非生长季降水对青藏高原高寒草甸优势种生物量稳定性的影响
2022-06-28刘琪源董世魁肖建南武胜男周秉荣
李 帅,刘琪源,董世魁,肖建南,武胜男,周秉荣,李 甫
1 山西农业大学资源环境学院,太谷 030801 2 北京师范大学环境学院,北京 100875 3 北京林业大学草业与草原学院,北京 100083 4 青海省气象科研所,西宁 810002
稳定性作为生态学和全球变化生物学中最具影响力的概念之一,是指生态系统在外界干扰(比如气候变化)下维持自身结构和功能的能力,且一直以来都是研究的热点[1—5]。稳定性是对种群和群落相互之间以及对环境变化的反应的综合衡量,尽管有许多方法可以测量稳定性,但一般采用生态变量(如生物量)的均值除以标准差来表征稳定性[1,6]。稳定性不仅可以反映外界干扰强度,而且对生态系统的功能和服务价值(如养分循环、生物量生产、畜牧产业的可持续性等)的维持具有很重要的作用[7—8]。
“质量比”假说认为,群落稳定性和生态系统功能在很大程度上取决于优势物种[9—14],群落间种间相互作用的强度也取决于优势物种[14—15]。有研究表明优势种的某些特征,比如数量可能会影响物种间的相互作用[16]。于此同时越来越多的研究表明,次优势物种可能也很重要[16—19]:比如有研究指出,相比于优势物种,次优势物种不仅直接响应气候变化,而且还可以间接对优势种产生影响[20—22]。
气候变化导致的气温升高和降水模式的变化将会对群落组成和生态系统功能产生重要影响[23—27]。根据IPCC的报告,全球平均升温的趋势为0.03—0.06℃/10a[23],并且预计会在未来进一步上升。处于高纬度区域的青藏高原气候变化会更加剧烈:有证据表明青藏高原的升温速率为0.37℃/10a显著高于全球速率[28];另外有研究表明在过去的五十年间,青藏高原的降水量增加了0.67mm/a[29],并且也将在未来持续增加[30],因此青藏高原整体呈现暖湿化的趋势[9,31]。相比于年际之间温度和降水的变化,生长季和非生长季气候变化模式的差异可能会对生态系统产生更重要的影响。有研究表明,非生长季的气候变暖会影响生态系统的稳定性[32—34]。在过去几十年里,非对称季节变暖在青藏高原上表现得十分明显[35—38]。不对称变暖可能与植物生产与消耗之间的平衡有关,也会导致植物生长过程中的水分有效性降低[39—42]。温度和降水的季节性不对称变化会引起物种组成[20,43]、营养循环和相互作用[44]的变化,最终影响生物量或稳定性[45—48]。
