江西地方芝麻种质资源的23个表型性状进行遗传多样性分析,结果发现11个数量性状的遗传多样性指数(1.6138~2.0627)整体高于质量性状的遗传多样性指数(0.3217~1.0307)。该结论与韩俊梅等(2019)对200份山西芝麻种质资源的13个农艺性状遗传多样性指数分析结果一致,但遗传多样性指数最大值和最小值所对应的性状不同,可能是数据调查、种植区域、材料来源、气候土壤条件等方面的差异所导致,需进行进一步科学验证。从变异系数来看,本研究供试种质的11个数量性状变异系数为4.70%~43.58%,除生育期外,其余的10个数量性状的变异系数均大于10.00%,说明132份江西省芝麻地方种质的表型变异较大,遗传多样性较丰富,与吕伟等(2019)的研究结果基本一致。此外,本研究3种粒色类型芝麻种质23个表型性状的遗传多样性进行比较,结果发现3种芝麻粒色类型的平均遗传多样性指数排序为白芝麻种质(1.3602)>黑芝麻种质(1.1782)>其他粒色芝麻种质(1.0476),表明3种粒色类型中白芝麻种质的遗传多样性较黑芝麻种质和其他粒色芝麻种质的遗传多样性丰富,可能与所收集的其他粒色芝麻种质数量较少有关,也可能与不同粒色芝麻有无商品化生产及不同地区间种质是否频繁交换有关。黑芝麻是江西省种植面积较大的芝麻粒色类型,其种质间交流频繁,已形成商品化生产,导致黑芝麻种质趋向单一化。白芝麻种质的种植面积与黑芝麻相比略低,种质间的交流较少,导致白芝麻的品种类型丰富。
本研究对芝麻种质资源的11个数量性状进行相关分析,结果显示生育期与株高、始蒴高度、蒴果长、千粒重和蛋白含量呈极显著正相关,但与含油量呈极显著负相关。刘盼等(2016)也研究发现,不同环境中芝麻籽粒含油量与蛋白质含量之间呈极显著负相关。这些结论可为芝麻育种的亲本选择和后代材料选育提供理论参考,如以高产高蛋白为育种目标时应选择株高较高、始蒴高度适中、蒴果较大、千粒重较重的材料为亲本,并据此进行后代选择;在以高油为育种目标时应选择生育期或偏早,籽粒偏小的亲本材料。
本研究对供试的132份芝麻种质资源进行聚类分析,可将其分为四大类,根据各类群的特点可以看出,类群间的差异主要集中于数量性状的差异性,说明132份江西芝麻种质资源的数量性状相似程度较小、独特性突出,且数量性状的差异更易体现在聚类结果中,与王玮勇等(2013)对甜瓜种质的研究结果相反,说明数量性状或质量性状在不同作物中的聚类作用存在一定差异。此外,从四大类群种质的来源地来看,不同类群种质的来源地存在地理位置的交错分布现象,表明江西省芝麻种质间不存在明显的地域性类群,且其亲缘关系的远近也与地理来源无必然关系(卫文星等,1994;Dikshit and Swain,2000;张鹏等,2008)。
通过描述和鉴定农艺性状、计算遗传多样性指数和变异系数以及种质聚类分析对作物种质资源进行鉴定、评价和遗传多样性分析,一直是种质资源研究利用的重要手段(李润枝等,2009)。但由于表型性状的鉴定易受鉴定人员的主观判断、标准的选择以及环境因素等影响,导致基于种质资源群体的表型性状开展遗传多样性分析具有一定的局限性。随着生物技术的不断发展,从DNA水平上对种质资源进行遗传多样性评价,并结合表型性状进行综合分析,有助于更准确和可靠地明确其遗传基础、挖掘优异种质并应用于芝麻品种改良。
4 结论
江西芝麻地方种质资源的主要表型性状变异较大,遗传多样性较丰富,其中以46份白芝麻种质的遗传多样性更丰富。种质间亲缘关系远近与地理来源无必然联系。以高产高蛋白为育种目标时应选择株高较高,始蒴高度适中、蒴果较大、千粒重较高的材料为亲本,并据此进行后代选择。
参考文献:
崔宏亮,王莉,宋晓晓,姚庆,林啸,西尔娜依·阿布都拉,安万刚,张学超. 2020. 新疆芝麻种质资源主要农艺性状的相关性及聚类分析[J]. 分子植物育种,18(23):7914-7925. [Cui H L,Wang L,Song X X,Yao Q,Lin X,Xernayi·Abudula ,An W G,Zhang X C. 2020. The correlation and cluster analysis of main agronomic traits of sesame germplasm resources in Xinjiang[J]. Molecular Plant Breeding,18(23):7914-7925.]doi:10.13271/j.mpb.018.007914.
