青藏高原燕麦附着耐低温乳酸菌的筛选与鉴定
2022-07-08蔺豆豆琚泽亮柴继宽赵桂琴
蔺豆豆,琚泽亮,柴继宽,赵桂琴
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
青藏高原是我国主要牧区之一,气候严酷,暖季短暂、冷季漫长,枯草期长达7个月之久。冷季气温大幅度下降,使家畜在暖季放牧中增加的体重及营养被大量消耗,加之冷季饲草料供给不足,使老、弱、病畜难以安全越冬,形成“夏壮、秋肥、冬瘦、春死”的恶性循环,成为制约青藏高原畜牧业发展的主要因素[1]。人工种草是解决这一问题的有效途径[2]。燕麦(Avena sativa)对青藏高原高寒气候有独特的适应能力,具有易种植栽培、抗逆性强、产量高、品质优等优点,是该地区人工种草的首选草种,对缓解家畜季节性饲草紧缺、防止冬季掉膘、促进当地草地畜牧业的可持续发展具有重要作用。燕麦可晒制成青干草,也可生产青贮。但该地区草产品加工机械化程度低,技术手段落后,调制燕麦干草仍采用自然晾晒的方式,青藏高原地区秋季多雨,晒制青干草受天气影响很大,营养损耗较多,因此更适于青贮,在灌浆至乳熟期刈割,稍加晾晒含水量即可达到青贮要求[3]。调制青贮受天气影响小、生产的草产品适口性好、最大限度保留了饲草的营养,适宜在青藏高原及周边海拔较高、收获季节多雨的冷凉地区推广应用,也是长期保存饲草料的一种方式[4-6]。
一般情况下,低海拔地区饲草青贮发酵在40~45 d 即可完成,而青藏高原地区牧草多在8月底至9月收获,这时最高气温已降至 15~18 ℃左右,且昼夜温差非常大[7],进入 10月后气温更低。Liu 等[8]发现 10 ℃下柱花草(Stylosanthes guianensis)青贮料中乳酸含量较30 °C 降低了61.11%,乙酸含量下降了63.91%,好气性细菌数量增加了83.72%。玉米(Zea mays)在不同温度(5、10、15、20 和25 ℃)下的青贮,pH 下降速度随温度降低而显著减缓,发酵 30 d 后,5 ℃下青贮料的 pH 仍为 5.0[9]。垂穗披碱草(Elymus nutans)在 5 ℃下青贮 60 d 后 pH 值为 6.08,乳酸含量为0.34%,基本无青贮效果;在15 ℃下青贮60 d 后pH 值降至5.68,乳酸含量仅为1.44%,发酵程度较低[7]。在青藏高原高寒牧区,由于秋冬季气温很低,完成青贮发酵所需的时间明显增加,燕麦与箭筈豌豆(Vicia sativa)混播捆裹青贮要80 d 才能完成发酵,较长的发酵时间增加了营养物质的流失[10]。添加外源乳酸菌可促进青贮发酵,但一般乳酸菌适宜生长温度为20~37 ℃[10-11],市售乳酸菌添加剂的适宜温度远高于青藏高原的气温,因此在低温发酵中收效甚微[12]。要想尽快完成发酵,就需要添加低温下活性较高的耐低温乳酸菌。目前市场上未见此类乳酸菌制剂,但已有相关研究报道。崔棹茗等[13]发现,青藏高原乳酸菌具有耐低温、耐酸和耐盐性强等特性。保安安[14]从不同地区不同发酵阶段的垂穗披碱草青贮料中也分离得到6 株产酸和耐低温能力强的优异菌株。陈明霞等[15]通过限制性培养方法筛选得到3 株耐低温乳酸菌并成功应用于黑麦草(Lolium multiflorum)低温青贮。从不同温度和发酵阶段的垂穗披碱草青贮料中也分离得到耐低温的清酒乳杆菌、植物乳杆菌和戊糖片球菌[7]。由此可见,从植物生长地采集和分离适应环境的乳酸菌是可行的。
