荒漠植物珍珠猪毛菜根际土壤细菌多样性与土壤理化性质相关性分析
2022-07-13魏艳晨陈吉祥王永刚孟彤彤韩亚龙李美
魏艳晨, 陈吉祥*, 王永刚, 孟彤彤, 韩亚龙, 李美
(1.兰州理工大学石油化工学院,兰州 730050;2.兰州理工大学生命科学与工程学院,兰州 730050;3.兰州理工大学能源与动力工程学院,兰州 730050)
西北荒漠区作为我国典型的中-重型生态脆弱区,是气候变化响应的敏感带,也是西北生态安全的重要屏障。景泰位于甘肃中部,河西走廊东端,地处甘肃、内蒙、宁夏交界处,属于黄土高原与腾格里沙漠过渡地带。气温多变、干旱多风少雨、沙化严重,是典型的干旱生态脆弱区[1]。珍珠猪毛菜(Salsola passerina)属藜科半灌木,广泛分布于景泰县干旱荒漠区砾石质、沙砾质戈壁和盐碱湖盆地,具有抗风沙、耐旱和耐寒的特性,是维护区域生态平衡的重要植物。研究荒漠区土壤细菌多样性对荒漠植物生态适应及生态系统修复有重要意义。不同荒漠区优势植物类群根际土壤的微生物类群及丰度有明显差异。康宝天等[2]研究祁连山高寒草甸和荒漠草原土壤细菌群落组成发现,不同草地土壤的细菌群落存在显著差异,放线菌门和绿弯菌门在高寒草甸土壤中的相对丰度显著低于荒漠草原,而变形菌门、酸杆菌门、拟杆菌门和Rokubacteria则相反。黄耀龙[3]研究发现,荒漠植物梭梭(Haloxylon ammodendron)与白刺(Nitraria tangutorum)根际土壤的细菌多样性差异显著。单立山等[4]和李善家等[5]对联生珍珠猪毛菜和联生红砂、单生珍珠猪毛菜和单生红砂的比根长、比表面积、根系生物量的垂直分布特征及叶片功能性状进行比较,验证了红砂-珍珠猪毛菜联生群落适应干旱胁迫环境。然而,在荒漠环境胁迫下,单生珍珠猪毛菜根际土壤和荒漠土壤的细菌在数量和多样性均可能具有差异,且二者土壤理化性状也可能存在差异。为此,本研究选取珍珠猪毛菜根际土壤和荒漠土壤(周边非根际土壤)为研究对象,采用高通量测序对珍珠猪毛菜根际和周边非根际土壤的细菌群落结构进行研究;通过对比根际和非根际土壤的可培养细菌数量、细菌总数、理化性质、微生物群落结构的变化和区别,分析土壤细菌群落组成和土壤理化性质的关系,为珍珠猪毛菜生态适应及荒漠草原生态修复提供理论基础。