青藏高原是世界上最高的高原,平均海拔超过4000m,高寒草甸作为青藏高原最典型的生态系统之一,也对气候变化十分敏感[49]。但是关于高寒草甸对于气候变化的响应的研究结果并不一致,一方面这可能与单一的控制性实验,比如单独增温和单独增水的实验多于复合型的实验[50—51],另一方面,鉴于研究年限的差异,短期的控制性实验得到的结果相比于长时间序列观测实验得到的结果还存在差异,这可能会造成对于气候变化的响应被低估或高估[47,52—53]。关于高寒草甸植物群落对生长季/非生长季不对称增温的响应的相关研究时间年限较短(少于5a)[40]。因此,本研究利用青藏高原东部的高寒草甸17年(即2001年至2017年)的观测数据,研究优势物种本身的特征(包括高度和多度)以及次优势物种生物量稳定性,结合生长季/非生长季不对称增温和降水,探究对于优势种的生物量稳定性的影响。
1 材料与方法
1.1 研究区概况
研究区位于青海省海北藏族自治州海晏县西海镇(36.93°N,100.95°E,海拔3100 m)。草地类型为高寒草甸,优势物种为紫花针茅和洽草,植被盖度大于80%,土壤类型为砂壤土,年均降水量 400 cm,年蒸发量为 1538 mm,年平均气温-0.45 ℃。
1.2 实验设计
从 2001 年至 2017 年,在研究区将地势平坦的面积 100 m×100 m的区域分为 50 m×50 m的 4块,每一块进一步被分为25 m×25 m的 4 个小区,生长季围封,非生长季放牧,地表无明显径流。每年9月底在4块中每一块随机选择一个小区,小区按顺时针方向轮换每四年一轮。从中选取1m×1m的对优势物种(紫花针茅)及次优势物种群(洽草)的地上生物量进行监测。同时记录紫花针茅的多度、高度,收集到的植物样品在烘箱中高温(65℃)烘干至恒重。在离最近的海晏县气象站取得日平均气温以及日降水量。
1.3 统计分析
计算2001年至2017年的年平均气温、生长季平均气温(4—10月)、非生长季平均气温(10月—第二年的4月)、年降水量、生长季降水量(4—10月)、非生长季降水量(10月—第二年的4月)。
计算了紫花针茅和洽草各年处理间的平均生物量(μ)与其处理间的标准差(σ)的比值为生物量稳定性[1,4—6]。分别计算了年平均气温、生长季平均气温、非生长季平均气温、年降水量、生长季降水量、非生长季降水量、紫花针茅的高度、多度、紫花针茅生物量稳定性、洽草生物量稳定性随时间的变化趋势;采用线性回归包括研究了环境因子包括,年平均气温、生长季平均气温、非生长季平均气温、年降水量、生长季降水量、非生长季降水量对生物量稳定性的影响,以及生物因子,包括紫花针茅的高度、多度、洽草生物量稳定性与紫花针茅生物量稳定性的关系。
鉴于2010年紫花针茅生物量稳定性和2007年洽草生物量稳定性的值异常高(与平均值的偏差超过三倍标准差),本研究分别进行了剔除。依据线型回归找到对稳定性影响较大的因子,考虑到年均温和生长季平均温度,以及年降水和生长季降水量呈现显著的正相关且相关性系数大于0.7,为了避免共线性,本研究只将生长季平均温度和生长季降水纳入到模型的分析之中,并据此用建立理论模型。为了识别环境和生态因素影响植物群落的生物量稳定,本文结合Yang[29]和Ma[5]等已有研究结果构建了的先验模型——结构方程模型,包括年平均气温、生长季平均气温、非生长季平均气温、年降水量、生长季降水量、非生长季降水量、紫花针茅生物量稳定性、洽草生物量稳定性、紫花针茅高度、紫花针茅多度。利用R语言‘basicTrendline’包来进行简单线性回归,利用Amos 21 (IBM, Chicago)进行结构方程模型分析,非显著χ2检验(P>0.05)、拟合优度指数(GFI)(>0.90)和近似均方根误差(RMSEA)(<0.05)[40]。