燕麦作为青藏高原地区主要的青贮原料,在青贮生产中面临秋冬气温低、发酵缓慢、营养损耗严重等问题[10,16],能否在其生产地分离得到耐低温的乳酸菌,用于促进低温发酵,目前尚未见相关研究报道。因此,本试验拟收集青藏高原不同海拔区燕麦植株附着乳酸菌资源,分析其耐低温和低pH 及产酸能力,筛选具有促进燕麦低温发酵的潜力菌株,以期为发掘和利用青藏高原本土特有乳酸菌资源、改善燕麦低温青贮发酵品质和促进优质燕麦青贮生产提供参考。
1 材料与方法
1.1 采样点地理信息
在甘肃省天祝抓喜秀龙乡、山丹军马二场、甘南州合作市那吾乡、甘南州玛曲县欧拉乡,青海省西宁市湟中县鲁沙尔镇、海北州海晏县西海镇、果洛玛沁县大武镇、玉树称多县清水河镇共8个地点进行采样。各样点的地理信息见表1。

表1 采样点地理信息Table 1 Geographical information of sampling sites
1.2 样品采集
于2020年8-9月在8个不同海拔区,采集处于开花至灌浆期的燕麦材料,装于提前灭菌的样袋,低温保存,带回实验室后在无菌环境下装入1 L 无菌聚乙烯瓶中密封,室温条件下进行青贮发酵,经3 和60 d 青贮发酵后用于乳酸菌的采集、分离、筛选和鉴定。
1.3 试验方法
1.3.1 乳酸菌的采集、分离与纯化 在无菌环境中分别称取青贮3 和60 d 后的燕麦青贮料10 g,放入锥形瓶中,加入90 mL 无菌生理盐水,置于摇床上振动(转速120 rpm·min-1)2 h,用无菌生理盐水进行梯度稀释,取3个适宜梯度涂板,每个梯度3个重复。在葡萄糖酵母膏蛋白胨培养基(glucose yeast extract peptone medium,GYP)(葡萄糖10 g,酵母提取物5 g,蛋白胨5 g,乙酸钠2 g,吐温805 mL,碳酸钙5 g,氯化钠5 g,盐溶液5 mL,琼脂15 g,溴甲酚紫0.04 g,蒸馏水1000 mL,pH=6.8)上对乳酸菌进行分离培养,37 ℃厌氧培养箱中培养48 h 后进行菌落挑选和分离。根据菌落大小、颜色、光泽、形状和是否具有透明环等特征采集单菌落。在MRS 培养基(蛋白胨10.0 g,牛肉粉5.0 g,葡萄糖20.0 g,酵母粉4.0 g,乙酸钠5.0 g,磷酸氢二钾2.0 g,硫酸镁0.2 g,柠檬酸三铵2.0 g,硫酸锰0.05 g,吐温801 mL,琼脂15 g,蒸馏水1 L,pH:6.2±0.2)上划线培养(37 ℃,1 d),重复划线分离培养3次,获得纯化的单菌株。对菌株进行革兰氏染色、镜检观察、过氧化氢酶试验,初步认定革兰氏染色阳性、过氧化氢酶阴性的菌株为乳酸菌[17]。利用MRS 液体培养基(不加琼脂)进行富集(37 ℃,1 d)后加入等体积甘油(40%),分装后-20 ℃保存备用。
将低温保存备用的菌株快速流水解冻,置于MRS 液体培养基中活化培养(37 ℃,1 d),进行2 次传代培养后,离心收集菌体,采用比浊法加入生理盐水调制菌液浓度为108 cfu·mL-1。
1.3.2 耐低温乳酸菌筛选 在MRS 液体培养基中接种活化菌株,接种量为3%,分别在20、15、10 和5 ℃恒温培养,其中5 ℃下培养10 d,10 和15 ℃下培养7 d,20 ℃下培养5 d,分析乳酸菌对温度的适应能力。配制含3.0%和6.0% NaCl 的MRS 液体培养基,接种活化菌株,接种量为3%,37 ℃培养3 d,分析乳酸菌的耐盐能力。用HCl溶液将 MRS 液体培养基 pH 分别调至 3.0、3.5、4.0、4.5 和 5.0,接种活化菌株,接种量为 3%,37 ℃培养 3 d,分析乳酸菌的耐酸性[18]。