崔彥芹,郭元章,蹇家利,关中波,侯少峰,徐桂真. 2020. 河北省芝麻地方品种资源表型多样性分析[J]. 植物遗传资源学报, 21(1):224-233. [Cui Y Q,Guo Y Z,Jian J L,Guan Z B,Hou S F,Xu G Z. 2020. Phenotypic diversity analysis of sesame germplasm resources in Hebei Provi-nce[J]. Journal of Plant Genetic Resources,21(1):224-233.] doi:10.13430/j.cnki.jpgr.20191107003.
董玉琛,曹永生,张学勇,刘三才,王玉芬,游光霞,庞斌双,李立会,贾继增. 2003. 中国普通小麦初核心种质的产生[J]. 植物遗传资源学报,4(1):1-8. [Dong Y C,Cao Y S,Zhang X Y,Liu S C,Wang Y F,You G X,Pang B S,Li L H, Jia J Z.2003. Establishment of candidate core collections in Chinese common wheat germplasm[J]. Journal of Plant Genetic Resources,4(1):1-8.] doi:10.3969/j.issn.1672-1810.2003.01.002.
韩俊梅,吕伟,任果香,文飞,王若鹏,刘文萍. 2019. 200份芝麻种质资源农艺性状遗传多样性分析[J]. 江苏农业科学,47(13):95-99. [Han J M,Lü W,Ren G X,Wen F,Wang R P,Liu W P. 2019. Genetic diversity analysis of agronomic traits of 200 sesame germplasm resources[J]. Jiangsu Agricultural Sciences. 47(13):95-99.] doi:10.15889/j.issn.1002-1302.2019.13.024.
孔凡洲,于仁成,徐子钧,周名江. 2012. 应用Excel软件计算生物多样性指数[J]. 海洋科学,36(4):57-62. [Kong F Z,Yu R C,Xu Z J,Zhou M J. 2012. Application of Excel software to calculate biodiversity index[J]. Marine Scien-ce,36(4):57-62.]
李润枝,陈晨,张培培,王晨,胡大鹏. 2009. 我国燕麦种质资源与遗传育种研究进展[J]. 现代农业科技, (17):44-45.[Li R Z,Chen C,Zhang P P,Wang C,Hu D P. 2009. Advances in research on genetic resources and breeding and genetics of oat(Avena sativa L.)[J]. Modern Agricultural Technology,(17):44-45.]doi:10.3969/j.issn.1007-5739. 2009.17.028.
刘盼,张艳欣,黎冬华,王林海,高媛,周瑢,张秀荣,魏鑫. 2016.基于近红外模型的芝麻核心种质油脂和蛋白质含量变异分析[J]. 中国油料作物学报,38(6):722-729. [Liu P,Zhang Y X,Li D H,Wang L H,Gao Y,Zhou R,Zhang X R,Wei X. 2016. Oil and protein contents analysis of sesame core collection based on near infrared reflectance spectroscopy model[J]. Chinese Journal of Oil Crop Science,38(6):722-729.] doi:10.7505/j.issn.1007-9084.2016.06.003.