1 材料与方法
1.1 研究区概况及样品采集
研究区地处甘肃省景泰县境内荒漠草原区(E 103°55′,N 36°55′),属温带大陆性干旱气候,海拔1 276~3 321 m,年均气温8.2℃,年均降水量185 mm,年蒸发量达3 038.5 mm,无霜期191 d。该区主要地貌为风沙地貌,土壤以荒漠土和灰钙土为主。植被结构简单且稀疏,主要有珍珠猪毛菜、红砂,其中珍珠猪毛菜分布较为广泛。
在研究区内取珍珠猪毛菜根际土壤(rizosphere soil,GJ)和非根际土壤(non-rizosphere soil,NG)。其中,GJ:选取长势一致的珍珠猪毛菜3株做重复,随机选取离植株茎秆2~5 cm处的3个采样点,用土钻法取样,垂直深度0—60 cm,将所取土样混匀后装入无菌保鲜袋于低温保存待测;NG:自同一采样点周边区域,去除地表0—5 cm处土壤,收集5—60 cm土壤装于无菌保鲜袋,于冷藏条件带回实验室进行分析。
1.2 土壤理化性质的测定
土壤pH采用pH计测定;土壤含水率(water content,WC)用烘干法测定;土壤全氮(total nitrogen,TN)用凯氏定氮法测定;土壤速效氮(available nitrogen,AN)采用碱解扩散法测定;土壤全磷(total phosphorus,TP)采用碱熔融-钼锑抗比色法测定;土壤速效磷(available phosphorus,AP)采用NaHCO3浸提-钼锑抗比色法测定;土壤全钾(total potassium,TK)采用氢氟酸消解-原子吸收分光光度计法测定;土壤速效钾(available potassium,AK)采用NH4Ac浸提-火焰光度计法测定[6]。
1.3 土壤细菌总数和可培养细菌数的测定
土壤细菌总数测定采用“LIVE/DEAD BacLight Bacterial Viability Kit”试剂盒,根据说明书进行。称取10 g土样于90 mL灭菌生理盐水30℃振荡30 min,使土壤中细菌均匀分散,制成10倍稀释度的土壤悬液,离心收集菌体,用生理盐水重悬。SYTO9和碘化丙啶核酸染色剂染色后,用激光共聚焦显微镜观察[7]。可培养细菌数采用稀释涂布平板法[8]测定。土壤悬液依次进行 10-1、10-2、10-3、10-4稀释。取不同稀释度土壤悬液100 μL,涂布LB平板,每个稀释度设置3个平行。