2 结果
2.1 气候因子、生物因子的变化趋势
2001年到2017年,年平均气温以k=0.041的趋势显著升高(P<0.05),生长季平均气温升高趋势不显著(P=0.068),非生长季平均气温升高却不显著(P>0.05)。年降水量和生长季降水量均增加了100 mm,非生长季降水量没有明显的趋势(图1)。非生长季降水量的变化趋势与整个温度降水上升趋势不一致,没有明显的趋势;年降水量集中在生长季,生长季降水较多,非生长季降水较少(图1)。
不同于气温与降水变化趋势,两种植物的生物量稳定性均呈下降趋势,其中洽草呈显著下降趋势(P<0.05),紫花针茅下降趋势不显著(P>0.05);紫花针茅的高度和多度升高缓慢但不显著(P>0.05)(图2)。

图1 年平均气温、生长季平均气温、非生长季平均气温、年降水量、生长季降水量、非生长季降水量的变化趋势Fig.1 The change trend of annual mean temperature, growth season average temperature, non-growth season average temperature, annual precipitation, growth season precipitation, non-growth season precipitation灰色区域是置信水平为95%的置信区间

图2 紫花针茅生物量稳定性、洽草生物量稳定性、紫花针茅高度、紫花针茅多度的变化趋势Fig.2 The change trend of biomass stability of Stipa purpurea, and Koeleria cristata, the change trend of height and abundance of Stipa purpurea灰色区域是置信水平为95%的置信区间
2.2 环境因子、生物因子与紫花针茅生物量稳定性的关系
线性回归分析显示,与紫花针茅生物量稳定性显著相关的有3个因子,即非生长季年降水、紫花针茅的高度以及洽草的生物量稳定性呈极显著相关(P<0.05)(图3)。非生长季降水量增加会促进紫花针茅的生物量稳定性(图3)。
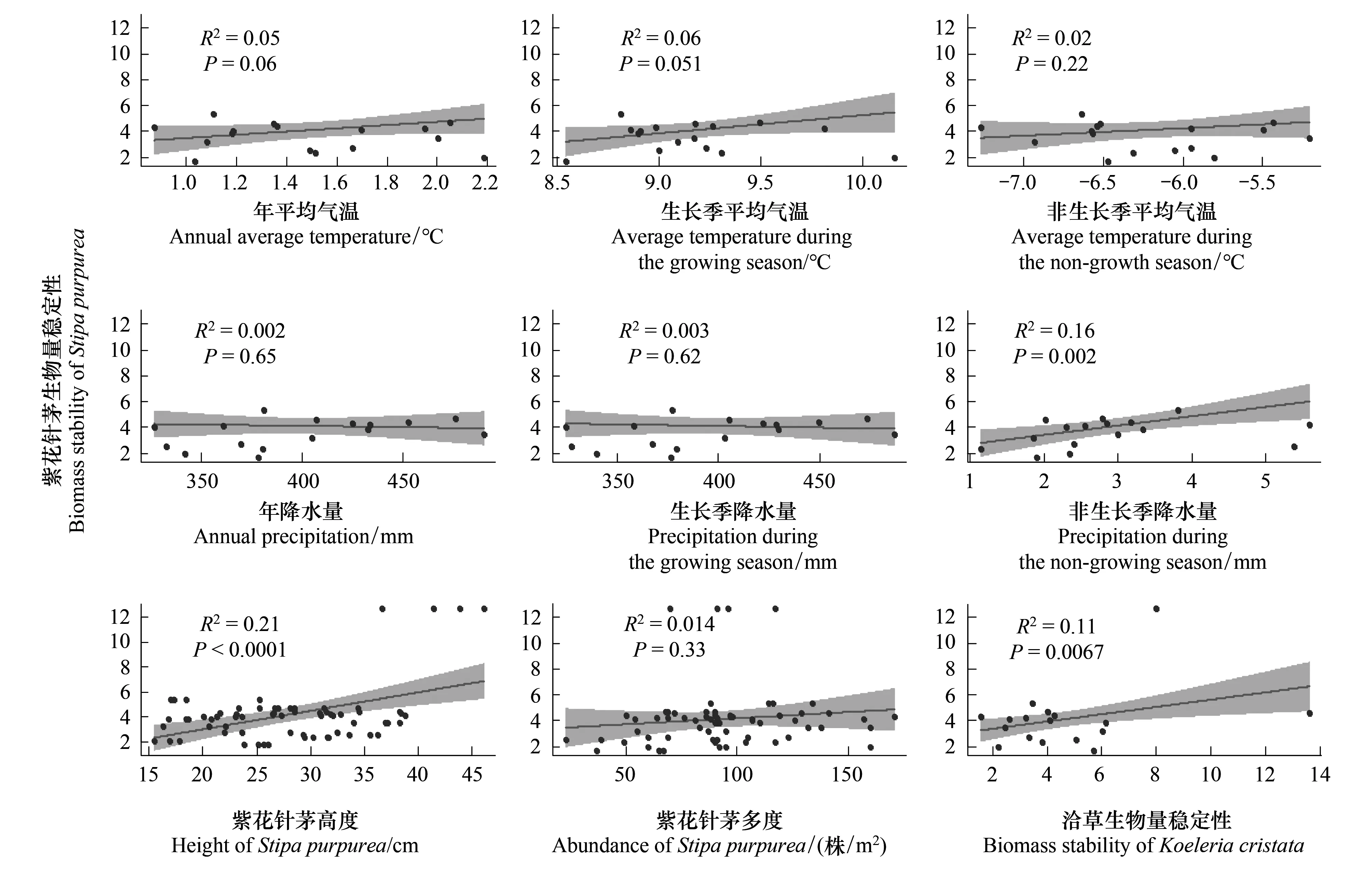
图3 环境因子、生物因子与优势种生物量稳定性的关系Fig.3 Relationship between environmental factors, biological factors and biomass stability of dominant species灰色区域是置信水平为95%的置信区间