将灭菌MRS 液体培养基分装,每试管5 mL,倒置放入杜氏小管,接种活化菌株,接种量为3%,37 ℃下培养3 d,观察是否产气。MRS 液体培养基中接种活化菌株,接种量为3%,37 ℃下培养30 h,每隔5 h 取样一次,测定培养液的pH 值,分析乳酸菌菌株的产酸速率。同时,以培养液为对照,测定样品在600 nm 波长下的吸光度值(optical density value,OD 值),分析乳酸菌的生长速度[19]。
1.3.3 耐低温乳酸菌的鉴定 利用16S rDNA 基因序列同源性分析方法进行菌种的初步鉴定。将活化的菌株在MRS 培养基中富集培养24 h 后离心收集菌体,参照生工Bacteria DNA Ki(t上海生工生物科技有限公司)试剂盒使用说明提取 DNA,进行 16S rDNA 扩增,引物序列为:27f(5′-AGAGTTTGATCCTGGCTCAG-3′),1492r(5′-TACCTTGTTACGACT-3′),由上海生工生物科技有限公司合成。PCR 扩增反应体系:2×PCR Master Mix,上下游引物各 1 μL,DNA 模板 1 μL,9.5 μL ddH2O,总体积为 25 μL。反应条件:94 ℃预变性 5 min;94 ℃变性 30 s,51 ℃退火 45 s、72 ℃延伸 1 min,30个循环;最后 72 ℃延伸 10 min,4 ℃保存。取 5 μL PCR 产物于1%琼脂糖凝胶上电泳以检测目的条带。将扩增产物送至上海生工生物科技有限公司测序。
将乳酸菌菌株 16S rDNA 基因序列用 BLAST(basic local alignment search tool,http://blast. ncbi. nlm. nih.gov/blast.cgi)在GenBank 中搜索,与待测菌株相似性最高的已知分类地位的菌种比对,初步确定待测菌株的属种。然后从GenBank 数据库中下载已知乳酸菌菌株的16S rDNA 基因序列,用Clustal X 进行序列比对后,用MEGA 5.0 的Neighbor-joining 法构建系统发育树,进行2000 次Bootstrap 检验。结合API 50 CHL 发酵试剂盒(BioMérieux,法国),按照试剂盒说明书测定不同菌株的碳源利用差异,鉴定菌种。
1.4 数据统计与分析
采用Excel 2019 对数据进行初步整理,采用SPSS 21.0 软件对不同菌株测定指标进行单因子ANOVA 模型分析,结合Duncan 法进行多重比较(P<0.05)。试验误差以平均值的标准误(standard error of mean,SEM)表示。
2 结果与分析
2.1 乳酸菌的分离纯化及耐低温菌株筛选
通过挑选GYP 培养基上有黄色溶菌环的菌株,初步获得232 株乳酸菌。对初获菌株进行革兰氏染色、镜检观察和过氧化氢酶试验,筛选出56 株乳酸菌。将其分别在 5 ℃培养10 d、10 和 15 ℃培养 7 d,20 ℃培养5 d,根据生长情况进行筛选,得到18 株耐低温乳酸菌菌株(表2)。
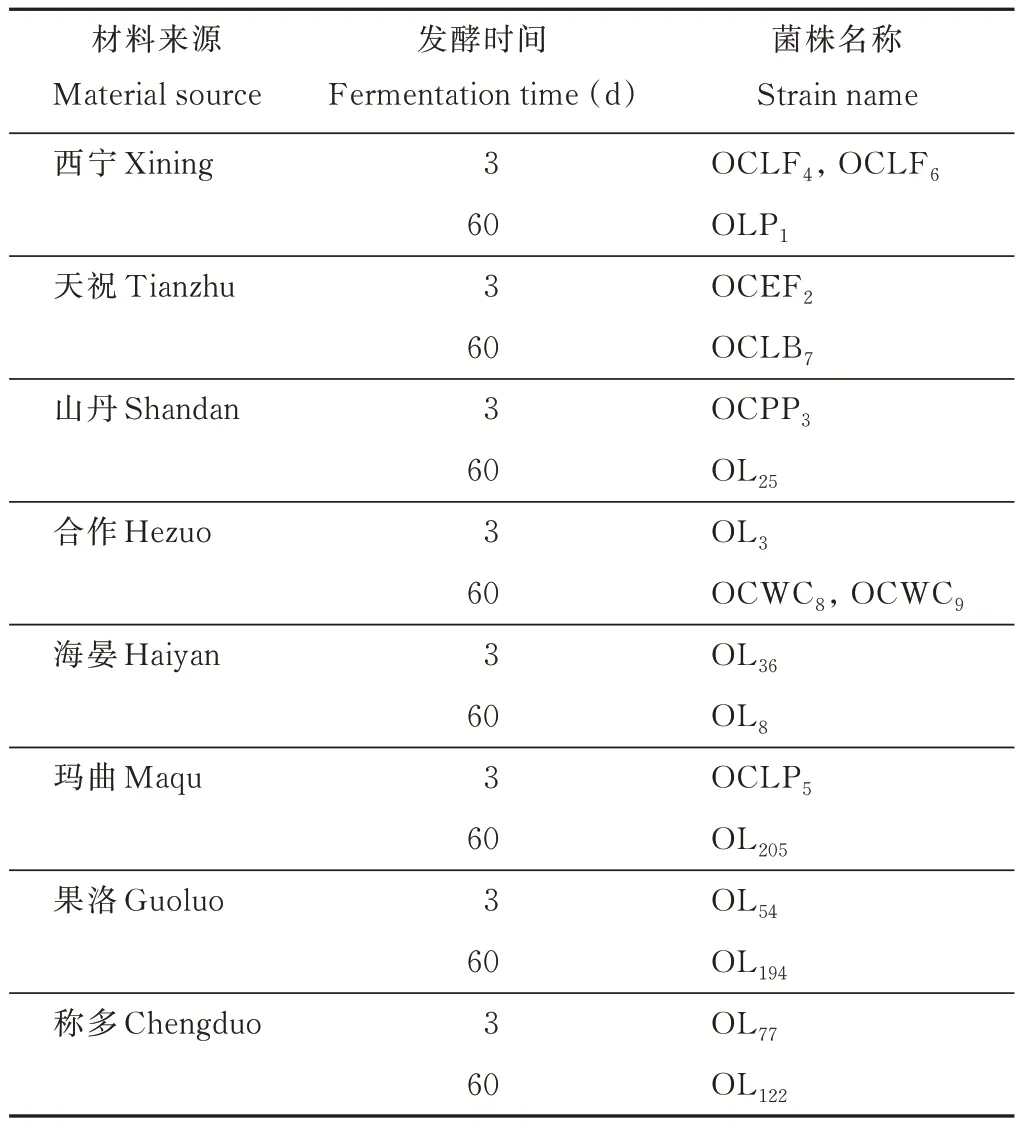