1.4 土壤细菌多样性分析
采用HiPure Soil DNA Kits试剂盒(D3142,广州美基生物科技有限公司)提取土壤样本的DNA进行PCR扩增,扩增区域为细菌16S rDNA的V3-V4区,引物为341F(5′-CCTACGGGNGGCWGCAG-3′)和806R(5′-GGACTACHVGGGTATCTAAT-3′)。使用AMPure XP Beads对扩增产物进行纯化,用ABI StepOnePlus Real Time PCR System(Life Technologies,产地美国)进行定量,最后根据Novaseq 6000的PE250模式pooling上机测序[9]。
1.5 数据处理
采用Uparse软件对所有样品的全部有效Tags序列聚类,以97%相似水平的OTUs序列进行分类分析。利用SPSS 22.0软件进行统计分析。采用Image J软件完成CLSM数据统计分析。
2 结果与分析
2.1 土壤理化性质分析
珍珠猪毛菜根际土壤和非根际土壤的理化性状存在显著差异(表1)。其中,珍珠猪毛菜根际土壤的全氮、全钾、速效氮、速效磷和速效钾均显著高于非根际土壤;土壤含水量和pH显著低于非根际土壤;土壤全磷含量在根际土壤和非根际土壤间差异不显著。
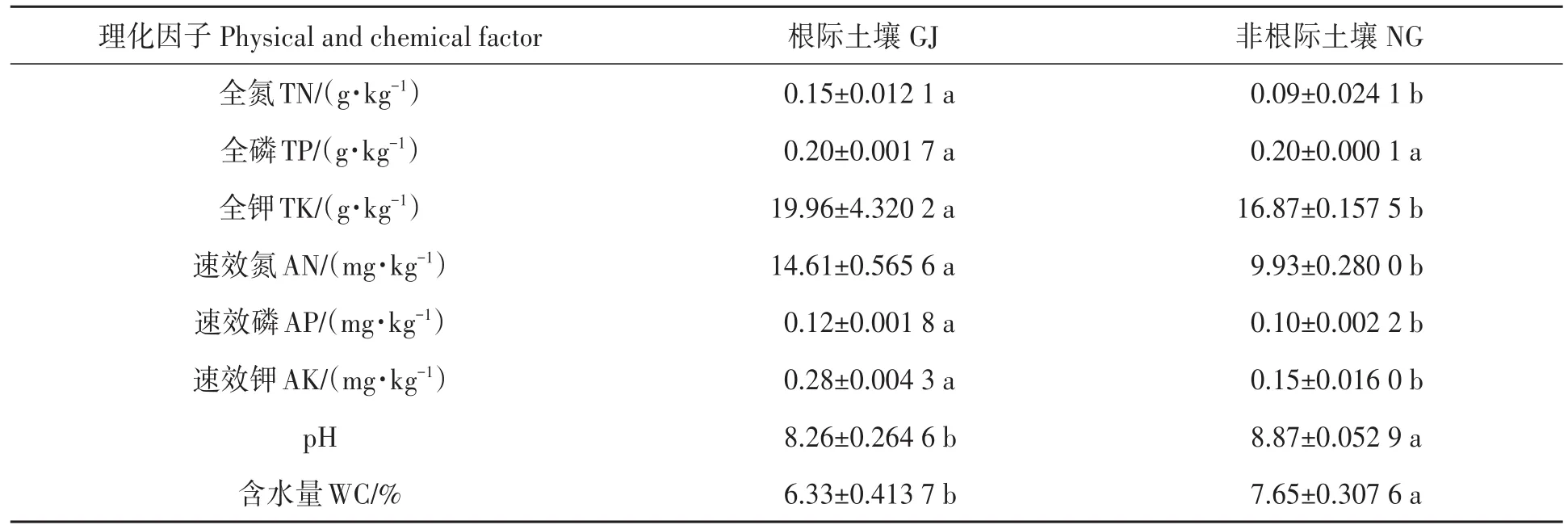
表1 珍珠猪毛菜根际和非根际土壤的理化性质Table 1 Physicochemical properties of GJ and NG
2.2 土壤细菌总数和可培养细菌数
对不同土壤的可培养细菌数、活细菌数和细菌总数的测定结果(图1)表明,根际和非根际土壤可培养细菌数分别为1.62×106CFU·g-1和5.40×105CFU·g-1;活细胞数分别为 2.52×106个·g-1和2.26×106个·g-1;细菌总数分别为1.33×107个·g-1和1.12×107个·g-1。由此表明,珍珠猪毛菜根际土壤的可培养细菌数、活细菌数和总细菌数均高于周边非根际土壤;且两种土壤样品的细菌总数和活细菌数均高于可培养细菌数。
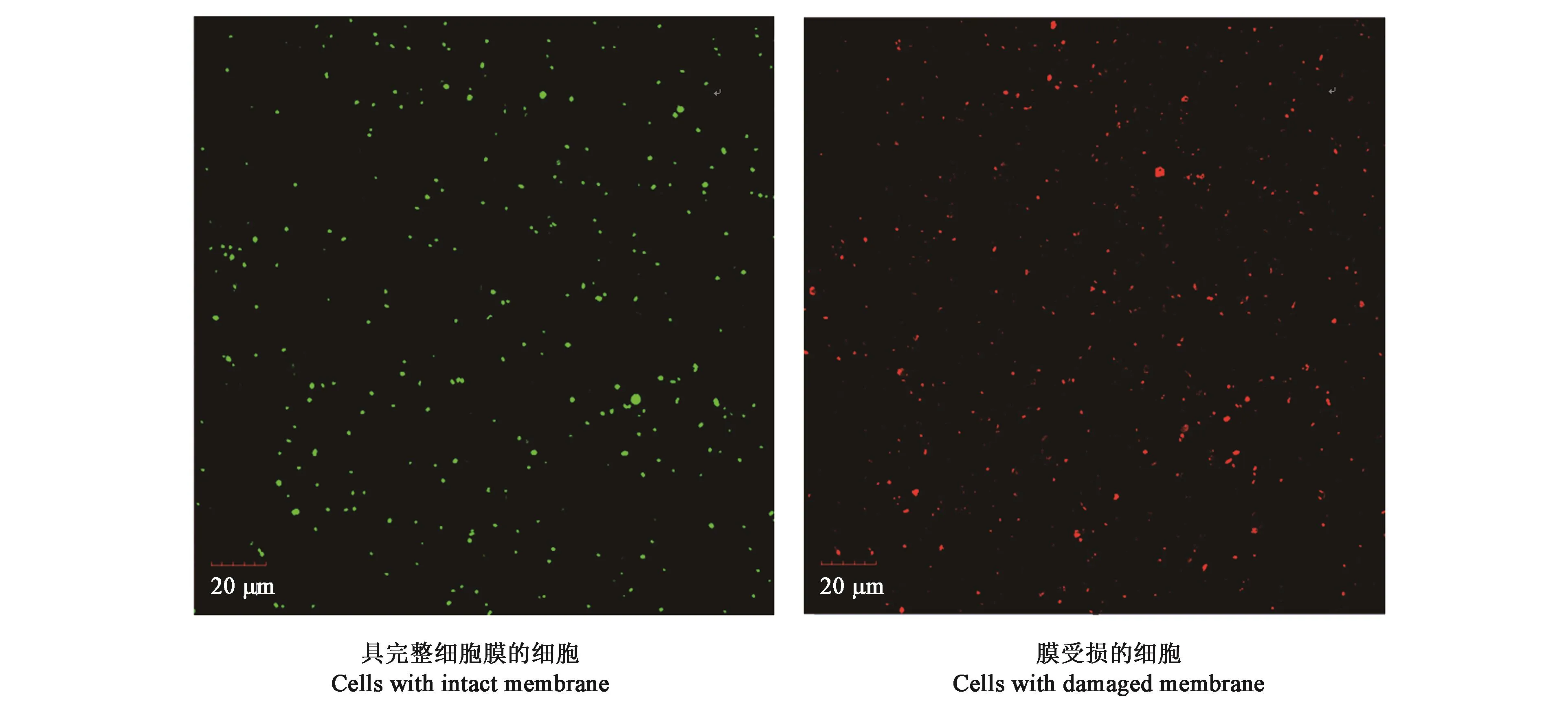