刘艳阳,汪红,芦海灵,梅鸿献,郑永战. 2014. 野生和栽培芝麻种子品质性状及近红外光谱分析[J]. 河南农业科学,43(5):44-47. [Liu Y Y,Wang H,Lu H L,Mei H X,Zheng Y Z. 2014. Quality characters and near infrared spectroscopy analysis of wild and cultivated species of sesame seed[J]. Journal of Henan Agricultural Sciences,43(5):44-47.] doi:10.3969/j.issn.1004-3268.2014.05.010.
吕伟,韩俊梅,任果香,文飞,王若鹏,刘文萍. 2019. 山西芝麻种质资源遗传多样性分析[J]. 作物杂志,(5):57-63. [Lü W,Han J M,Ren G X,Wen F,Wang R P,Liu W P. 2019. Genetic diversity analysis of sesame germplasm resources in Shanxi[J]. Crops,(5):57-63.] doi:10.16035/j.issn.1001-7283.2019.05.010.
吕伟,刘文萍,任果香,文飞,韩俊梅,王若鹏. 2017. 不同浓度生根粉对芝麻生长及产量的影响[J]. 作物杂志,(5):100-105. [Lü W,Liu W P,Ren G X,Wen F,Han J M,Wang R P. 2017. Effects of different concentrations of ABT on growth and yield of sesame[J]. Crops,(5):100-105.] doi:10.15889/j.issn.1002-1302.2019.12.024.
任小平,姜慧芳,廖伯壽,雷永,王圣玉,李栋. 2007. 龙生型花生的遗传多样性[J]. 武汉植物学研究,25(4):401-405. [Ren X P,Jiang H F,Liao B S,Lei Y,Wang S Y,Li D. 2007. Genetic diversity of Arachis hypogaea var. hirsute in peanut[J]. Journal of Wuhan Botanical Research,25(4):401-405.] doi:10.3969/j.issn.2095-0837.2007.04.014.
孙建,刘红艳,赵应忠,乐美旺,饶月亮,颜廷献,颜小文,周红英. 2009. 芝麻种质资源叶绿素含量的多样性分析[J]. 江西农业学报,21(12):5-9. [Sun J,Liu H Y,Zhao Y Z,Le M W,Rao Y L,Yan T X,Yan X W,Zhou H Y. 2009. Analysis of diversity in leaf chlorphyll content of sesame germplasm resources[J]. Acta Agriculturae Jiangxi,21(12):5-9.] doi:10.3969/j.issn.1001-8581.2009.12.002.
王炜勇,俞少华,李鲁峰,张飞,沈晓岚,俞信英,潘钢敏,楼春燕. 2013. 浙江省薄皮甜瓜地方品种的表型遗传多样性[J]. 植物遗传资源学报,14(3):448-454. [Wang W Y,Yu S H,Li L F,Zhang F,Shen X L,Yu X Y,Pan G M,Lou C Y. 2013. Phenotypic diversity analysis of Cucumis melo var. makuwa landraces of Zhejiang Province[J]. Journal of Plant Genetic Resource,14(3):448-454.] doi:10. 13430/j.cnki.jpgr.2013.03.023.
卫文星,张红,路风银,卫双玲. 1994. 芝麻品种主成分分析和遗传距离测定及其在杂交育种中的应用[J]. 华北农学报,9(3):29-33. [Wei W X,Zhang H,Lu F Y,Wei S L. 1994. Principal component analysis and genetic distance measurement of sesame varieties and their application in cross breeding[J]. Acta Agriculturae Boreali-Sinica,9(3):29-33.] doi:10.3321/j.issn:1000-7091.1994.03.007.
杨航,于二汝,魏忠芬,丰斌,李慧琳. 2020. 贵州地方芝麻种质资源品质性状的分析与评价[J]. 植物遗传资源学报,21(2):369-376. [Yang H,Yu E R,Wei Z F,Feng B,Li H L. 2020. Analysis and evaluation of quality-related traits in sesame germplasm resources in Guizhou[J]. Journal of Plant Genetic Resources,21(2):369-376.] doi:10.13430/j. cnki.jpgr.20191028005.