表2 燕麦青贮中分离的乳酸菌信息Table 2 Information of lactic acid bacteria isolated from oat silage
2.2 耐低温乳酸菌生理特性分析
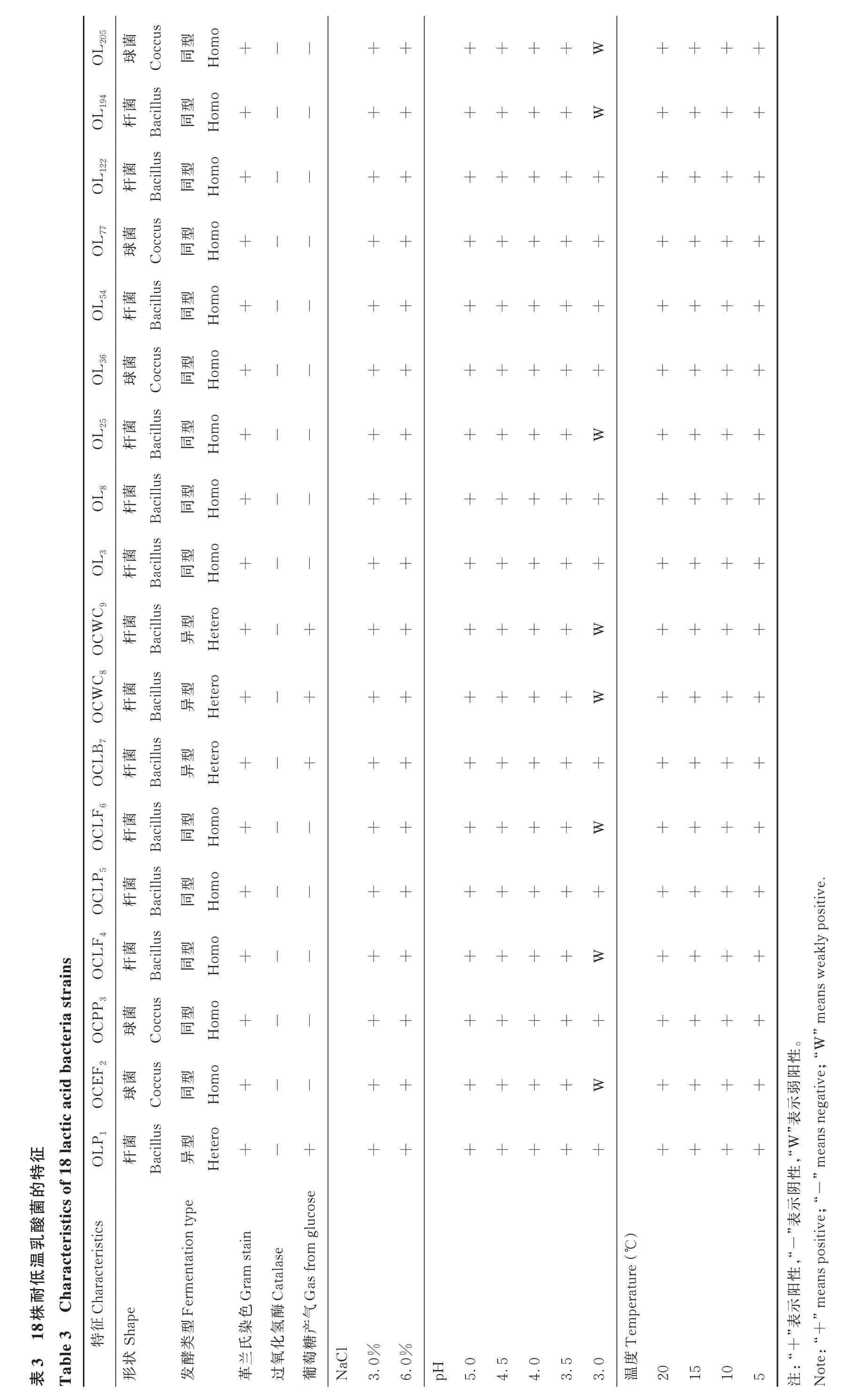
分离筛选的18 株耐低温乳酸菌生理特性如表3所示。所有乳酸菌菌株均为革兰氏阳性、过氧化氢酶促阴性细菌。其中,OLP1、OCLB7、OCWC8和 OCWC9为异型发酵乳酸菌,OCEF2、OCPP3、OCLF4、OCLP5、OCLF6、OL3、OL8、OL25、OL36、OL54、OL77、OL122、OL194和 OL205为 同 型 发 酵 乳 酸 菌 。 OCEF2、OCPP3、OL36、OL77和 OL205为球菌,OLP1、OCLB7、OCWC8、OCWC9、OCLF4、OCLP5、OCLF6、OL3、OL8、OL25、OL54、OL122和OL194为杆菌。
OL205菌 型mo球ccus Co 同Ho +--OL19 4菌 型mo杆Bacillus同Ho +--OL122菌 型mo杆Bacillus同Ho +--OL77菌 型mo球ccus Co 同Ho +--OL54菌 型mo杆Bacillus同Ho +--OL36菌 型mo球ccus Co 同Ho +--OL25菌 型mo杆Bacillus同Ho +--OL8菌 型mo杆Bacillus同Ho +--OL3菌 型mo杆Bacillus同Ho +--WC9菌 型OC 杆Bacillus异Hetero+-+WC8菌 型OC 杆Bacillus异Hetero+-+L B7 OC 菌 型杆Bacillus异Hetero+-+L F6 OC 菌 型mo杆Bacillus同Ho +--L P5 OC 菌 型mo杆Bacillus同Ho +--acteria strains LF4 OC 菌 型mo杆Bacillus同Ho +--P P3 OC 菌 型mo球ccus Co 同Ho +--cid b EF2 ccus菌 型mo OC 球Co 同Ho +--征8 lactic a Bacillus菌Hetero P1型+-+OL特f 1杆 异se菌e cs o的p o ++ ++++W ++++ ++ ++++W ++++ ++ +++++ ++++ ++ +++++ ++++ ++ +++++ ++++ ++ +++++ ++++ ++ ++++W ++++ ++ +++++ ++++ ++ +++++ ++++ ++ ++++W ++++ ++ ++++W ++++ ++ +++++ ++++ ++ ++++W ++++e. ++ +++++ ++++ositiv ++ ++++W ++++eakly p eans w ++ +++++ ++++。性阳e;“W”m ++ ++++W ++++弱示ativ eg ++ +++++ ++++eans n性阴cteristi 示酸,“-”e;“-”m aracteristics乳tation ty Catalase c lu m g 表ositiv s p Gas fro性低ra stain en Ch℃)阳温ble 3 C Ferm 8 株e 色表Gram耐ha酶erature(ap 气+”+”mean表征Sh 3 1型 染氢产化类氏糖Temp兰表Ta 状 酵 氧萄,“W”0%0%特pH5.050度形NaCl发过3.葡6.05革4.4.3.3.示温2015105:“te:“注No