图1 激光共聚焦显微镜下染色细胞图Fig.1 Image of the stained cells with fluorescent dye under confocal laser microscope
2.3 土壤细菌群落多样性分析
通过测序,珍珠猪毛菜根际和非根际土壤中OUT数分别为5 655和4 665个。其中,根际和非根际土壤特有的OUT数分别为2 580和1 590个,共同的为3 075个。多样性分析结果(表2)表明,两种土壤中细菌的Chao1指数无显著差异;而珍珠猪毛菜根际土壤的Shannon指数显著高于非根际土壤。非度量多维尺度(nonmetric multidimensional scaling,NMDS)分析(图2)表明,根际和非根际土壤样本被分离,表明两种土壤的细菌群落组成具有明显差异。
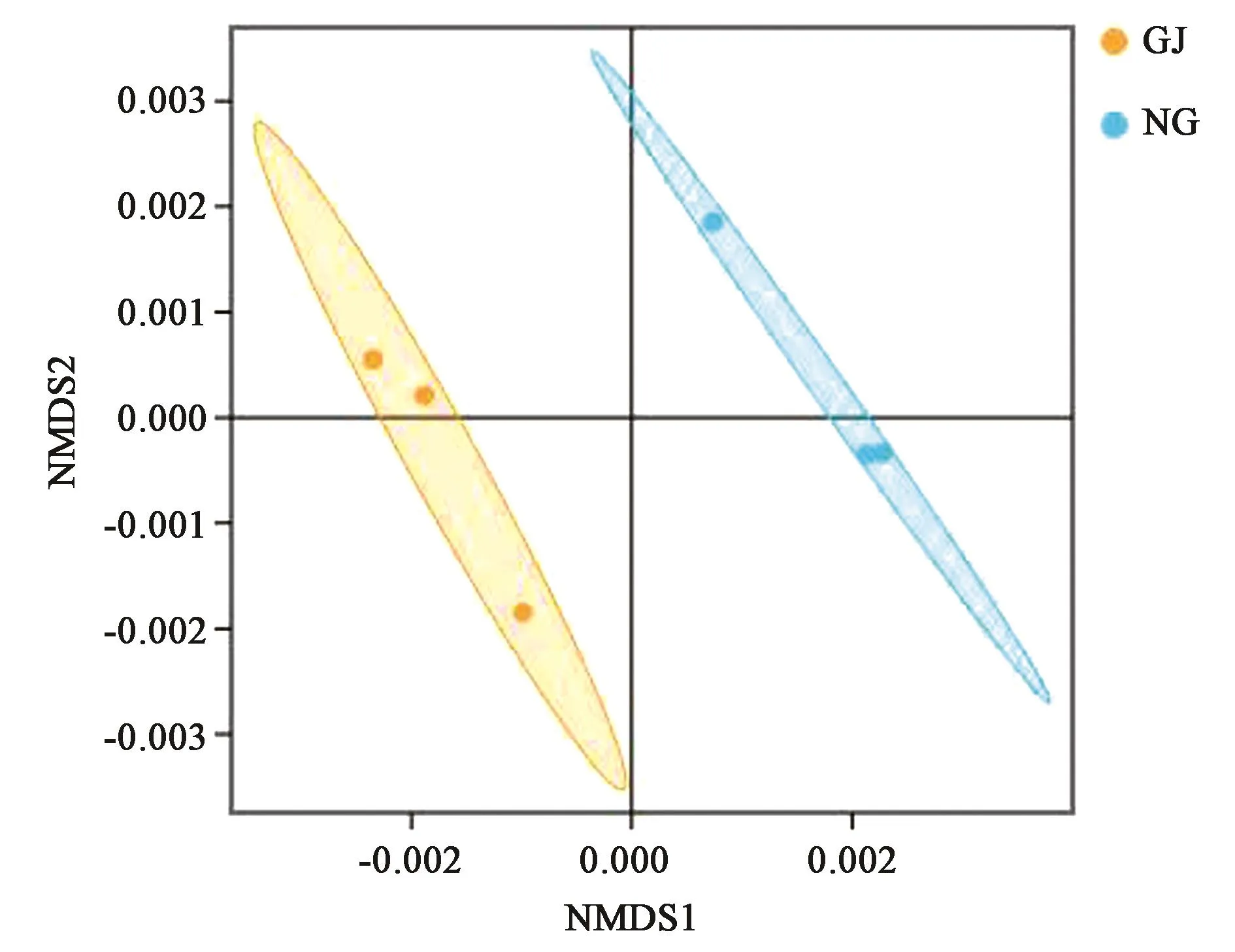
图2 基于Weighted UniFrace NMDS分析的样本二维排序图Fig.2 NMDS plot based on weighted UniFrac distances

表2 土壤细菌的多样性指数Table 2 Diversity indices of soil bacteria
2.4 不同土壤中细菌门组成
珍珠猪毛菜根际土壤和非根际土壤样品中共检测出33门细菌(图3)。其中,放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、浮霉菌门(Planctomycetes)、芽单胞菌门(Gemmatimonadetes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidetes)等在2种土壤样本中均有分布。放线菌门相对丰度最大,成为最优势菌门,在根际土壤中约占40.6%~43.3%,在非根际土壤中约占46.5%~52.0%。变形菌门、浮霉菌门和芽单胞菌门的相对丰度也较高。其中,浮霉菌门在根际土壤中的相对丰度(15.3%)显著高于非根际土壤(9.1%)。而WPS-2和Latescibacteria为珍珠猪毛菜根际土壤特有的2个菌门;软壁菌门(Tenericutes)和Halanaerobiaeota为非根际土壤特有的菌门。
