miR-363在脆性骨折患者血中的表达及对MC3T3-E1细胞成骨分化及增殖的影响
2022-06-29张爱新
张爱新, 张 卓, 张 杰
(北京市平谷区医院急诊科,北京市101200)
脆性骨折又称骨质疏松性骨折,是指轻微外伤所导致的非暴力性骨折[1]。尽管目前有许多方法可以改善脆性骨折的治疗,并加快骨折愈合过程,但治疗前景仍不能十分满意[2]。微小RNA(microRNA,miRNA)是一类小的非编码RNA,通过与信使RNA(messenger RNA,mRNA)的3′-非翻译区(3′-untranslated region,3′-UTR)特异性结合而调节目的基因表达[3-4]。研究发现,骨折愈合过程中一些miRNAs通过调节软骨细胞、成骨细胞和破骨细胞分化,在骨形成、吸收、重塑和修复中起重要的调节作用[5]。miR-363可通过减少多发性骨髓瘤患者骨溶解而抑制肿瘤进展[6],而其在成骨分化中的研究尚未见相关报道。本实验旨在研究miR-363在脆性骨折患者血中的表达及对小鼠胚胎成骨细胞前体细胞(mouse embryonic osteoblast precursor cells,MC3T3-E1)成骨分化及增殖的影响,并探讨其对Dickkopf相关蛋白1(Dickkopf-1,DKK1)与Wnt/β-catenin信号通路的调节作用。
1 资料和方法
1.1 仪器和试剂
普通PCR仪购自美国BIO-RAD公司;LightCycler480荧光定量PCR仪购自瑞士Roche公司。Trizol试剂、反转录与荧光定量PCR试剂均购自日本Takara公司;RIPA裂解液与Lipofectamine 2000转染试剂购自美国Life technology公司;双荧光素酶报告基因试剂购自美国Promega公司;模拟物及阴性对照购自苏州GenePharma公司;杜尔贝科改良伊格尔培养基(dulbecco's modified eagle medium,DMEM)、胎牛血清与Opti-MEM均购自美国Gibco公司;兔抗人DKK1与甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗体购自美国Abcam公司;噻唑蓝(MTT)与二甲亚砜购自美国Sigma-Aldrich公司;MC3T3-E1细胞购自中科院上海细胞库。
1.2 外周血标本收集
收集2015年1月—2018年12月本院急诊科就诊的34例脆性骨折患者外周血标本各5 mL,其中男20例,女14例,年龄32~74岁,平均(58.22±7.09)岁;收集11例本院体检的正常健康者外周血标本各5 mL,其中男6例,女5例,年龄40~76岁,平均(56.67±5.84)岁;同时收集上述患者术后1周、2周及4周外周血标本各5 mL。采集后的新鲜外周血标本立即放入抗凝管中,并置于-80 ℃冰箱中冷冻保存以备后续实验使用。纳入标准:临床表现为脆性骨折患者,影像学诊断明确。排除严重肝肾功能障碍、凝血功能障碍、关节炎、全身免疫系统疾病。本实验经过本院医学伦理委员会批准,并获得全部参与者的知情同意。