18 株乳酸菌都能在3.0%和6.0% NaCl 条件下稳定生长,具有良好的耐盐性(表3),其在pH 为3.0~5.0 条件下也生长良好,尤其是 OLP1、OCPP3、OCLP5、OCLB7、OL3、OL8、OL36、OL54、OL77和 OL122在 pH 为 3.0 的条件下仍可良好生长,表明其对酸性环境的适应性更强。由于同型乳酸菌产酸效率更高,造成的干物质损失更小,因此,选取 14 株同型发酵乳酸菌(OCEF2、OCPP3、OCLF4、OCLP5、OCLF6、OL3、OL8、OL25、OL36、OL54、OL77、OL122、OL194和OL205)测定产酸速率和生长速度。
由表 4 可知,培养 5 h 后,菌株 OCPP3、OL3、OL8、OL25、OL36、OL54、OL77和 OL122的产酸速度较快,pH 均降至4.7 以下,显著低于其他菌株(P<0.05)。培养 10 h 后,菌株 OL77产酸最快,pH 降至 3.74,其次为 OL3和 OL25,pH降至 3.79。菌株 OCEF2、OCLF4、OCLP5、OCLF6、OL194和 OL205产酸引起的 pH 值降幅加大。培养 15 h 后,菌株OCPP3、OL3、OL8、OL25、OL36、OL54、OL77和 OL122的 pH 降至 3.8 以下,显著低于其他菌株(P<0.05)。培养 20 h后,所有菌株的pH 值变化均趋于平缓,在30 h 后所有菌株的pH 值均降至4.2 以下,最低的为菌株OL8,pH 为3.61,其次为 OL77和 OL54,pH 均为 3.62。
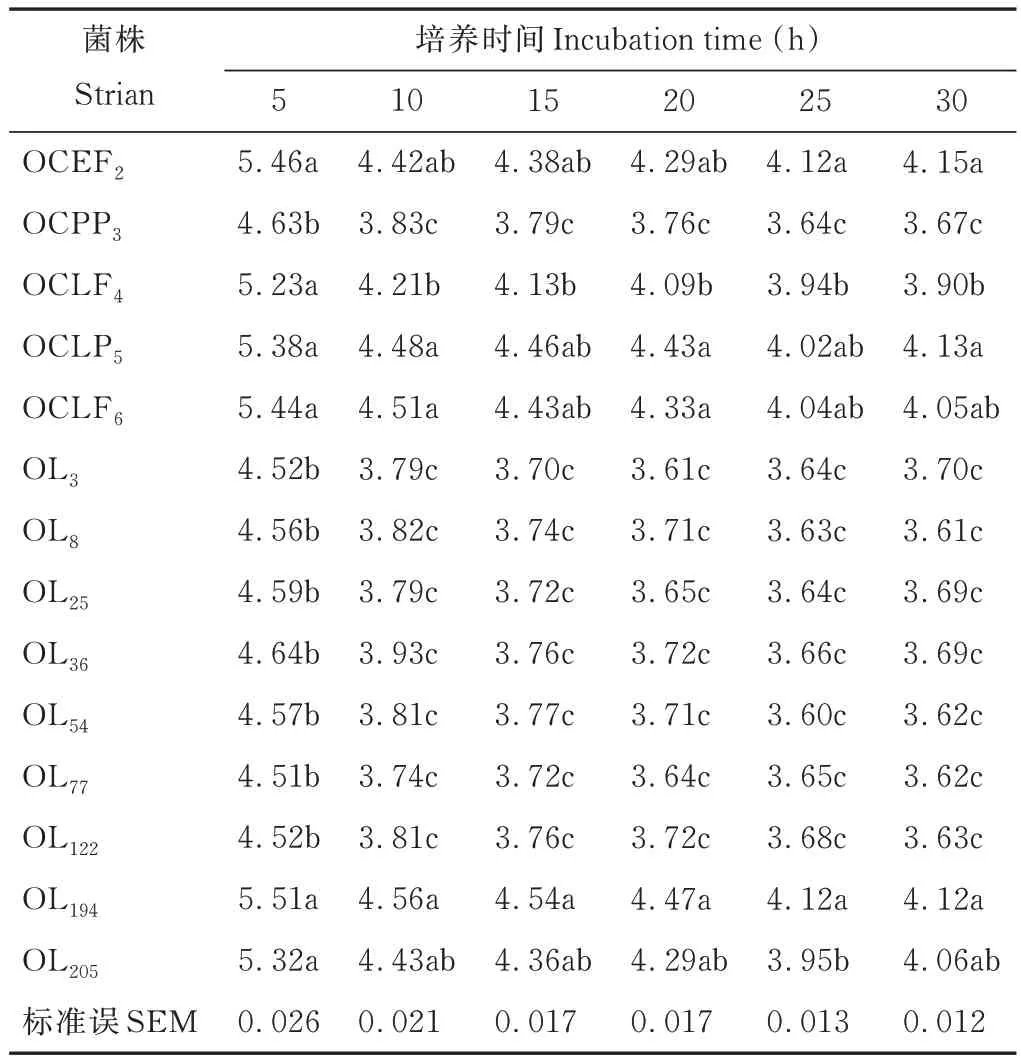
表4 供试乳酸菌的产酸速率Table 4 Acid production rate of tested lactic acid bacteria
供试菌株的生长速度呈现出和产酸速率类似的规律(表 5)。培养 0~5 h,菌株 OCPP3、OL3、OL8、OL25、OL36、OL54、OL77和OL122处于对数生长期,OD 值迅速上升,均大于1.60(P<0.05),其他菌株的生长在起始阶段较为迟滞。5~10 h,菌株 OCEF2、OCLF4、OCLP5、OCLF6、OL194和 OL205的生长速度增加,OD 值上升,平均为 1.68。10~25 h,所有菌株的生长速度开始放缓,仍以菌株 OCPP3、OL3、OL8、OL25、OL36、OL54、OL77和 OL122的繁殖量较高(P<0.05)。培养 25 h 后,菌株 OL77的 OD 值在所有培养菌株中最高,达2.52,其次为OL54,为2.50。培养30 h 后所有菌株均未出现明显衰退。
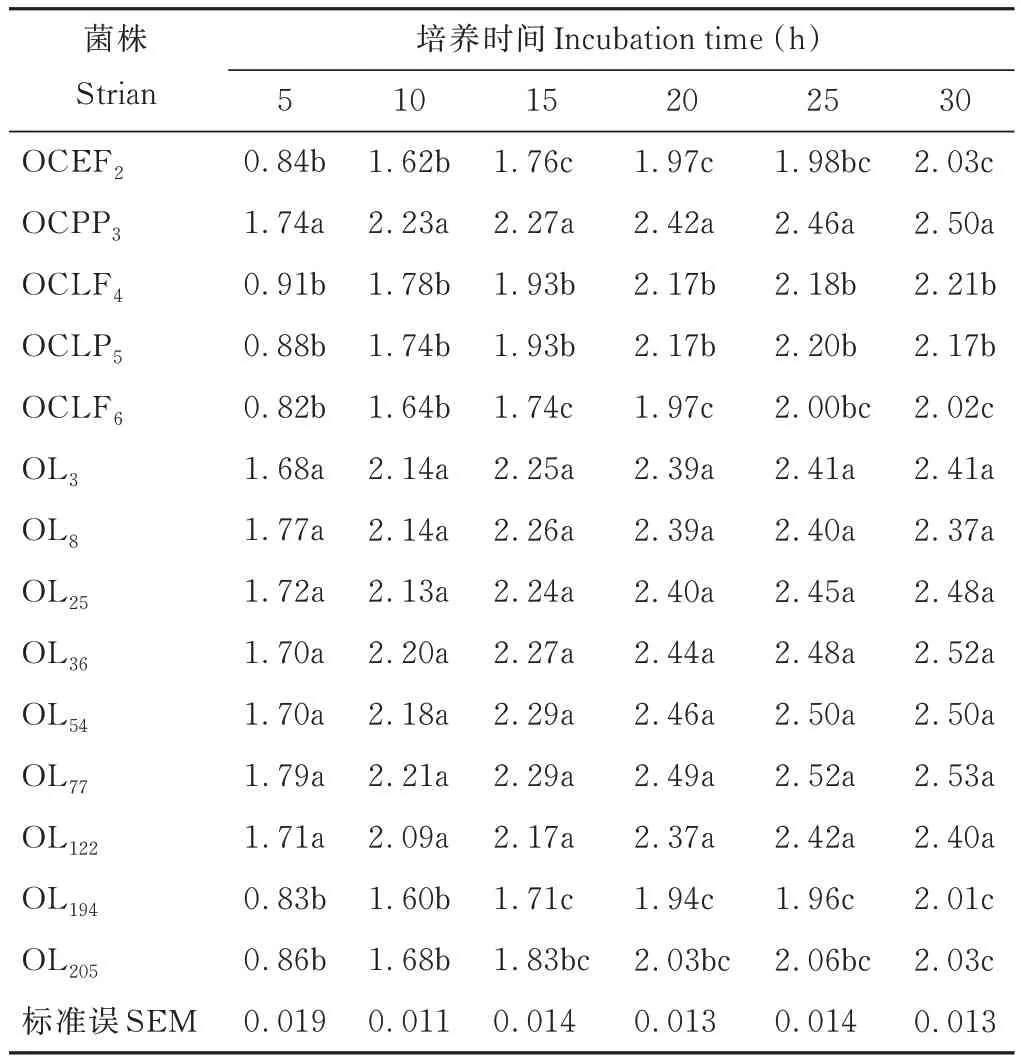
表5 供试乳酸菌的生长速率Table 5 Growth rate of tested lactic acid bacteria
2.3 耐低温乳酸菌的分子鉴定