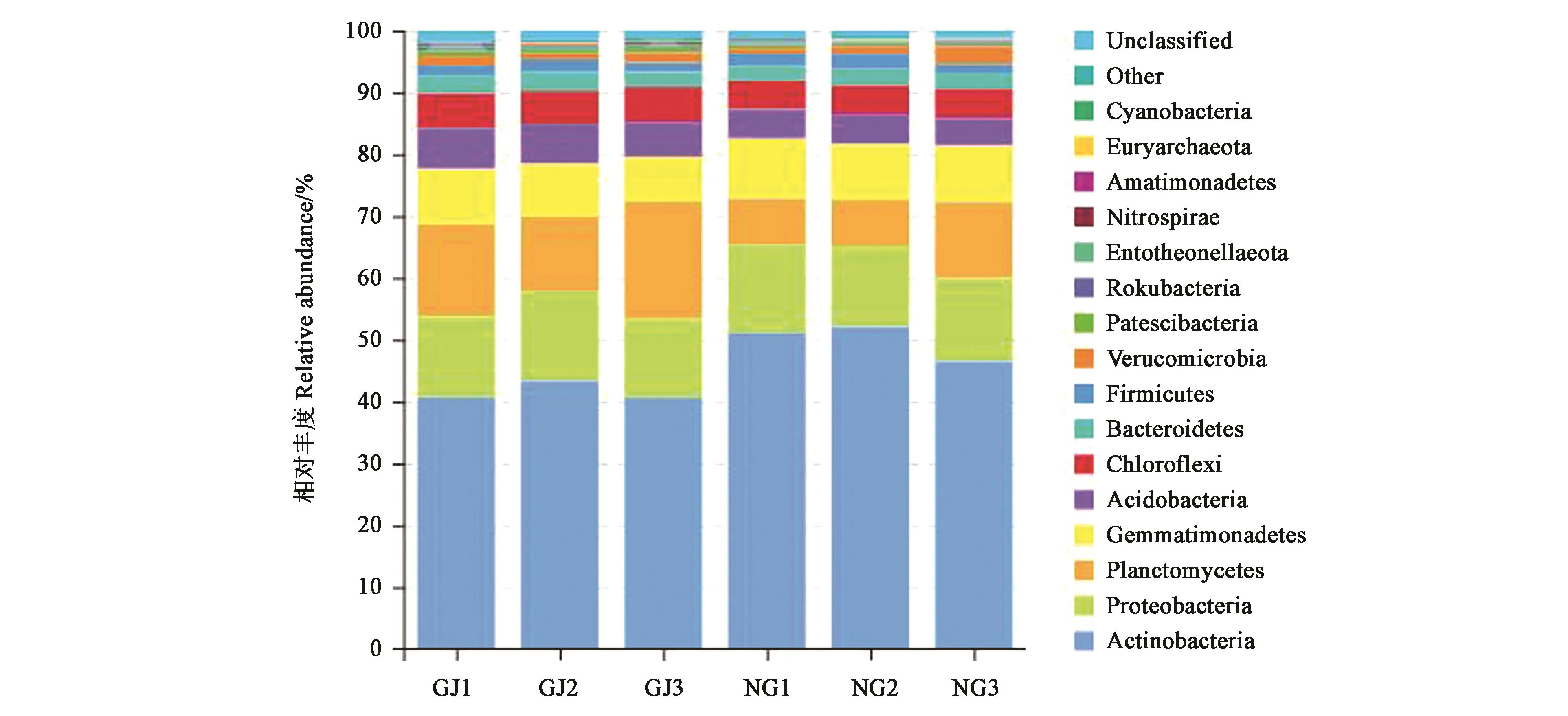
图3 珍珠猪毛菜根际和非根际土壤细菌门水平的相对丰度Fig.3 Relative abundance of bacterial communities at the phylum level in different soils of Salsola passerine
2.5 不同土壤中细菌属组成
珍珠猪毛菜根际土壤和非根际土壤分别包含367和398个细菌属。其中,红色杆菌属(Rubrobacter) 、RB41、类 诺 卡 氏 菌 属(Nocardioides)、链霉 菌 属(Streptomyces)、Pir4_lineage、芽 孢 杆 菌 属(Bacillus)、红 杆 菌 属(Solirubrobacter)、芽单胞菌属(Gemmatimonas)、小梨形菌属(Pirellula)假诺卡氏菌属(Pseudonocardia)、Subgroup_10、分 支 杆 菌 属(Mycobacterium)、半角藻属(Haliangium)、MND1、Gaiella和Pelagibius在2种土壤中的相对丰度均较高(图4)。其中,根际土壤相对丰度最高的优势菌属为红色杆菌属,占3.8%;而非根际土壤的主要优势菌属为类诺卡氏菌属(Nocardioides),占2.9%。芽球菌属(Blastococcus)和鞘氨醇单胞菌属(Sphingomonas)在根际土壤中的相对丰度也较高(>1%);而拟无枝酸菌属(Amycolatopsis)和糖霉菌属(Glycomyces)在非根际土壤中的相对丰度较高(>1%)。
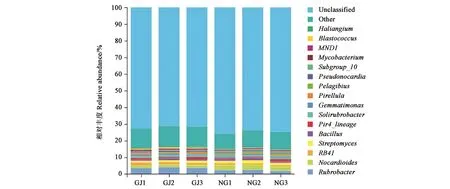
图4 珍珠猪毛菜根际和非根际土壤细菌属水平的相对丰度Fig.4 Relative abundance of bacterial communities at the genus level in different soils of Salsola passerine
2.6 土壤细菌与土壤理化性状间的关系