1.3 荧光定量PCR检测基因表达
按照Trizol试剂常规提取标本中总RNA,参照试剂盒说明书以RNA为模版进行反转录合成cDNA,以cDNA为模版进行荧光定量PCR反应。反应体系如下:模版cDNA 1 μL,上下游引物各0.5 μL,2×SYBR Green PCR Master Mix 10 μL,去离子水8 μL。反应条件为:95 ℃ 10 min,随后98 ℃ 10 s,59 ℃ 30 s,72 ℃ 30 s,共40个循环。U6与GAPDH为内参基因,人miR-363、DKK1、矮小相关转录因子2(runt-related transcription factor 2,RUNX2)、骨钙素(osteocalcin,OCN)、细胞周期蛋白D1(cyclin D1,CCND1)、连环蛋白β1(catenin beta 1,CTNNB1)和骨髓细胞瘤癌基因(myelocytomatosis oncogene,MYC)引物序列见表1。每组设计3个复孔,独立重复3次,并通过2-ΔΔCt法分析各基因的表达水平。

表1 引物序列
1.4 细胞培养及转染
MC3T3-E1细胞复苏后,采用DMEM培养基与10%胎牛血清在37 ℃、含5%CO2细胞培养箱中进行培养。取对数期生长的细胞进行细胞铺板,当细胞融合度达90%左右时,按照Lipofectamine 2000转染试剂说明书进行细胞转染,分别转染模拟物与阴性对照,6 h后采用荧光显微镜观察转染情况。
1.5 MC3T3-E1细胞诱导成骨分化实验
将转染后的细胞使用6孔板进行细胞铺板,每孔细胞数约为6×106个,当细胞融合度达到80%左右时,使用添加10 mmol/L β-甘油磷酸和50 mg/L抗坏血酸的DMEM培养基与10%胎牛血清开始诱导分化,分化培养基每3天更新一次,持续分化处理28天。
1.6 茜素红S染色实验检测MC3T3-E1细胞成骨分化
收集约1×106个诱导成骨分化28天MC3T3-E1细胞,用PBS缓冲液洗涤细胞3次,每次5 min。将细胞用冷却的0.05%戊二醛固定10 min,并用PBS缓冲液洗涤细胞3次,每次5 min。37 ℃下用40 mmol/L茜素红S染色5 min。将细胞用蒸馏水洗涤3次,每次5 min,然后用PBS缓冲液冲洗10 min。采用倒置显微镜下观察细胞显色情况。
1.7 MTT实验检测MC3T3-E1细胞增殖
将转染后的细胞使用96孔板进行细胞铺板,每孔细胞数约为4×103个,每孔每天加入16 μL MTT(5 g/L)并室温孵育4 h后去上清,连续3天,之后加入130 μL二甲亚砜孵育15 min溶解结晶。测定各孔450 nm波长时的光密度(optical density,OD),并绘制细胞增殖曲线。
1.8 双荧光素酶报告基因实验