结合表6 和图1 可知,菌株OLP1与类谷糠乳杆菌(Lactobacillus parafarraginis)处于同一族群,进化亲缘度100%,与对应标准菌株序列相似性为99.73%。菌株OCEF2与屎肠球菌(Enterococcus faecium)处于同一族群,进化亲缘度98%,序列相似度为97.94%。OCPP3、OL36、OL77和 OL205与戊糖片球菌(Pediococcus pentosaceus)处于同一族群,进化亲缘度98%,且与标准菌株的序列相似度达97.99%以上。菌株OCLF4和 OCLF6与香肠乳杆菌(Lactobacillus farciminis)处于同一族群,进化亲缘度为96%,与标准菌株的序列相似度分别为99.51%和98.72%。OCLP5与类食品乳杆菌(Lactobacillus paralimentarius)和消化乳杆菌(Lactobacillus alimentarius)处于同一族群,进化亲缘度为89%,与标准菌株的序列相似度为98.43%。OCLB7与布氏乳杆菌(Lactobacillus buchneri)处于同一族群,进化亲缘度为100%,序列相似度99.46%。OCWC8和OCWC9与魏氏乳杆菌聚在同一族群中,进化亲缘度为100%,与标准菌株16S rDNA 基因序列相似度达 99% 以上。菌株 OL3、OL8、OL25、OL54、OL122和OL194与植物乳杆菌(Lactobacillus plantarum)、戊糖乳杆菌(Lactobacillus pentosus)和 类植物 乳杆 菌(Lactobacillus paraplantarum)聚为一个族群,进化亲缘度为 100%,其中,菌 株 OL3、OL54、OL122和 OL194的16S rDNA 基因序列与植物乳杆菌标准菌株的相似度均大于戊糖乳杆菌,而菌株OL8和OL25与植物乳杆菌和戊糖乳杆菌标准菌株的相似度一致,但仅根据这一结果尚难鉴定到种水平。
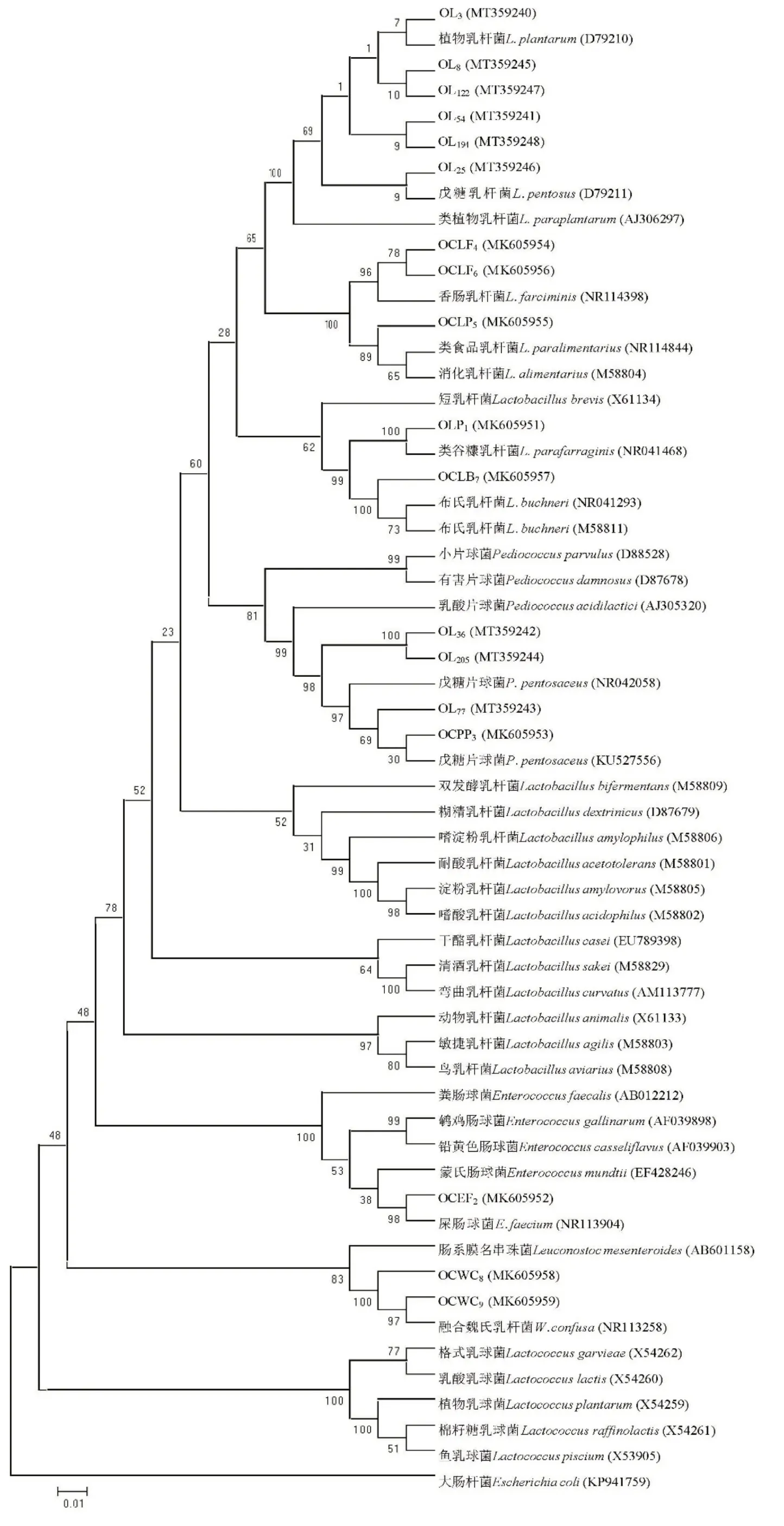
图1 筛选菌株及相关菌种的系统发育树Fig.1 Phylogenetic tree of screened lactic acid bacteria strains and related species
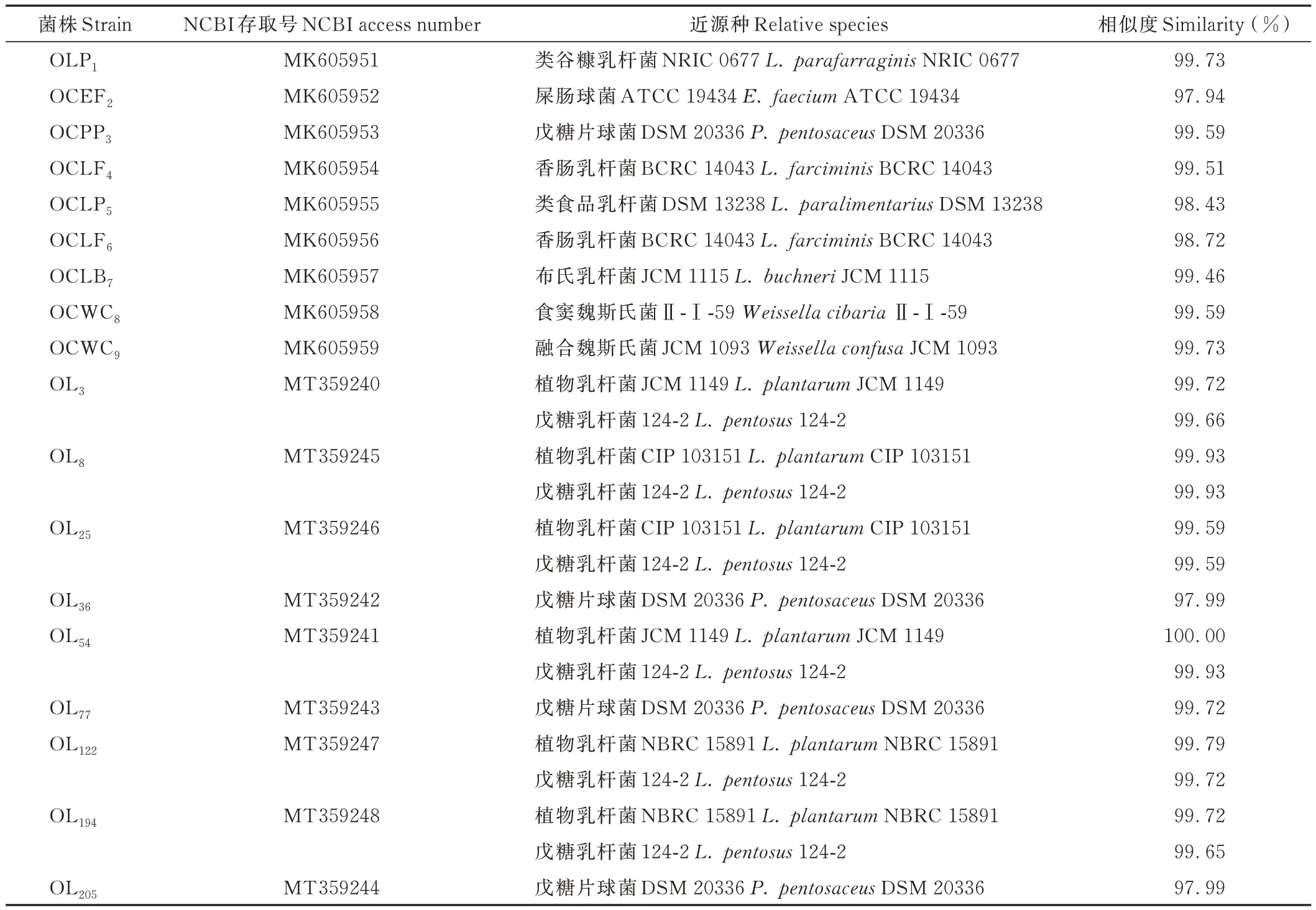
表6 乳酸菌菌株的16S rDNA 基因序列分析Table 6 Analysis of 16S rDNA gene sequences of lactic acid bacteria
筛选的18 株耐低温乳酸菌可初步分为9 种,反映了燕麦附生乳酸菌类群的丰富性。结合之前的产酸速率及生长速度试验,菌株 OCPP3、OL3、OL8、OL25、OL36、OL54、OL77和 OL122生长速度快、产酸速率高。因此,对这8个菌株进行糖发酵特性研究。
2.4 8个优选菌株的糖发酵特性分析
从表7 可知,8个菌株均能发酵的糖有L-阿拉伯糖、核糖、半乳糖、D-葡萄糖、D-果糖、D-甘露糖、甘露醇、山梨醇、N-乙酰-葡萄糖胺、苦杏仁苷、熊果苷、七叶灵、水杨苷、纤维二糖、麦芽糖、乳糖、蜜二糖、蔗糖、海藻糖、松三糖、β-龙胆二糖和葡萄糖酸盐,共22 种。均不能发酵的糖有赤藓糖醇、D-阿拉伯糖、L-木糖、阿东醇、L-山梨糖、半乳糖醇、肌糖、淀粉、肝糖、木糖醇、D-来苏糖、D-海藻糖、L-岩藻糖、L-阿拉伯糖醇、2-酮基-葡萄糖酸盐和5-酮基-葡萄糖酸盐,共16 种。

表7 8个优选菌株的糖发酵特性Table 7 Sugar fermentation profiles of 8 lactic acid bacteria strains