2.3 环境因子、生物因子对紫花针茅生物量稳定性的影响
通过路径分析(根据Chi-square=13.00,df=7,P=0.08),紫花针茅的生物量稳定被非生长季降水、紫花针茅的高度和多度以及洽草的生物量稳定性解释了45%,其中高度、非生长季降水和洽草的生物量稳定性的贡献最大。紫花针茅的高度和多度,以及洽草的生物量稳定性都与生长季/非生长季温度和降水的变化,没有显著的关系(图4)。
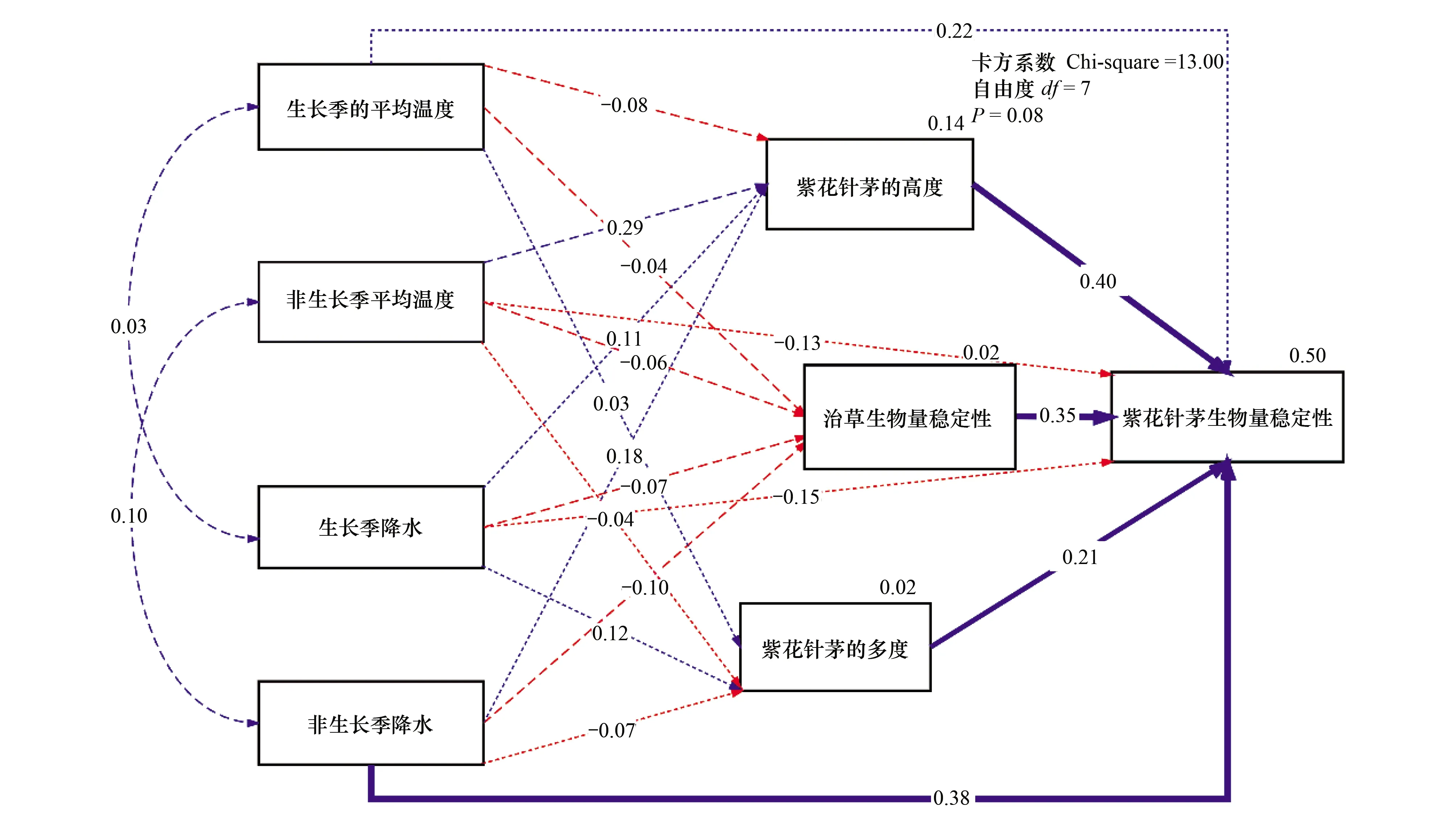
图4 紫花针茅生物量稳定性的环境因子和生物因子的结构方程模型Fig.4 Structural equation model of environmental factors and biological factors for biomass stability of Stipa purpurea响应量(紫花针茅生物量稳定性、洽草生物量稳定性、紫花针茅高度、紫花针茅多度)右上角的百分比表示模型(R2)能解释的方差;虚线表示不显著的影响;单向箭头上的数字为解释度系数,双向箭头上的数字为相关系数。每个结构内变量与响应量连线上的百分比是被解释的变量,红色代表负效应,蓝色代表正效应
3 讨论
随着全球气候变化的加剧,青藏高原的气候变化更加明显,高山草甸生态系统也受到了较为严重的影响[5,51,54]。本研究通过长期观测资料,分析生长季平均气温、非生长季平均气温、生长季降水量、非生长季降水量的数据,旨在厘清生长季/非生长季的气温和降水的不同步变化对于生态系统稳定性的影响。
近二十年的观测数据与其它研究结果一致,表明受全球气候变化影响,青藏高原上整体气候呈现暖湿化趋势[55],本研究发现,生长季的温度和降水与全年的温度降水均呈现显著的正相关,这也与青藏高原往往雨热同季的气候特征相吻合,即在生长季呈现暖湿化的趋势。但是,本研究并没有发现非生长季温度和降水呈现显著的趋势,这表明青藏高原过去十几年的季节不对称气候变化呈现增加的趋势,这与相关研究的结论一致[35—38]。
有研究表明,气候变化对植物的影响表现为:在高纬度地区,低温对植物生长的限制作用大于低纬度和高海拔地区[55],因此高寒植物的生物量稳定性随着气候变化的变化(特别是温度的增加)可能会发生比较大的变化[56]。但本研究发现,优势物种的生物量稳定性及其生态特征(高度和多度)在过去十几年期间,并没有发生显著的变化,与优势物种不同,本研究发现次优势物种的生物量稳定性在过去十几年期间呈现显著下降的趋势,这可能与优势物种之间的相互作用有关系。