通过microRNA.org生物信息学数据库预测miR-363靶基因。以psiCHECK2为基础载体,构建psiCHECK2-DKK1-wt野生型报告载体与psiCHECK2-DKK1-mut突变型报告载体。将转染后的细胞使用24孔板进行铺板,每孔细胞数约为8×104个,并转染野生型或突变型报告载体。24 h后按照双荧光素酶报告基因试剂说明书检测各孔海肾和萤火虫基因的荧光素酶强度。
1.9 Western blotting实验检测DKK1蛋白表达
收集约1×106个MC3T3-E1细胞,采用RIPA裂解液提取总蛋白,常规定量和变性。制备12%十二烷基硫酸钠聚丙烯酰胺凝胶,上样、电泳、转膜及封闭,加入抗DKK1与GAPDH抗体4 ℃孵育过夜,TBS-T缓冲液洗涤3次,每次5 min。37 ℃孵育相应二抗1 h,TBS-T缓冲液洗涤3次,每次5 min。暗室添加超敏化学发光液孵育20 s并曝光拍照。
1.10 统计学处理
2 结 果
2.1 miR-363在外周血中的表达情况
荧光定量PCR结果显示,miR-363在脆性骨折患者外周血中的表达水平低于正常健康者外周血(P<0.05);脆性骨折患者术后1周、2周及4周外周血中miR-363的表达水平均高于术前,且呈上调趋势,以4周的表达水平为最高(均P<0.05;表2)。

表2 miR-363 在正常健康者与脆性骨折患者外周血中的表达水平
2.2 MC3T3-E1细胞中过表达miR-363
模拟物组与阴性对照组MC3T3-E1细胞中可见大量红色荧光,而空白组中无红色荧光,提示细胞转染效率良好。荧光定量PCR实验分析可见模拟物组MC3T3-E1细胞中miR-363表达水平(142.93±14.66)高于阴性对照(1.09±0.09)与空白组(1.00±0.13)(均P<0.05),而阴性对照与空白组中miR-363表达水平差异无显著性(P>0.05;图1)。
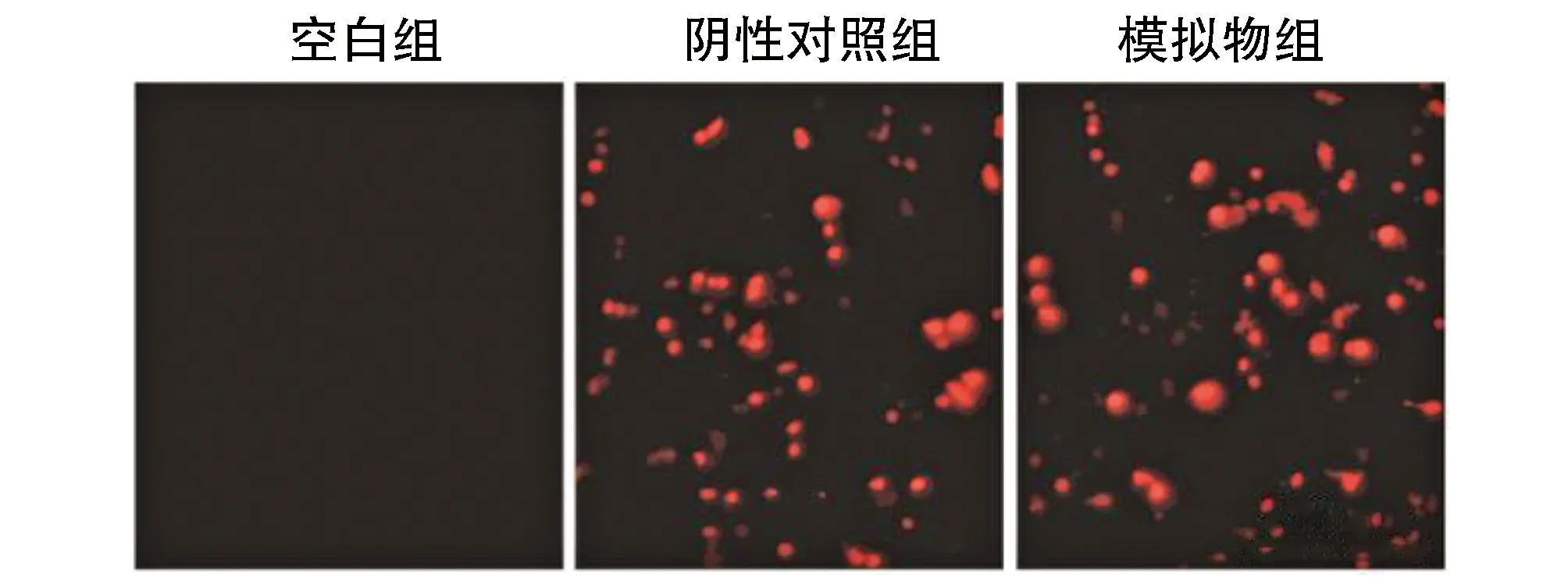
图1 MC3T3-E1细胞中过表达miR-363
2.3 过表达miR-363促进MC3T3-E1细胞成骨分化与增殖
茜素红S染色实验显示,模拟物组MC3T3-E1细胞矿化程度高于阴性对照组(P<0.05)。荧光定量PCR实验显示,模拟物组MC3T3-E1细胞中RUNX2及OCN mRNA表达水平均高于阴性对照组(均P<0.05)。MTT实验显示,在细胞培养后1天、2天、3天,模拟物组MC3T3-E1细胞增殖均高于阴性对照组(均P<0.05;图2)。

图2 过表达miR-363促进MC3T3-E1细胞成骨分化与增殖
2.4 过表达miR-363抑制靶基因DKK1表达
生物信息学显示,DKK1 mRNA 3′-UTR与miR-363存在互补结合位点。双荧光素酶报告基因实验显示,模拟物抑制野生型报告载体荧光素酶强度(P<0.05),而对突变型报告载体没有影响(P>0.05)。Western blotting实验显示,模拟物组MC3T3-E1细胞中DKK1蛋白表达水平低于阴性对照组(P<0.05;图3)。