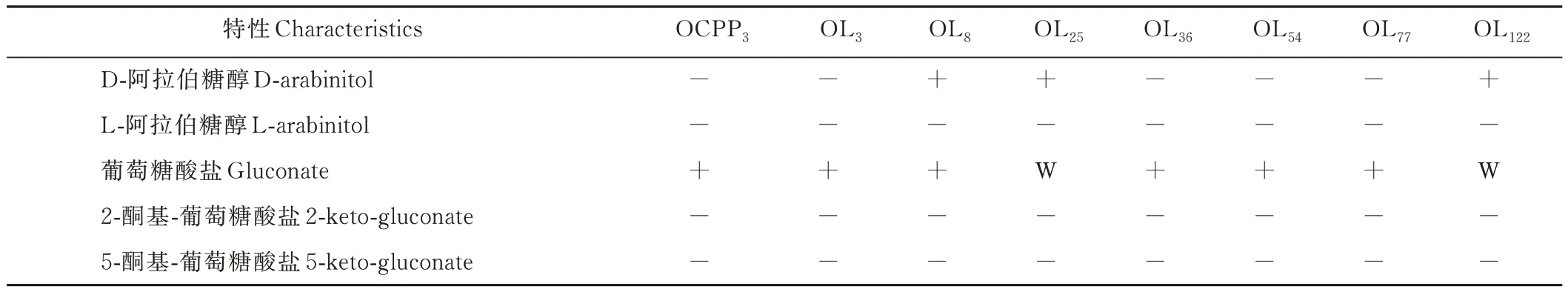
续表Continued Table
除此之外,菌株 OCPP3、OL36和 OL77均能发酵 D-木糖、D-棉籽糖、D-塔格糖,均不能发酵β-甲基-木糖苷、α-甲基-D-葡萄糖苷、D-松二糖和D-阿拉伯糖醇。菌株OL3和OL54均能发酵D-木糖、β-甲基-木糖苷、鼠李糖、胰岛素、D-松二糖和D-塔格糖,均不能发酵α-甲基-D-甘露糖苷、α-甲基-D-葡萄糖苷、D-棉籽糖和D-阿拉伯糖醇。菌株OL8、OL25和 OL122均能发酵 α-甲基-D-甘露糖苷、D-棉籽糖和 D-阿拉伯糖醇,均不能发酵 D-木糖、β-甲基-木糖苷、鼠李糖、胰岛素和D-塔格糖。
结合伯杰氏细菌鉴定手册和前人研究结果,戊糖乳杆菌可以发酵D-木糖和鼠李糖,植物乳杆菌不能发酵鼠李糖;植物乳杆菌能利用α-甲基-D-甘露糖苷、D-棉籽糖,而戊糖乳杆菌不能利用α-甲基-D-甘露糖苷和D-棉籽糖[18]。因此将菌株OL3和OL54鉴定为戊糖乳杆菌,菌株OL8、OL25和OL122鉴定为植物乳杆菌。
3 讨论
青贮过程受温度影响比较大,青贮时间较长时尤为明显[13]。低温会抑制微生物的活动和繁殖,延长青贮发酵时间。燕麦附着乳酸菌数量在高海拔地区均低于105 cfu·g-1,难以快速启动乳酸发酵[20]。随着发酵时间的延长,可溶性糖(water soluble carbohydrate,WSC)含量由于乳酸菌的消耗而显著降低,芽孢杆菌及酵母菌等对青贮原料中较难利用的物质如纤维等具有较强的降解能力,使青贮原料中可溶物质增加,营养流失加剧。适宜的添加剂可促进发酵进程并改善青贮发酵品质,在气温偏低、青贮发酵缓慢的高寒地区,添加剂的使用更具有重要意义。Zhang 等[12]研究发现,与对照菌株相比,在5 ℃下给小麦(Triticum aestivum)秸秆(干物质含量32%)接种筛选的耐低温植物乳杆菌,青贮发酵30 或60 d 后,可以显著改善青贮饲料的发酵品质。15 ℃条件下,添加3 株耐低温乳酸菌均可促进黑麦草的发酵,降低pH 值,减少蛋白质或氨基酸的分解,发酵品质显著提高[21]。因此,从高海拔地区燕麦附生乳酸菌种群中筛选生长速度快、产酸能力强的耐低温菌株,用于促进当地燕麦低温青贮发酵是可行的。
在青贮发酵中,约有20 多种乳酸菌起作用,可分成同型发酵乳酸菌和异型发酵乳酸菌两大类,其代谢过程差异较大,作用也不尽相同[22]。多数研究结果表明,同型发酵乳酸菌在产生乳酸和改善青贮饲料品质方面比异型发酵乳酸菌更有效[23];异型乳酸菌则更有利于提高有氧稳定性[22]。本研究中,通过对不同海拔地区燕麦青贮乳酸菌的分离筛选,获得18 株耐盐、耐酸和耐低温乳酸菌资源,其中OLP1、OCLB7、OCWC8和OCWC9为异型发酵乳杆菌,OCEF2、OCPP3和OCLF4等14 株为同型发酵乳杆菌,为燕麦低温青贮乳酸菌资源利用提供了基础材料。4 株异型发酵乳杆菌均来自发酵60 d 的燕麦青贮料,说明燕麦自然发酵后期异型乳酸菌可能成为主导菌株。这与Zhou 等[9]的研究结果类似,玉米在不同温度下自然青贮,在20 和25 ℃下,植物乳杆菌和戊糖片球菌在1 d 内占据优势,7 d 后异型发酵菌布氏乳杆菌开始出现,并最终占据主导地位;15 ℃可能是布氏乳杆菌生长的温度下限。尽管如此,在低于15 ℃的温度下发酵60 d 后,其他异型发酵乳酸菌仍在乳酸菌种群中占主导地位。本研究中,异型发酵乳杆菌包括类谷糠乳杆菌、布氏乳杆菌和魏氏乳杆菌,反映了燕麦自然青贮发酵后期异型发酵乳酸菌种类的丰富性。而菌株OCLB7(布氏乳杆菌)可以在低于15 ℃的环境中生长,可见青藏高原极端环境中分离出的乳酸菌对低温的适应能力更强。