2.6.1 土壤细菌数量与理化因子相关性分析对土壤细菌数量与土壤理化性状进行相关性分析,结果(表3)表明,可培养细菌数量与土壤pH呈极显著负相关,与其他理化性状相关不显著;而细菌总数与土壤养分含量、pH和土壤含水率相关均不显著。
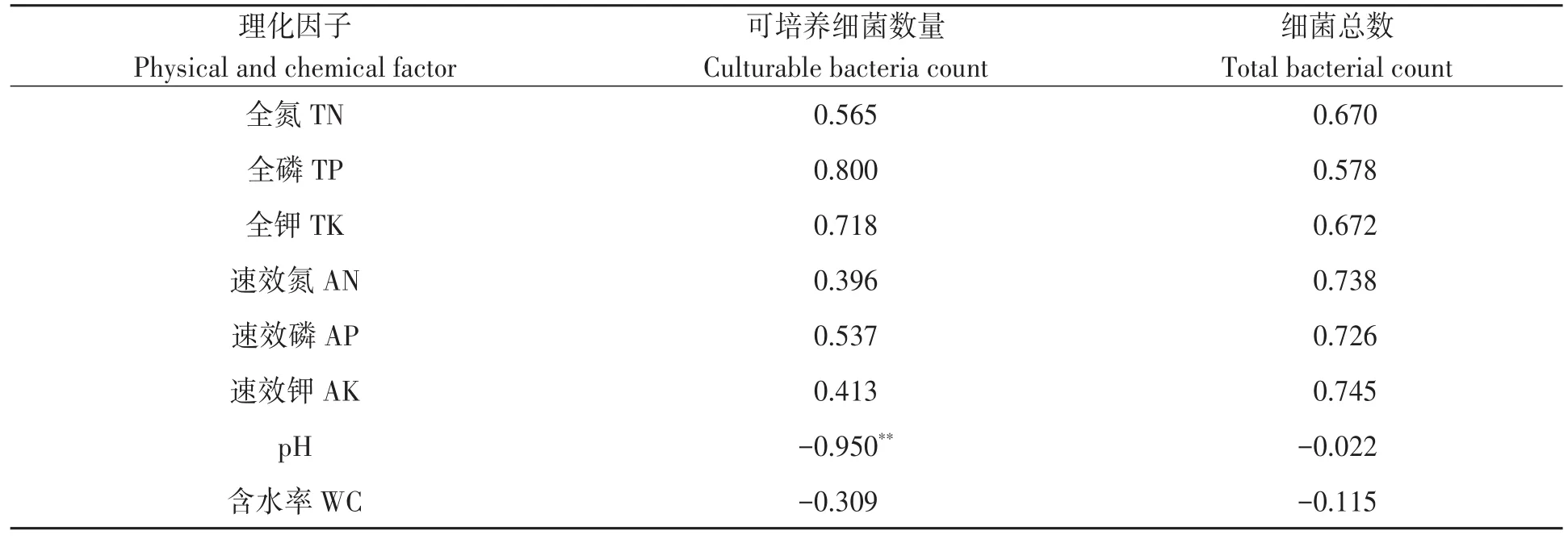
表3 土壤可培养细菌和细菌总数与理化因子之间的相关性Table 3 Correlation between the number of bacteria and soil physical and chemical properties
2.6.2 细菌多样性指数与土壤理化因子相关性分析 对细菌多样性与土壤理化性状进行相关分析,结果(表4)表明,Shannon指数和Simpson指数与土壤速效氮、速效磷和速效钾含量均达到极显著正相关;Simpson指数与土壤全氮含量呈显著正相关;ACE指数和Chao l指数均与土壤速效钾含量呈显著正相关。
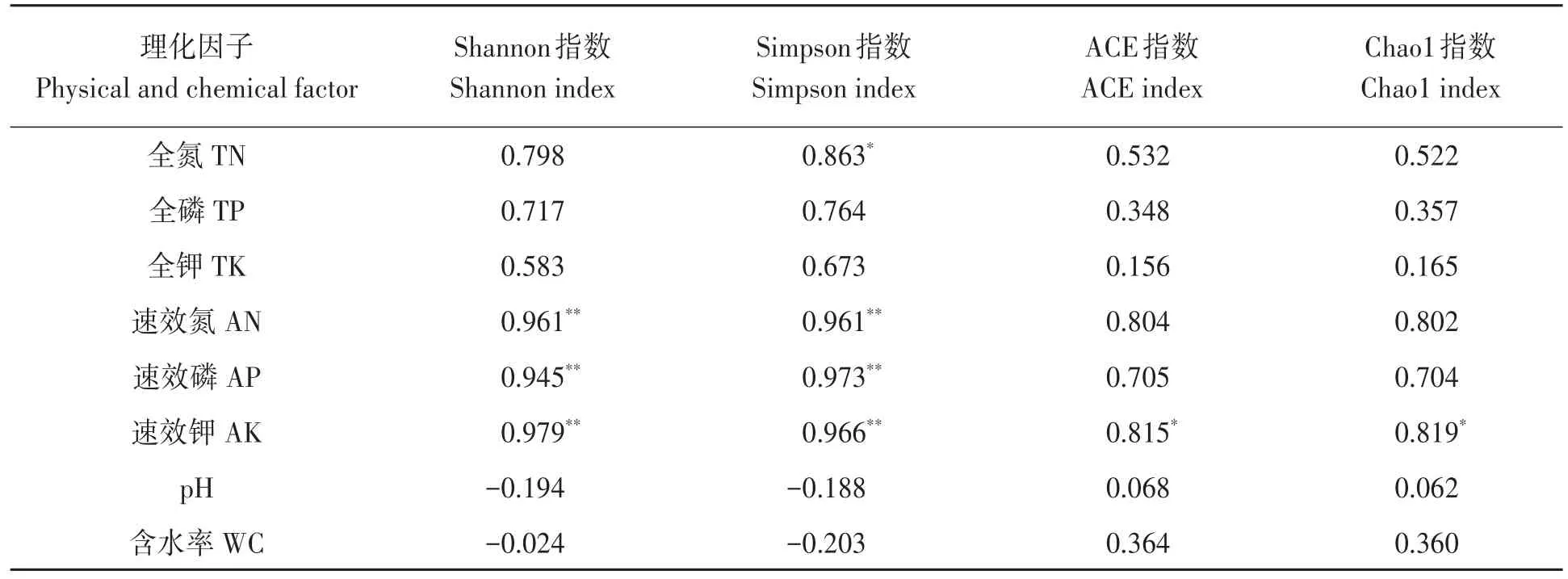
表4 细菌多样性与土壤理化因子之间的相关性Table 4 Correlation between physical and chemical properties and diversity index of bacteria
2.6.3 细菌优势菌门与土壤理化因子相关性分析 对细菌优势菌门与土壤理化性状进行相关性分析,结果(表5)表明,放线菌门的相对丰度与土壤全磷、速效氮、速效钾含量呈显著负相关,与土壤pH呈极显著正相关;酸杆菌门与土壤全氮、速效氮、速效钾含量呈显著正相关,与土壤pH和含水率呈显著负相关;绿弯菌门与土壤全氮和全磷含量呈显著正相关,与土壤含水率呈显著负相关,与速效氮和速效钾含量呈极显著正相关,与pH呈极显著负相关。
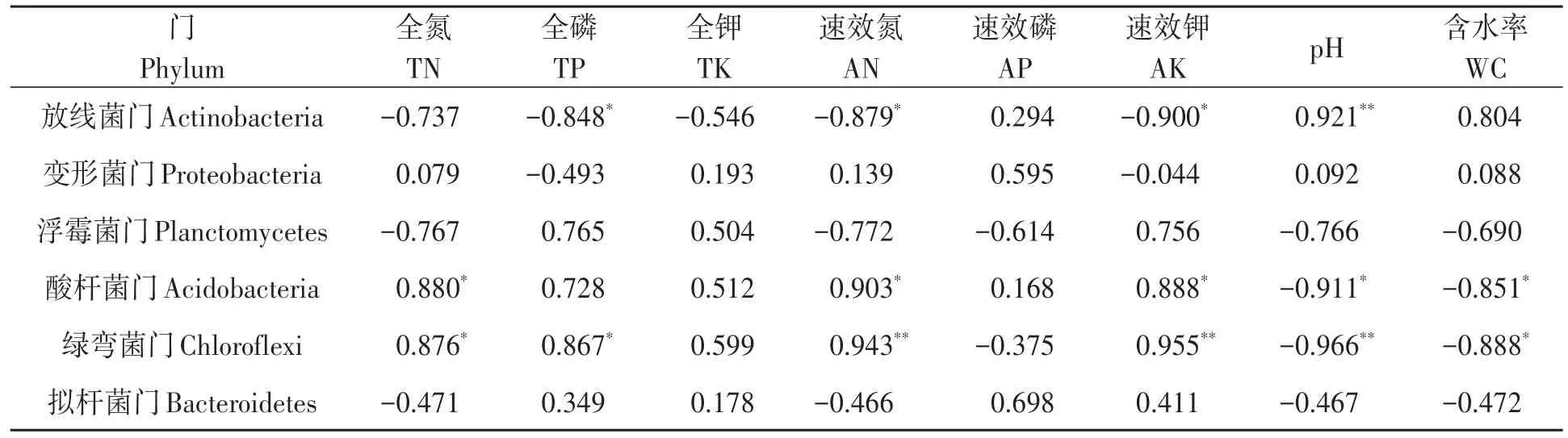
表5 不同土壤细菌优势菌门与土壤理化因子的相关性Table 5 Correlation of dominant communities of bacteria at the phylum level with soil physical and chemical factors