图3 过表达miR-363下调靶基因DKK1表达
2.5 过表达miR-363促进Wnt/β-catenin信号通路相关基因表达
荧光定量PCR实验显示,模拟物组MC3T3-E1细胞中CCND1、CTNNB1及MYC mRNA表达水平均高于阴性对照组(均P<0.05;表3)。
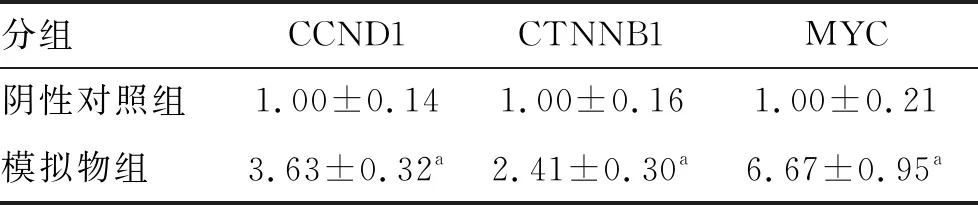
表3 阴性对照组与模拟物组CCND1、CTNNB1及MYC mRNA表达水平
3 讨 论
研究表明,miRNAs在成骨分化中发挥重要的调节作用[7]。Chen等[8]发现miR-7-5p调控趋化因子样受体1表达促进人间充质干细胞成骨分化。Liu等[9]发现miR-155通过下调骨形态发生蛋白9表达诱导小鼠胚胎成纤维细胞成骨分化。Yan等[10]报道miR-22通过抑制组蛋白脱乙酰基酶6表达促进人牙周膜干细胞的成骨分化。此外,Ren等[11]报道miR-23a-5p通过靶向有丝分裂原激活的蛋白激酶13表达抑制人骨髓间充质干细胞的成骨分化。miR-363位于人基因组Xq26.2区域,在胃癌、乳头状甲状腺癌、肝细胞癌和肺腺癌等多种肿瘤中起抑癌作用[12]。本文结果显示过表达miR-363促进MC3T3-E1细胞成骨分化及增殖,初步揭示了miR-363在骨折愈合中的作用。
DKK1是一种分泌型糖蛋白,通过特异性阻断成骨细胞中的Wnt/β-catenin信号通路传导,从而抑制成骨细胞的发育和活性[13]。研究发现,提高DKK1水平与活性可能导致成骨细胞活性受损和骨质流失[14]。此外,全基因组表达分析已将DKK1确定为一种与妇女绝经后骨矿物质密度变化相关的基因,与骨矿物质密度呈负相关[15]。本研究采用生物信息学分析miR-36的靶基因,发现DKK1是miR-363的靶基因,miR-363可与DKK1 mRNA 3′-UTR结合而抑制其蛋白表达。该结果初步表明,miR-363可能通过抑制靶基因DKK1表达促进MC3T3-E1细胞成骨分化及增殖。
Wnt/β-catenin信号通路是由配体无翅型MMTV整合位点家族蛋白和膜蛋白受体结合激发的一组多种下游通道的信号转导途径,在胚胎发育、组织器官形成以及肿瘤发生中起重要作用[16]。在胚胎骨骼形成过程中,抑制Wnt/β-catenin信号通路可阻止成骨细胞分化,从而诱导多能间充质细胞分化成软骨细胞[17]。此外,在动物和人类的骨折愈合中,Wnt/β-catenin信号通路的许多下游基因均被激活,包括CCND1、CTNNB1及MYC等[18]。本文结果显示过表达miR-363增加MC3T3-E1细胞中CCND1、CTNNB1及MYC的表达水平,初步表明miR-363可能通过激活Wnt/β-catenin信号通路促进MC3T3-E1细胞成骨分化及增殖。
综上所述,miR-363促进MC3T3-E1细胞成骨分化与增殖,该机制可能与调控靶基因DKK1与Wnt/β-catenin信号通路相关。miR-363有希望作为治疗骨折愈合的新靶点。