不同细菌种类16S rDNA 基因在功能和进化上具有同源性,基因序列变异频率缓慢,因此在整体序列结构上极端保守,且拥有适宜的分子量大小,存储了丰富的生物信息,是系统进化分类的可靠选择[24]。对16S rDNA 序列进行分析,可以快速对微生物进行初步分类鉴定,基于16S rDNA 构建的系统进化树也可以确定菌种在进化中的位置。一般认为,在种分类水平上若2个分类单位间16S rDNA 序列同源性高于97.5%,则属于同一个种[25]。本研究中,菌株OLP1与类谷糠乳杆菌进化亲缘度为100%;OCEF2与屎肠球菌进化亲缘度为98%;OCPP3、OL36、OL77和OL205与戊糖片球菌进化亲缘度为98%;OCLB7与布氏乳杆菌进化亲缘度为100%;OCWC8和OCWC9与魏氏乳杆菌进化亲缘度为100%,基本可以确定属种。但OCLF4和OCLF6与香肠乳杆菌进化亲缘度为96%;OCLP5与类食品乳杆菌和消化乳杆菌进化亲缘度为89%,尚需要进一步鉴定。菌株OL3、OL8、OL25、OL54、OL122和OL194与植物乳杆菌、戊糖乳杆菌和类植物乳杆菌进化亲缘度为100%,但具体为哪一种尚不能确定。植物乳杆菌和戊糖乳杆菌在16S rDNA 序列上仅有2 bp 的差异,表现出极高的序列相似性,只通过16S rDNA 序列分析很难区分[13]。因此结合糖发酵结果,菌株OL3和OL54均能发酵D-木糖和鼠李糖,均不能发酵α-甲基-D-甘露糖苷和D-棉籽糖,因此鉴定为戊糖乳杆菌。而菌株OL8、OL25和OL122均能发酵α-甲基-D-甘露糖苷、D-棉籽糖,均不能发酵D-木糖和鼠李糖,因而将其鉴定为植物乳杆菌[21]。
乳酸菌作为生物型添加剂直接增加了青贮原料中乳酸菌的数量,提高了发酵初期乳酸菌与其他微生物的竞争能力,有利于促进乳酸发酵的进程。关于植物乳杆菌、戊糖片球菌和戊糖乳杆菌作为青贮添加剂的报道较多,效果也非常显著[12,26-31]。生长速度和产酸速率是筛选青贮用乳酸菌的重要指标[18]。本研究筛选的耐低温菌株中,戊糖片球菌 OCPP3、OL36和 OL77,戊糖乳杆菌 OL3和 OL54,植物乳杆菌 OL8、OL25和 OL122的生长速度更快,产酸速率也显著高于其他菌株;其中OL77、OL54和OL122表现较好,可作为燕麦低温青贮调制的备选添加菌株。
4 结论
1)青藏高原地区燕麦附着乳酸菌资源丰富、种类多样。从不同海拔地区初步分离到232 株乳酸菌资源,经进一步筛选,获得56 株菌株。
2)56 株菌株中,18个菌株体现出了良好的耐低温、耐低pH 和耐盐能力,其中4 株为异型发酵乳酸菌,14 株为同型发酵乳酸菌。
3)14 株同型发酵乳酸菌中,有8 株产酸速率和生长速度较快,其中3 株为戊糖片球菌,2 株为戊糖乳杆菌,3 株为植物乳杆菌。
4)综合考虑菌株的耐低温能力、产酸速率和生长速度及多样性,戊糖片球菌OL77、戊糖乳杆菌OL54和植物乳杆菌OL122可作为青藏高原地区燕麦低温青贮发酵的备选添加菌株。