2.6.4 细菌优势菌属与土壤理化因子相关性分析 对细菌优势菌属与土壤理化性状进行相关性分析,结果(表6)表明,细菌优势菌属与不同土壤理化性状具有一定的相关性。链霉菌属与土壤pH呈显著正相关。红色杆菌属与土壤全氮含量呈显著正相关,与速效氮和速效钾含量呈极显著正相关,与土壤pH和含水率呈极显著负相关。RB41与土壤速效氮和速效钾含量呈显著正相关,与土壤pH呈显著负相关。类诺卡氏菌属和Pelagibius与土壤全氮含量呈显著负相关,与土壤含水率呈显著正相关,与速效氮和速效钾含量呈极显著负相关,与土壤pH呈极显著正相关。Pelagibius与土壤全磷含量呈显著负相关。由此表明,土壤养分含量、pH和含水量是影响土壤细菌群落组成的重要因子。
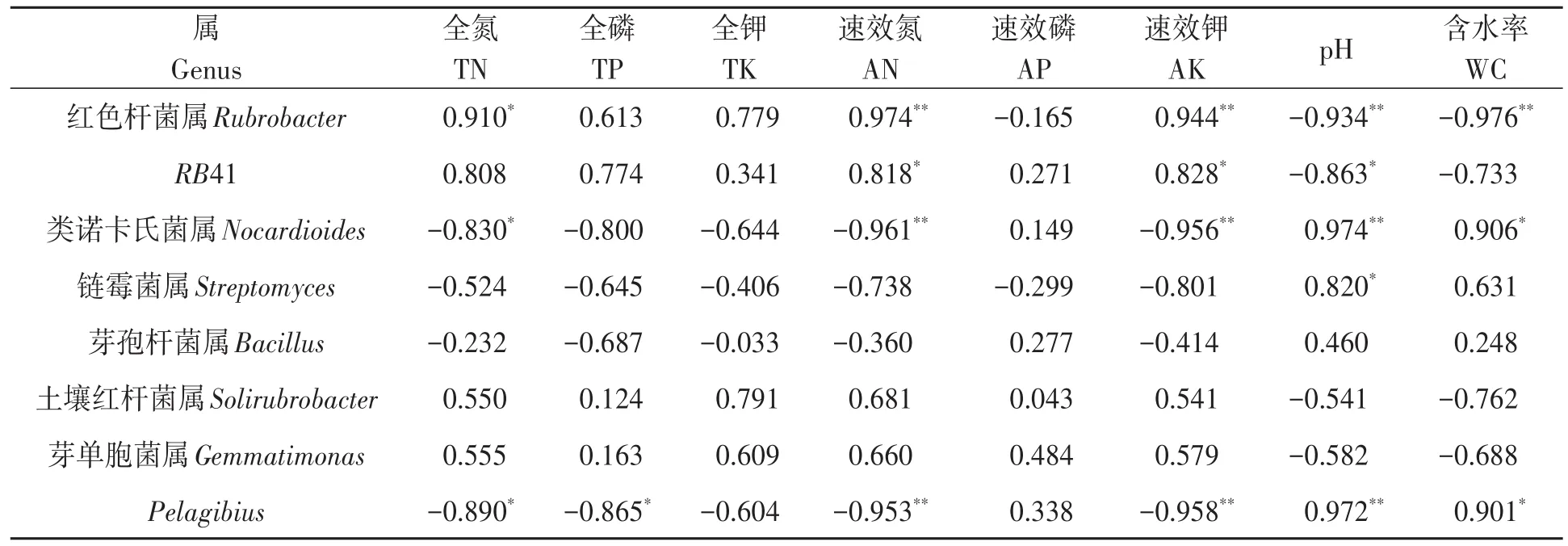
表6 不同土壤细菌优势菌属与土壤理化因子的相关性Table 6 Correlation of dominant communities of bacteria at the genus level with soil physical and chemical factors
3 讨论
景泰位于黄土高原与腾格里沙漠过渡地带,为典型荒漠草原,土壤pH 8.23~8.93。研究表明,荒漠土壤pH显著高于珍珠猪毛菜根际土壤,与杨阳等[10]研究结果相一致,可能是由于植物呼吸作用产生CO2及代谢作用产生的有机酸降低了土壤pH[11];而根际土壤含水率低于非根际土壤,可能与植物吸收水分有关。除全磷含量外,珍珠猪毛菜根际土壤养分含量均显著高于非根际土壤,说明植物根系从环境摄取养分并促进了根际养分的积累,体现了“根际效应”有利于增加土壤养分[12]。根际微生物为植物提供养料和水分,增强植物对氮素摄入,促进土壤磷富集,有利于植物生长[13-14]。
珍珠猪毛菜根际土壤可培养细菌数为1.62×106CFU·g-1,活细胞数为 2.52×106个·g-1,细菌总数为1.33×107个·g-1;与非根际土壤相比,都有不同程度增加,表明根际土壤有利于微生物活动。值得注意的是,不同土壤的可培养细菌数量比活细菌数和细菌总数低,可能是在营养贫瘠的环境中存在着较多难培养细菌或不可培养细菌(Viable but Non-Culturable)[15-16],由此可见,纯培养方法可能不能全面反映土壤微生物的类群和数量。