有研究表明,优势物种在温度升高时优势度会升高[57—59],从而增加同时期优势种与次优势物种竞争作用的强度[60]。探究优势物种和次优势物种之间相互作用的方向和强度对气候变化的响应是理解气候变化对植物群落影响的基础。优势物种决定了群落中种间相互作用的强度并能对气候变化做出及时的响应[24],优势种群对生态系统结构和功能具有潜在的影响[41—42]。但是本研究发现,在环境因子里面只有非生长季降水与紫花针茅生物量稳定性呈现显著的正相关,此外紫花针茅自身的生态特征,如高度也与其生物量稳定呈现显著的正相关,以上表明紫花针茅作为优势种对气候变化更敏感且受到的影响是正面的[34],此外优势物种的生物量稳定性也会受到自身特征以及次优势物种的影响。
在高寒地区,温度是平衡竞争和促进关系的决定因素[61]。生长季平均气温的升高直接缓解高寒地区低温胁迫[62]。非生长季平均气温升高减少了霜冻灾害的发生频率,提高了植物的抗旱性[63],同时还提高了幼苗的存活率。其次,部分永冻层和积雪随着冬季温度升温而融化,增加了土壤在冬季的含水量,保护了植物地下掩埋部分,有利于植物在其后春夏两季的生长发育[63—64]。但是,本研究并没有发现生长季或者非生长季温度并没有对优势物种或者次优势物种稳定性产生影响,这与Fu等人[41]的研究结果并不一致,这可能与时间的年限有关系,即短期的控制性实验与长期气候变化记录是存在一定的差异的,本研究猜测优势物种与次优势物种的相互作用缓冲了季节不对称增温导致的效应,但是相关的结果还需要进一步的实验验证。此外,本研究并没有考虑次优势物种的高度和多度对于次优势物种和优势物种生物量稳定性的影响,这主要是其数据的缺失导致无法满足统计学要求,这会造成一定的误差,但是本文的研究结果可以反映生长季和非生长季不同步气候变化背景下,优势物种本身的特征(包括高度和多度)以及次优势物种生物量稳定性对于优势种的生物量稳定性的影响。
降水是高寒群落生产的主要驱动力,影响群落生物量稳定性[62,65],尤其高寒地区,降水往往成为植物生长的限制因子[66]。降水量的季节变化会影响生物量的稳定性[67],分析得出非生长季降水量与紫花针茅生物量稳定性的正相关性,大于生长季平均气温、非生长季平均气温,甚至大于生长季降水量。然而紫花针茅多度对降水增加的响应与年降水量无关,这与Wu等人的研究一致[68],只与生长季/非生长季降水量有关。本研究猜测,降水量的增加会在某种程度上解除温度升高导致的水分限制,特别是在植物开始返青的时候,最终生物量的稳定性就会随着暖湿化的气候变化而增加[69,70—71]。于此同时,本研究也从优势物种和次优势物种稳定性与气候变化的关系中验证了本研究的推论,即优势物种决定了群落中种间相互作用的强度并能对气候变化做出及时的响应[38,61]。
4 结论
青藏高原上高寒草甸对于气候变化较为敏感,但形成了对持续的暖湿化进行适应的策略,以保持自身生物量稳定性;优势种和次优势种对生长季/非生长季不对称升温的响应并不显著;前人的研究很少关注非生长季降水量对于生态系统稳定性的影响,本研究发现非生长季的降水会促进次优势物种的生长,最终促进高寒草甸中优势物种的生物量稳定性,预计在青藏高原未来的暖湿化的气候变化下,非生长季的降水结合物种间尤其是优势物种之间相互作用,并最终决定高寒草甸的稳定性。