杨学乐,何录秋,张璐,李志清. 2019. 湖南芝麻种质资源农艺性状的遗传多样性[J]. 湖南农业大学学报(自然科学版),45(6):577-582. [Yang X L,He L Q,Zhang L,Li Z Q. 2019.Genetic diversity of main characters of sesame germplasm resources in Hunan[J]. Journal of Hunan Agricultural University (Natural Science Edition),45(6):577-582.] doi:10.13331/j.cnki.jhau.2019.06.003.
俞信英,沈晓岚,李鲁峰,王炜勇,俞少华,郁永明. 2015. 浙江省沿海地区芝麻地方品种的表型遗传多样性[J]. 浙江农业科学,56(5):748-751. [Yu X Y,Shen X L,Li L F,Wang W Y,Yu S H,Yu Y M. 2015. Phenotypic genetic diversity of sesame landraces in coastal areas of Zhejiang [J].Journal of Zhejiang Agricultural Sciences,56(5):748-751. ] doi:10.16178/j.issn.0528-9017.20150562.
张鹏,张海洋,郑永战,郭旺珍,魏利斌,张天真. 2008. 芝麻种质资源因子分析及聚类分析[J]. 中国油料作物学报,30(1):71-78. [Zhang P,Zhang H Y,Zheng Y Z,Guo W Z,Wei L B,Zhang T Z. 2008. Factor and cluster analysis of sesame (Sesame indicum) germplasm resources[J]. Chinese Journal of Oil Crop Sciences,30(1):71-78.] doi:10.3321/j.issn:1007-9084.2008.01.012.
张秀荣,冯祥运. 2006. 芝麻種质资源描述规范和数据标准[M]. 北京:中国农业出版社. [Zhang X R,Feng X Y. 2006. Descriptors and data standard for sesame(Sesame indicum L.)[M]. Beijing:China Agricultural Press.]
Dikshit U N,Swain D. 2000. Genetic divergence and heterosis in sesame[J]. Indian Journal of Genetics & Plant Bree-ding,60(2):213-219. doi:https://worldveg.tind.io/record/28932.
Keylock C. 2005. Simpson diversity and the Shannon–Wiener index as special cases of a generalized entropy[J]. Oikos,109:203-207. doi:10.1111/j.0030-1299.2005.13735.x.
Moazzami A A,Kamal-Eldin A. 2006. Sesame seed is a rich source of dietary lignans[J]. Journal of the American Oil Chemists Society,83(8):719-723. doi:10.1007/s11746-006-5029-7.
Nakano D,Itoh C,Ishii F,Kawanishi H,Takaoka M,Kiso Y,Tsuruoka N,Tanaka T,Matsumura Y. 2003. Effects of sesamin on aortic oxidative stress and endothelial dysfunction in deoxycorticosterone acetate—salt hypertensiverats[J]. Biological and Pharmaceutical Bulletin,26(12):1701-1705 . doi:10.1248/bpb.26.1701.
Zhang Y,Wang L,Gao Y,Li D,Yu J,Zhou R,Zhang X. 2018. Genetic dissection and fine mapping of a novel dt gene associated with determinate growth habit in sesame[J]. BMC Genetics,19(1):19-24. doi:10.1186/s12863-018-0614-y.
收稿日期:2021-05-06
基金项目:国家现代农业产业技术体系项目(CARS-14);第三次全国农作物种质资源普查与收集行动项目(111721301354052138)
通讯作者:危文亮(1972-),https://orcid.org/0000-0002-5350-9930,博士,教授,主要从事作物遗传育种研究工作,E-mail:whwenliang@163.com; 乐美旺(1963-),https//:orcid.org/0000-0001-5390-0932,博士,研究员,主要从事芝麻良种繁育与遗传育种研究工作,E-mail:mwyuecarl@163.com
第一作者:王郅琪(1994,https://orcid.org/0000-0002-3121-4891,研究方向为芝麻种质资源,E-mail:2609757974@qq.com