景泰荒漠草原土壤细菌具有丰富的多样性,其中,荒漠土壤中细菌类群包含33门367属,而珍珠猪毛菜根际土壤细菌类群涉及33门398属。根际土壤和非根际土壤中OUT数、多样性指数的显著差异进一步验证了两种土壤中细菌在群落组成上存在差异。且根际土壤细菌的多样性显著高于非根际土壤,与前人研究结果相一致[17-18]。本研究发现放线菌门在两种土壤中的丰度均最高,其中在非根际土壤中有更高的相对丰度(49.9%),可能是由于荒漠土壤为放线菌提供了木质素等营养物质[2]。放线菌门的许多属种具有耐旱、耐盐碱等特性,是干旱、盐碱等类型土壤中的优势菌群[19-20],本研究表明,放线菌门与土壤理化因子间无显著相关性。根际土壤中浮霉菌门的丰度仅次于放线菌门,其相对丰度从非根际土壤的9.1%增加到15.3%。酸杆菌门在根际土壤中的相对丰度从非根际土壤的4.7%增至6.2%,且与土壤pH呈显著负相关,表明其更适应于pH较低的土壤环境[2]。绿弯菌门和拟杆菌门在珍珠猪毛菜根际土壤中的相对丰度亦高于非根际土壤;且绿弯菌门与土壤全氮、全磷、速效氮和速效钾呈显著正相关,与杨秉珣等[21]对川西北沙化地区的研究结果一致。植物根际可改变土壤微环境,使得某些细菌类群的丰度或多样性增加或减少,形成有益于植物自身的根际细菌群落结构[22],如西瓜根际土壤中的优势类群是厚壁菌门、变形菌门和放线菌门[23];菊芋根际土壤中的优势类群是变形菌门、放线菌门、拟杆菌门和酸杆菌门[24],而造成不同植物根际土壤中优势菌群差异的机理尚需进一步研究。
在属水平上,荒漠土壤主要优势菌属为类诺卡氏菌属,其次为红色杆菌属。红色杆菌属属于放线菌门细菌,抗性较强,具有很好的固氮能力[25],与土壤全氮、速效氮和速效钾含量呈显著正相关,与土壤含水率和pH呈极显著负相关。珍珠猪毛菜根际土壤的最优势菌为红色杆菌属,其次为RB41,芽单胞菌属在根际土壤中相对丰度明显增加,可能与多磷酸盐积累有关[26],与土壤理化因子无显著关系。
珍珠猪毛菜根际和非根际土壤的细菌群落结构存在显著差异,土壤pH和含水率及全氮、全磷、速效氮、速效钾含量是影响土壤细菌群落组成的主要因子,但这些菌群对珍珠猪毛菜生长的影响尚不清楚,故需进一步研究菌群的生长代谢,掌握细菌群-珍珠猪毛菜的协同规律和环境生存机制,为生态恢复提供理论依据。
